
Abstract
A number of single nucleotide polymorphisms (SNPs) within the mprF open reading frame (ORF) have been associated with daptomycin-resistance (DAP-R) in Staphylococcus aureus. Such SNPs have been found throughout the mprF ORF, although there are clearly preferred “hot spots” within this gene frequently linked to DAP-R phenotype. These mprF SNPs are often correlated with a gain-in-function phenotype, either in terms of increased production (synthase activity) and/or enhanced translocation (translocase activity) of lysyl-phosphatidylglycerol (L-PG) within its cell membrane. However, it is unclear if multiple hot spot mprF SNPs can accumulate within mprF ORFs and cause additive elevations of DAP minimum inhibitory concentrations (MICs). In this study, we used a previously well-characterized plasmid complementation system in S. aureus Newman
Introduction
O
In the current study, we utilized an isogenic mprF deletion (
Materials and Methods
Bacterial strains and culture conditions
The bacterial strains and plasmids used in this study are shown in Table 1. We used the wild-type S. aureus Newman strain,
17
its isogenic mprF knockout mutant strain (
Ampr, ampicillin resistance; CL, cardiolipin; Cmr, chloramphenicol resistance; Emr, erythromycin resistance.
All S. aureus strains were cultured in either Tryptic Soy Broth (TSB; Difco Laboratories, Detroit, MI) or Mueller-Hinton Broth (MHB; Difco Laboratories) depending on each assay. Liquid cultures were grown in Erlenmeyer flasks at 37°C with shaking (250 rpm) for aeration in a volume that was less than 10% of the flask volume.
DNA sample preparations and mprF complementation
Genomic DNA samples were isolated from S. aureus cells as described by Dyer and Iandolo. 21 Extraction and purification of plasmid DNA samples were performed using Wizard Plus kits (Promega, Inc., Madison, WI.). Preparation of competent Escherichia coli DH5α and transformation of plasmid constructs were accomplished as described previously. 22
Electroporation of plasmids into electrocompetent S. aureus cells was performed as described by Schenk and Laddaga. 23 Briefly, overnight S. aureus cultures grown in B2 broth (1.0% casein hydrolysate, 2.5% yeast extract, 0.1% K2HPO4 [pH 7.5], 0.5% glucose, 2.5% NaCl) at 37°C with shaking (250 rpm) were diluted into 25 ml of fresh B2 broth at OD600 of 0.25. The cells were grown until they reached mid-log phase of growth (∼OD600 of 0.5), pelleted by centrifugation, washed thrice with deionized water, and then resuspended in 5 ml of 10% glycerol solution. The cells were then incubated for 15 min at RT, pelleted again, and resuspended in 0.5 ml of 10% glycerol solution and used for electroporation. The cells and plasmid DNA were electroporated at 100 ohms resistance, 25 μF capacitance, and 2.3 kV in a Gene Pulser™ apparatus (Bio-Rad Laboratories).
Complementation with double point–mutated forms of mprF genes in Newman
Antibiotics and MIC determination
Ampicillin and chloramphenicol were used at 100 and 5 μg/ml, respectively, during the cloning experiments. DAP was kindly provided by Cubist Pharmaceuticals (Lexington, MA). The MICs to DAP and oxacillin (OX) were determined by standard E-test. Three independent experimental runs were performed to determine the DAP and OX MICs.
Population analysis
To determine DAP-R profiles of the study strains, DAP population analyses were performed as described before.16,24 Briefly, initial inoculum of ∼2 × 108 CFU/ml S. aureus cells was exposed to 0–8 μg/ml of DAP concentrations representing sublethal-to-lethal levels. All DAP population analyses were done in the presence of 50 μg/ml calcium (Ca2+). The area-under-the curve was calculated as before. 10 Three independent experimental runs were performed.
hNP-1 and LL-37 susceptibility assays
Since standard MIC testing in MHB or TSB may inhibit CAMP activities, in vitro survival assays were performed with hNP-1 and LL-37 using a 2-hr microdilution method in Eagle's minimal Essential medium.19,25 The two prototypical antimicrobial peptides, hNP-1 (an α-defensin in neutrophils) and LL-37 (a human cathelicidin found in neutrophils and skin), were purchased from Peptide International (Louisville, KY). A final inoculum of 5 × 103 CFUs of overnight grown S. aureus cells was exposed to hNP-1 (10 μg/ml) and LL-37 (1 μg/ml). 9 These concentrations of hNP-1 and LL-37 represented a sublethal level against the wild-type Newman S. aureus strain, as determined in pilot experiments. Three independent runs were performed on separate days.
CM PL contents
To quantify in our study strains the relative proportions of L-PG among the three major staphylococcal PLs (i.e., L-PG, PG, and cardiolipin [CL]),5,6,26 CM PLs were extracted, separated, and identified using two-dimensional thin-layer chromatography (2D-TLC) as described previously.19,27 Individual TLC PL spots were scraped from the TLC plates and then quantified by a well-defined spectrophotometric analysis.19,27 Data are expressed as the proportionalities (±standard deviation [SD]) of the L-PG, PG, and CL for each staphylococcal strain. Four independent experiments were performed on separate days to analyze the PL profiles.
Determination of surface positive charge
To assess relative net charges on the study strains, cytochrome c (Sigma, St. Louis, MO) binding to cell surface was measured by quantifying the amount of the unbound cytochrome c in reaction mixtures.10,19,27,28 For the assay, S. aureus cells were grown overnight for ∼17 hr in TSB, washed thrice with 20 mM morpholinepropanesulfonic acid (MOPS) buffer (pH 7.0), and resuspended in the MOPS buffer at OD600 of 1.0 (∼109 CFUs). The bacterial cells were then incubated with cytochrome c (50 μg/ml) for 15 min and pelleted; the amount of cytochrome c unbound to cell surfaces in the supernatant was determined by measuring OD530. The more unbound cytochrome c in the supernatant indicates the more positively charged the cell surface. Three independent cytochrome c binding assays were performed on separate days.
DAP binding analysis
Whole cell DAP binding assays were performed as described before.9,19 Briefly, ∼108 CFUs of each strain were exposed to 5 μg/ml of DAP for 10 min, centrifuged, and then supernatants were analyzed for residual unbound DAP by a radial diffusion assay and standard curve technique as described previously.19,29 The amount of bound DAP (±SD) was then calculated by subtracting unbound DAP amounts from the initial 5 μg/ml of DAP. These assays were repeated at least four times for each S. aureus strain on separate days.
Statistical analyses
Data were analyzed by the Kruskal–Wallis analysis of variance test with the Tukey post hoc correction for multiple comparisons. Differences less than p-value of 0.05 were considered statistically significant.
Results
DAP and CAMP susceptibility profiles
In agreement with our prior observations,
9
Newman
p < 0.01 versus Newman wild-type (WT) strain.
p < 0.05 versus CS295L and CT345A strains, respectively.
p < 0.01 versus CS295L and CT345A strains, respectively.
DAP, daptomycin; MICs, minimum inhibitory concentrations; SD, standard deviation; SNPs, single nucleotide polymorphisms.
As shown in our previous publication,
9
disruption of mprF in the Newman parental strain resulted in significantly enhanced susceptibility to the innate host defense CAMPs, hNP-1 and LL-37. Complementation of the Newman
Population analyses
The Newman
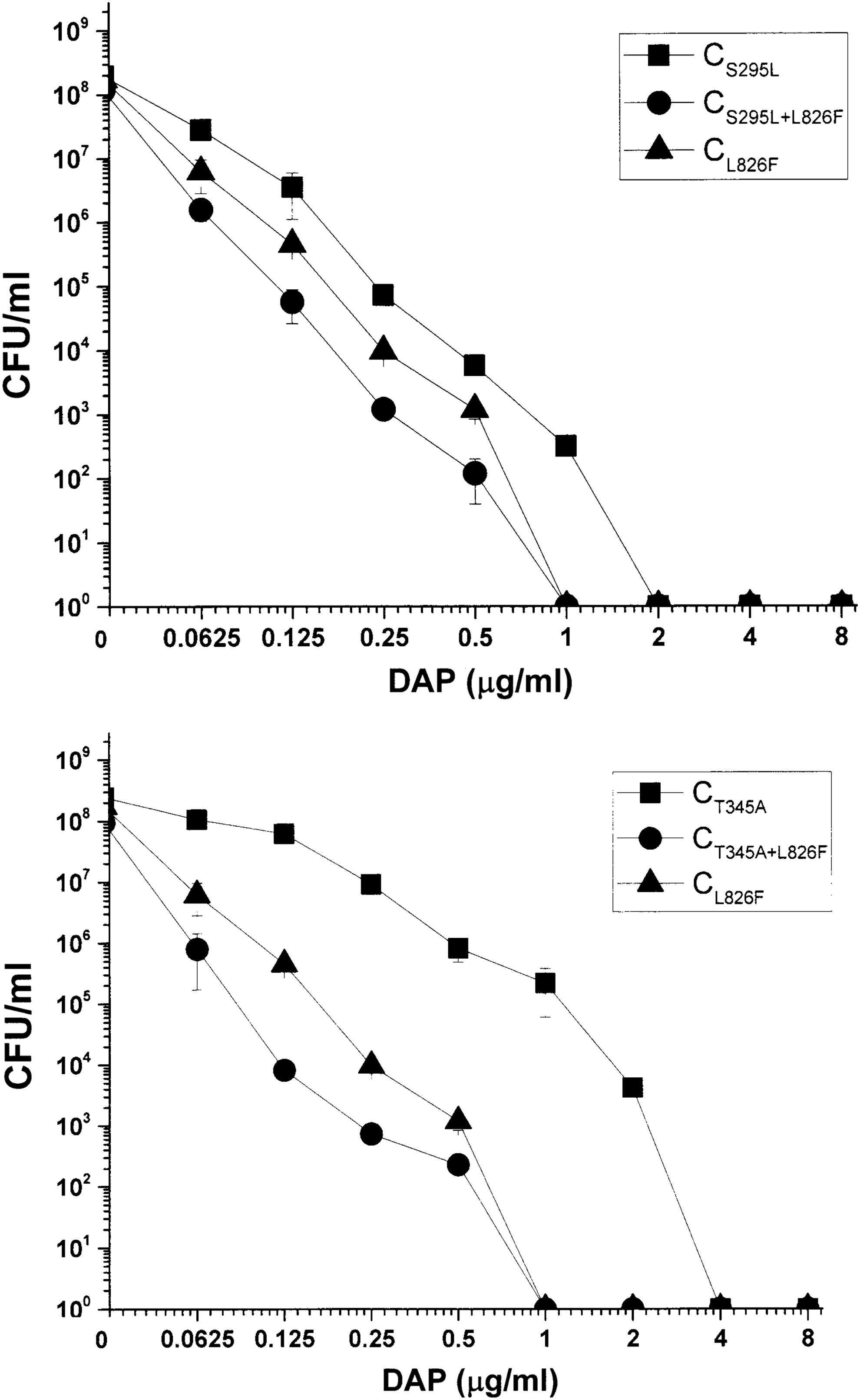
DAP population analyses of study strains. Data represent the mean ± SD for three independent experiments. DAP, daptomycin; SD, standard deviation.
Synthesis of L-PG
L-PG content of both SNP constructs was at near-parental levels. In contrast, the L-PG content was not detectable in the two strains expressing dual point–mutated mprF genes (Table 3). Interestingly, the two strains expressing dual point–mutated mprF genes showed significantly increased PG amounts, as a compensatory adaptation to the inability to produce L-PG.
p < 0.05 versus Newman wild-type (WT) strain, CS295L, and CT345A strains, respectively.
L-PG, lysyl-phosphatidylglycerol; ND, not detectable; PG, phosphatidylglycerol; PL, phospholipid.
Net surface positive charge
As anticipated, the CS295L+L826F and CT345+L826F strains had a significantly decreased surface positive charge compared to the single point–mutated CS295L and CT345A strains, respectively (Fig. 2). These reductions of surface positive charge for the CS295L+L826F and CT345+L826F strains were well correlated with the nondetectable level of L-PG in the two dual SNP variant constructs (Table 3).
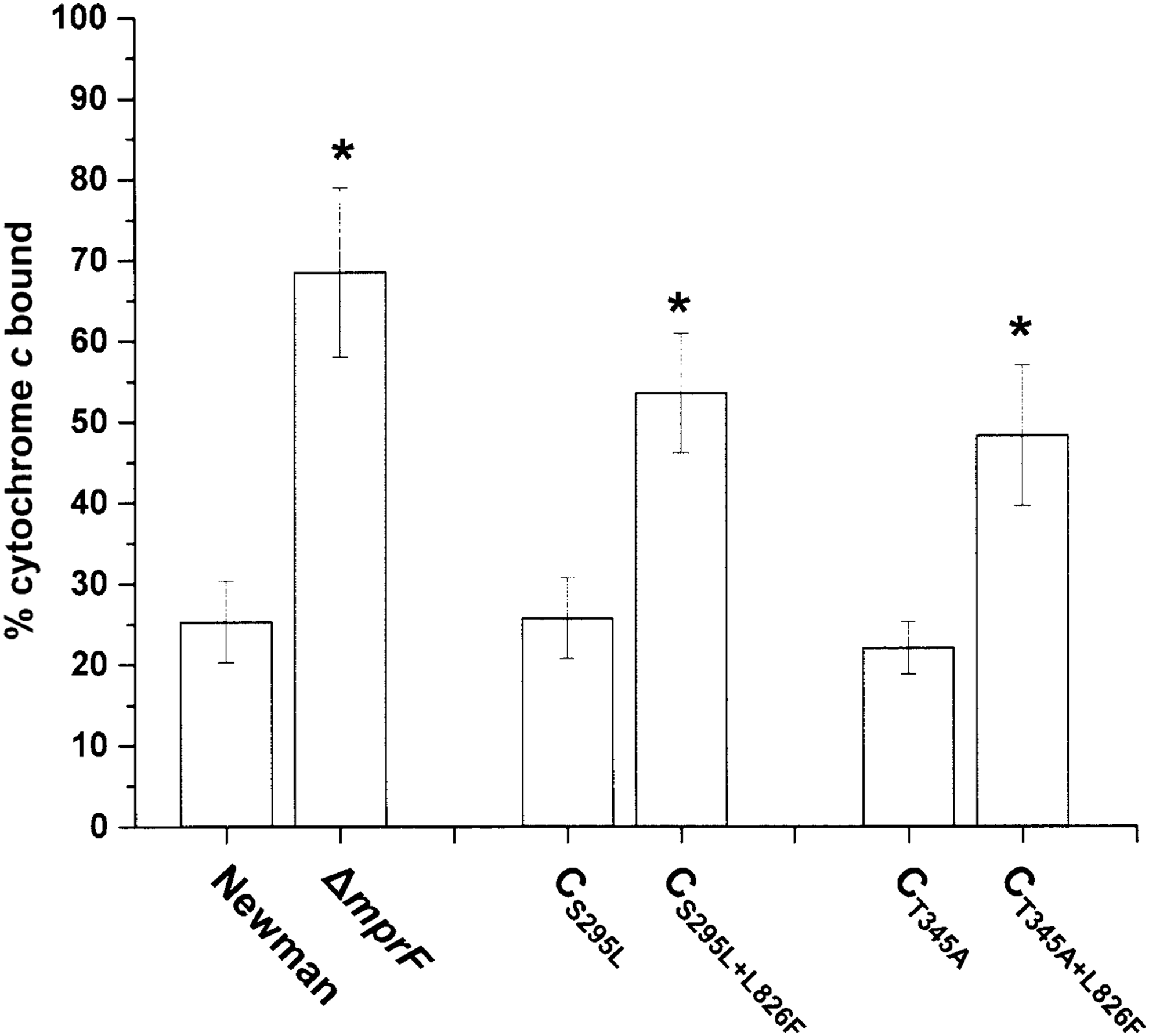
Cytochrome c binding analyses for determination of staphylococcal net positive surface charge. Percent of cytochrome c bound after 15 min of incubation with Staphylococcus aureus strains at room temperature was shown. Data are depicted as the mean ± SD from three independent assays. *p < 0.01.
DAP whole cell binding
Mirroring our prior investigation,
9
the Newman
Discussion
Although several adaptive changes have been proposed to explain the DAP-R phenotype in S. aureus and other Gram-positive pathogens, 31 the mechanisms of DAP-R still remain incompletely understood. There have been a number of recent reports, including ours, indicating the presence of SNPs within the mprF. ORF is associated with a gain-in-function phenotype in terms of DAP-R in S. aureus.1,2,4,9,10,12,19,32 These SNPs have mainly been observed in previously defined “hot spots” within the mprF ORF, usually within the central bifunctional domain or the C-terminal synthase domain of this protein.1,2 These SNPs have generally resulted in enhanced L-PG synthesis and/or translocation, with resultant modifications in surface positive charge modifications.1,2,9,12,19 Among the hot spot point mutations within the mprF ORF, the S295L, T345A, and L826F SNPs (S295L and T345A in the bifunctional domain; L826F in the C-terminal domain) were most frequently observed among DAP-R S. aureus strains in association with excess production of L-PG.1,2,9,10,12,19 However, no clinically derived or laboratory-derived DAP-R S. aureus strains have been reported to concomitantly contain two or more such hot spot SNPs in correlation with increased DAP MICs in the DAP-R range (≥2 μg/ml).
In this current investigation, we used a previously well-characterized in trans plasmid complementation strategy to express dual point–mutated forms of mprF ORFs (one in the bifunctional domain [S295L or T345A] and the other in the synthase domain [L826F]) within the Newman
The incorporation of dual point–mutated form of mprFs (CS295L+L826F and CT345+L826F) into the plasmid system yielded significantly decreased DAP MICs below the Newman parental level (0.5 μg/ml). In contrast, and in agreement with a previous publication, 9 single point–mutated form of mprFs (CS295L, CT345, CL826F) caused an increase in DAP MICs (Table 1). Correlating with the MIC results, there were significant leftward shifts in DAP-R population analyses in the two dual point–mutated constructs compared to the each single point–mutated form of mprFs (Fig. 1). These data suggest that, although individual mprF hot spot point mutations can cause the DAP-R phenotype, accumulation of these SNPs within mprF in a single strain does not confer further elevation of DAP-R, but in fact paradoxically renders such isolates more DAP-S.
Next, PL profile analyses revealed that the proportional amount of L-PG synthesized was decreased to essentially undetectable levels in the dual point–mutated CS295L+L826F and CT345+L826F complemented strains. We speculate that the accumulation of multiple hot spot point mutations within mprF ORFs may have caused loss of synthase activity of MprF. Furthermore, the inability to synthesize L-PG in the dual SNP complemented strains well correlated with decreased net positive surface charge (Fig. 2) and enhanced DAP binding in those strains (Table 1). These data support the notion that a “charge repulsion” mechanism for the DAP-R phenotype in the single point–mutated mprF is likely in-play.9,13,16,19,33
Recent publications from our group and others have indicated that DAP-R MRSA or MSSA strains frequently display cross-resistance to prototypical host defense CAMPs.1,2,15,19 Correlating with the L-PG synthesis and net surface positive charge data, the dual SNP-complemented strains displayed enhanced susceptibilities to the host defense CAMPs, hNP-1 and LL-37, versus the SNP-complemented strains, respectively. These data indicate that, at least in part, reduced net surface positive charge, as a result of loss in L-PG synthesis, is responsible for the reduced hNP-1 and LL-37 susceptibilities in the dual SNP-complemented strains.
We recognize that there are limitations in the current studies, including: (1) the limited number of combinations of mprF SNPs tested; (2) evaluating only one single MSSA parental strain, and no MRSA background isolates; (3) use of a plasmid-based complementation approach rather than chromosomal mprF point mutation constructs; and (4) since the plasmid used in our studies only expresses during exponential growth, this may well underlie the relatively blunted PL and surface charge responses seen in our single mprF mutant constructs compared to the parental strain. This latter phenomenon has been previously noted by our group. 9 Collectively, these limitations are currently being addressed in our laboratory.
In combination with previously published results, 9 our data suggest that: (1) individual hot spot point mutations within mprF gene can recapitulate the DAP-R phenotype observed in clinically-derived DAP-R donor strains through gain-in-function mechanisms (i.e., increased L-PG synthesis and net surface positive charge); and (2) accumulation of multiple hot spot point mutations results in decrease of L-PG synthase activity, which likely reduces DAP MICs caused by mitigation of surface charge repulsion of calcium-DAP. Our current findings are in line with those of Friedman et al. 3 exhibiting that the selective pressure of serial in vitro growth in sublethal concentrations of DAP results in single, but not multiple, mprF SNPs. Our data may also explain, at least in part, why DAP MICs among DAP-R S. aureus clinical isolates are never very high (i.e., >4 μg/ml). If such cumulative SNPs would have been additive, high-level DAP-R isolates would have likely been observed.
Our data leave open the possibility that clinical S. aureus strains exposed to DAP in vivo may well accumulate multiple mprF point mutations, while remaining “DAP-S” by MIC testing. Such strains would not be identified easily in the microbiology laboratory, and being DAP-S, such isolates would likely be readily cleared from infection sites by DAP. It would now be interesting to reexamine clinical strains from patient receiving DAP therapy for emergence of multiple hot spot SNPs in mprF.
The mechanism(s) by which multiple mprF SNPs lead to reduced DAP MICs and other phenotypic perturbations remain to be defined. This scenario could represent a “fitness or metabolic burden” on the organism. In contrast, it is conceivable that compensatory mutations in other genes previously associated with DAP-R (e.g., yycFG; rpoBC; or vraSR32,34,35) could also be playing a role.
Footnotes
Acknowledgments
This study was supported by the Basic Science Research Program of the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2015R1D1A1A01057442 to S.J.Y.) and the Chung-Ang University Graduate Research Scholarship (to K.M.K.) in 2016. This research was also supported by a grant RO-1 AI-39108-18 (to A.S.B.) from the National Institutes of Health.
Disclosure Statement
No competing financial interests exist.