
Abstract
Transmission of tigecycline-nonsusceptible pathogenic Enterobacteriaceae from companion animals to human should be a concern because tigecycline is a last-line drug for treating multidrug-resistant Enterobacteriaceae in human medicine. However, tigecycline susceptibility of Enterobacteriaceae isolated from companion animals has not been investigated. In this study, we investigated the tigecycline susceptibility of Klebsiella pneumoniae complex and Escherichia coli isolates from dogs and cats, and evaluated their human pathogenicity potential. Tigecycline susceptibility of K. pneumoniae, including Klebsiella quasipneumoniae (n = 86) and E. coli (n = 100) strains isolated from dogs and cats was investigated. The antimicrobial susceptibility, capsular serotype, multilocus sequence type, ompK36 group, presence of virulence genes, and serum resistance of tigecycline-nonsusceptible isolates were evaluated. All E. coli isolates were susceptible to tigecycline. Two K. pneumoniae (minimum inhibitory concentration [MIC], 4 mg/L) and one K. quasipneumoniae (MIC, 8 mg/L) isolates were tigecycline resistant. Sixteen K. pneumoniae and one K. quasipneumoniae isolates were tigecycline intermediate (2 mg/L). All tigecycline-nonsusceptible isolates (n = 20) were also ciprofloxacin nonsusceptible. These isolates harbored five to nine virulence genes; 16 isolates were resistant to the human serum. In addition, STs of 13 K. pneumoniae isolates were reported to be found in strains isolated from human; isolates considered high-risk clones in human (ST11, ST15, and ST147) were also identified. In conclusion, the isolation of tigecycline-nonsusceptible K. pneumoniae from companion animals is an impact from the viewpoint of One Health approach to antimicrobial resistance that companion animals are a reservoir of human pathogenic lineages.
Introduction
T
We recently identified 7 tigecycline-nonsusceptible isolates among 471 E. coli clinical isolates; 6 of the 7 isolates belonged to the major extraintestinal pathogenic lineages, O25b:H4-ST131 and O1-ST648 that are isolated from patients, including those with urinary tract and blood stream infections, in many countries.5,7,8 Notably, these major pathogenic lineages are frequently coresistant to other antimicrobial agents.5,8 Some of these lineages produce the New Delhi metallo-β-lactamase and are resistant to most β-lactam antimicrobials, in addition to fluoroquinolones and aminoglycosides.9,10 For the time being, these isolates remain susceptible to colistin and tigecycline.9,10 Nevertheless, we recently reported several O25b:H4-ST131 isolates that exhibited colistin resistance. 11
Although it was thought that tigecycline resistance does not result in crossresistance to any antimicrobials, including tetracyclines, because of its designed unique backbone,12,13 we recently demonstrated that acquisition of tigecycline nonsusceptibility is associated with the development of fluoroquinolone resistance in E. coli. 5 Some reports describe the isolation of tigecycline-nonsusceptible K. pneumoniae isolates resistant to β-lactams, including carbapenems, fluoroquinolones, and amikacin,6,14 which are indicators of multidrug resistance. These findings emphasize the difficulty faced by clinicians when choosing an appropriate antimicrobial therapy, and the notion that the choice is nonexistent if these resistances are accumulated.
Companion animals are part of our daily life. Dogs and cats especially have a very close contact with human, e.g., indoors. Thus, infected and colonized bacteria might transfer between human and companion animals by two-way. The animals sometimes suffer from bacterial infectious diseases, such as pneumonia, urinary tract infections, and blood stream infections, and receive antimicrobial treatments at veterinary hospitals.
Antimicrobial resistance might emerge in the animal hosts in the same manner as in a human population. Many studies reported the isolation of antimicrobial-resistant bacteria from dogs and cats and it has been suggested that some of these isolates are shared with human.15,16
Our previous study showed that antimicrobial-resistant E. coli isolates possessing several virulence genes for humans were present in urogenital specimens from infected companion animals. 17 Another study revealed that multidrug-resistant extended-spectrum β-lactamase (ESBL)-producing K. pneumoniae isolates of lineages common to those found in humans were present in several clinical specimens (such as urine, skin, genitals, respiratory organs, and ears) of companion animals. 18 These findings suggest that E. coli and K. pneumoniae clinical isolates derived from companion animals pose a potential risk to human health because of their pathogenicity and antimicrobial resistance.
Considering the current value of tigecycline in the management of human infectious disease, it would be alarming if tigecycline-nonsusceptible human pathogenic lineages emerged in companion animals and transferred to human. Nevertheless, this has never been investigated. In the current study, we investigated the tigecycline susceptibility and coresistance of K. pneumoniae and E. coli to general antimicrobials (including fluoroquinolones) using clinical isolates collected from dogs and cats.17,18 The presence of virulence genes and human serum resistance was also examined, to estimate the pathogenicity potential of these isolates in human from the viewpoint of One Health approach to antimicrobial resistance.
Materials and Methods
Bacterial isolates
K. pneumoniae (n = 86) and E. coli (n = 100) isolates were from clinical specimens from 136 dogs and 50 cats obtained in the years 2003–2016 at veterinary clinics throughout Japan that had been described in previous studies17,18 and by Harada et al. (unpublished results). Among K. pneumoniae isolates, 40 were fluoroquinolone susceptible (ciprofloxacin minimum inhibitory concentration [MIC]: ≤0.5 mg/L) and 46 were fluoroquinolone nonsusceptible (ciprofloxacin MIC: >0.5 mg/L); among E. coli isolates, 39 were fluoroquinolone susceptible and 61 were fluoroquinolone nonsusceptible.
K. pneumoniae complex strains were allocated to species (KpI, K. pneumoniae; KpII, Klebsiella quasipneumoniae; and KpIII, Klebsiella variicola) on the basis of the phylogeny of rpoB sequences. 19 Unless otherwise stated, K. pneumoniae refers to all these phylogenetic groups. These isolates were isolated from various clinical specimens (urine, uterus, vagina, pus, and kidney for E. coli; urine, nasal cavity, nasal discharge, pus, skin, uterus, prostate, genitalia, surgical incision, bronchus, lung, oral cavity, gingiva, abdominal dropsy, gastric mucosa, pleural effusion, blood, and pad for K. pneumoniae), as mostly described in previous studies.17,18
This study was carried out in accordance with the recommendations of Guidelines for Proper Conduct of Animal Experiments of the Science Council of Japan.17,18 Bacterial strains were isolated as a part of routine veterinary examination, and did not include any more invasive operation against the companion animals (dogs and cats).17,18
Susceptibility testing
Strain susceptibilities to tigecycline (AdooQ BioScience, Irvine, CA) and amikacin (Wako Pure Chemical, Tokyo, Japan) were tested using the agar plate dilution method. 20 Antimicrograms for other agents were from previously published studies.17,18 E. coli ATCC25922 was used as a reference. The MIC results were interpreted according to the EUCAST breakpoints. 21
Genetic analysis
Multilocus sequence typing (MLST) was performed according to a protocol available on the website of Institute Pasteur (Paris, France) (http://bigsdb.web.pasteur.fr/klebsiella/klebsiella.html). The ompK36 groups were determined by PCR, as described previously. 22 The presence of 14 virulence genes (allS, entB, fimH, iutA, kfuBC, magA, mrkD, rmpA, uge, ureA, wabG, wcaG, and ybtS) was determined by PCR, as previously described.23,24 The major capsular serotypes that are also associated with invasive infections 25 (K1 and K223; and K5, K20, K54, and K5725) were determined by PCR targeting the capsular serotype-specific gene wzi. Each positive strain in our bacterial collection was used as a control after nucleotide sequence verification.
Serum resistance assay
Serum resistance was determined as previously described. 26 Briefly, each strain was inoculated (1 × 105 CFU/ml) into a pooled human complement solution (Cedarlane, Burlington, Canada) diluted (1:3, v/v) in cation-adjusted Mueller Hinton broth II (Becton Dickinson, Franklin Lakes, NJ), and incubated at 37°C for 4 hr. After incubation, the suspension was spread on Mueller Hinton agar (Becton Dickinson), and incubated overnight at 37°C. The strains that grew on the Mueller Hinton agar were defined as serum resistant. K. pneumoniae ATCC13883 was used as a serum-susceptible control.
Results
Tigecycline susceptibility of K. pneumoniae and E. coli isolates from dogs and cats
The distribution of tigecycline MIC values in K. pneumoniae isolates was shifted toward higher values than in E. coli isolates (Fig. 1).
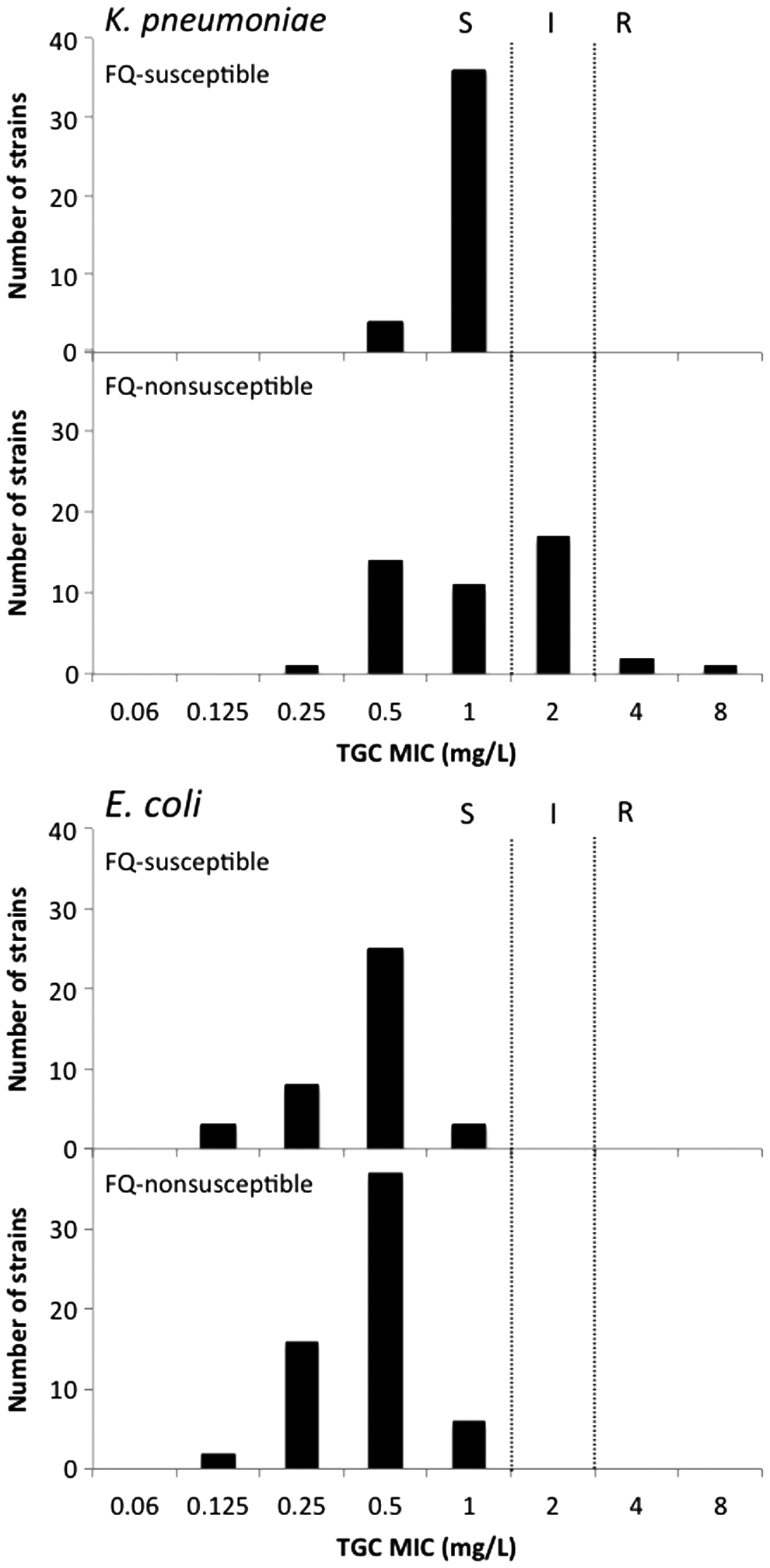
Distribution of tigecycline MIC values in clinical Klebsiella pneumoniae and Escherichia coli isolates from dogs and cats. FQ, fluoroquinolone; MIC, minimum inhibitory concentration; TGC, tigecycline; I, intermediate; R, resistant; S, susceptible.
In K. pneumoniae, MIC50 values for tigecycline were 1 mg/L in both fluoroquinolone-susceptible and -nonsusceptible isolates; MIC90 values were 1 and 2 mg/L in those isolates, respectively. Three isolates were tigecycline-resistant (tigecycline MIC of 4 mg/L for strains KL54 and KL118; and tigecycline MIC of 8 mg/L for strain KL34); 17 isolates were defined as “intermediate” (tigecycline MIC of 2 mg/L). All (n = 20) tigecycline-nonsusceptible K. pneumoniae isolates were also fluoroquinolone nonsusceptible (ciprofloxacin MIC ranging from 0.5 to 256 mg/L). All tigecycline-nonsusceptible isolates were also susceptible to amikacin (amikacin MICs were 0.5 or 1 mg/L).
All E. coli isolates were susceptible to tigecycline (Fig. 1). The tigecycline MIC50 (0.5 mg/L for both fluoroquinolone-susceptible and -nonsusceptible isolates) and MIC90 (0.5 and 1 mg/L for fluoroquinolone-susceptible and -nonsusceptible isolates, respectively) were twofold lower than those of K. pneumoniae.
MLST, capsular serotypes, and ompK36 groups of tigecycline-nonsusceptible isolates
We next determined MLST STs, capsular serotypes, and ompK36 groups of the 20 tigecycline-nonsusceptible K. pneumoniae isolates (Table 1). Among these, 11 isolates were previously shown to be ESBL positive; their STs have been identified as ST15, ST655, and ST1844 (n = 2 of each); and ST881, ST147, ST337, ST709, and ST2173 (n = 1 of each). 18 We determined STs for seven other tigecycline-nonsusceptible isolates; these were ST11, ST32, ST15, ST314, and ST655 (n = 1 of each), and ST1043 (n = 2). The remaining two isolates (KL34 and KL107), with no MLST assignment, belonged to the phylogenetic group KpII, K. quasipneumoniae. 19
These sequence types were determined in a previous study. 18
KL34 and KL107 strains are K. quasipneumoniae.
+, serum-resistant; −, not resistant; I, intermediate; R, resistant; MIC, minimum inhibitory concentration.
Only K2 and K54 capsular serotypes were detected in one and two isolates, respectively (Table 1). Furthermore, the examined tigecycline-nonsusceptible K. pneumoniae isolates belonged to any of the four major ompK36 groups (A–D).
Virulence genes harbored by tigecycline-nonsusceptible isolates
Tigecycline-nonsusceptible K. pneumoniae isolates harbored 5–9 of the 14 virulence genes (Table 1).
fimH (encoding type I fimbriae), mrkD (encoding type III fimbriae), wabG (with a role in the synthesis of the lipopolysaccharide core) ureA (encoding a urease gamma subunit), and entB (siderophore) were detected in all isolates. uge (encoding UDP-galacturonate 4-epimerase) was detected in all but one of the isolates. kfuBC (encoding an iron transport/phosphotransferase function) was detected in half of the isolates. Genes encoding siderophores, ybtS and iutA, were detected in individual four K. pneumoniae isolates. allS (encoding a regulator of the allantoin regulon) and rmpA (regulator of mucoid phenotype A) were detected in each of the isolates. wcaG (capsular fucose synthesis) was detected in two K. pneumoniae isolates. magA (mucoviscosity-associated gene A; marker of serotype K1) was not detected.
Serum resistance
Eighty per cent (16/20) of the tigecycline-nonsusceptible strains were serum resistant (Table 1). Two of the four serum-sensitive strains were K. quasipneumoniae.
Discussion
In the current study, we investigated tigecycline susceptibility of K. pneumoniae and E. coli isolates from dogs and cats. The distribution of tigecycline MIC toward higher values in K. pneumoniae than in E. coli (Fig. 1) was similar to what had been observed in human clinical isolates in a previous large-scale surveillance study. 3 These results suggest that tigecycline susceptibility displays almost the same situation in Enterobacteriaceae clinical isolates derived from human and from companion animals, with K. pneumoniae being less susceptible to tigecycline than E. coli.
We identified 20 tigecycline-nonsusceptible K. pneumoniae isolates, including 2 K. quasipneumoniae isolates; no tigecycline-nonsusceptible E. coli isolates were identified. Hence, these K. pneumoniae complex isolates may be concerned if they also exhibit multidrug resistance. All tigecycline-nonsusceptible isolates were susceptible to amikacin and meropenem. 18 This suggested that these isolates are not multidrug-resistant, and the infections that they cause might still be treated using these antimicrobials.
In Japan, tigecycline has never been used in the veterinary field. The tigecycline-nonsusceptible isolates from companion animals should have therefore developed, and have been selected, without any exposure to tigecycline. Our previous study revealed that the emergence of tigecycline nonsusceptibility is clearly associated with the development of fluoroquinolone resistance in E. coli clinical isolates from human. 5 Likewise, in the current study, tigecycline-nonsusceptible K. pneumoniae were detected only in the fluoroquinolone-nonsusceptible population (Fig. 1).
Another study reported that the prevalence of tigecycline nonsusceptibility in ESBL-positive clinical isolates of K. pneumoniae from human was two times higher than in entire K. pneumoniae isolates. 3 According to our previously published antibiograms, 18 the cefotaxime- and gentamicin-nonsusceptibility rates for K. pneumoniae complex strains derived from dogs and cats were 39.3% and 25.8%, respectively, and those for the tigecycline-nonsusceptible isolates are 60.0% and 55.5%, respectively. This suggested that nonsusceptibility rates to cephalosporins and gentamicin in tigecycline-nonsusceptible isolates were higher than in the entire K. pneumoniae complex population.
On the other hand, cononsusceptibility to tigecycline and fluoroquinolones is higher than cononsusceptibility to tigecycline and cephalosporin/gentamicin in K. pneumoniae complex. Although antimicrobial treatment histories of the dogs and cats were not available, tigecycline-nonsusceptible K. pneumoniae might emerge during treatment with veterinary antimicrobials, especially with fluoroquinolones. The previously mentioned report revealed a higher prevalence of fluoroquinolone nonsusceptibility in clinical isolates from dogs and cats (46%) 18 than in human clinical isolates (3.5–23.4%).3,4,27 Based on the higher occurrence of fluoroquinolone nonsusceptibility, it may be deduced that the occurrence of tigecycline-nonsusceptible K. pneumoniae in dogs and cats is much more frequent than in humans (3.5–4.7%).3,4
Transmission of tigecycline-nonsusceptible isolates from companion animals to human would be concerning if these isolates would also be pathogenic to humans. In the current study, multiple virulence genes contributing to infection in human, such as urinary tract infections, pneumonia, and blood stream infections, were noted in the tigecycline-nonsusceptible K. pneumoniae isolates from dogs and cats (Table 1). All tigecycline-nonsusceptible isolates possessed virulence genes, those were mostly found in K. penumoniae/K. quasipneumoniae isolates, such as fimH, mrkD, wabG, and ureA.23,24,28 An essential gene of capsule synthesis, uge, which is associated with urinary tract infections, pneumonia, and septicemia, 29 was detected in all but one of isolates. In addition, most of the tigecycline-nonsusceptible K. pneumoniae isolates were resistant to serum that is required for resistance to complement-mediated killing and establish blood stream infections. 26
Although two of the serum-sensitive tigecycline-nonsusceptible isolates were K. quasipneumoniae, this organism is also capable to infect human, similarly to K. pneumoniae. 30 Thus, these tigecycline-nonsusceptible isolates should harbor pathogenicity for humans, and probably for companion animals, because these isolates were obtained from clinical specimens of the companion animals.
On the other hand, some specific serotypes, such as K1, K2, K5, K20, K54, and K57, are known to be of hypervirulent lineage, which potentially have the ability to cause severe clinical cases (liver abscesses, pneumonia, meningitis, and endophthalmitis).23,25 They possess the virulence-associated genes, such as rmpA, magA, kfuBC, allS, and/or wcaG. A gene encoding a regulator of capsular polysaccharide synthesis, rmpA, which is associated with the hypermucoviscous phenotype, 31 was detected in one isolate. Although magA was not detected, kfuBC (detected in half of isolates) and allS (in one strain), which are highly prevalent among strains derived from tissue-invasive diseases, e.g., pyogenic liver abscessed and septic complications,32,33 were detected.
Another capsular synthesis gene, wcaG, associated with the specific capsular type K1 and K54,25,34 was detected in two K54-positive isolates. For siderophores, the presence of iutA (found in four isolates) or the coexistence of widely found siderophores, entB and ybtS (found in another four strains), is clearly associated with community-acquired invasive infections. 30 Three iutA-positive isolates were hypervirulent serotypes, K2 and K54. Thus, the prevalence suggested that some of the tigecycline-nonsusceptible K. pneumoniae isolates (such as KL26, KL27, and KL53) might potentially be able to cause severe and invasive infections in human, as well as in companion animals. It is a large impact from the viewpoint of One Health approach to antimicrobial resistance that companion animals are a reservoir of human pathogenic tigecycline-nonsusceptible K. pneumoniae.
The tigecycline-nonsusceptible isolates were isolated from various sources (Table 1). Direct transmission of the tigecycline-nonsusceptible isolates from sources, such as nasal discharge, skin, gingiva, and urine, could occur easily between companion animals and humans during daily close contacts.
Isolates from other sites (such as uterus and genitalia) could also be transferable to human. Because these sources are generally aseptic, the isolation of the tigecycline-nonsusceptible K. pneumoniae from these sources indicates that they must cause allopatric infections and capable of invasion. Invasive strains may colonize or infect commensal organs of companion animals, which readily make contact with human, and thereby facilitate the transfer of tigecycline-nonsusceptible isolates to human. Although the actual frequency of transmission between companion animals and humans is still unclear, the potential risk of transmission would be higher than that of other animals, such as livestock and wild animals.
STs of the tigecycline-nonsusceptible isolates from companion animals comprised some lineages (ST11, ST15, ST32, ST147, ST337, ST655, ST709, and ST1043) that have been seen in human isolates.18,35–40 Remarkably, 5 of the 20 tigecycline-nonsusceptible isolates (25%) were international high-risk clones (ST11, ST15, and ST147), spreading epidemically worldwide, with multidrug resistance and invasive pathogenicity in human.22,39,41 Recently, Ovejero et al. reported the first identification of two tigecycline-resistant K. pneumoniae isolates, which were ST11 and ST147, from dogs in Spain. 42 Thus, this study indicates that some of the high-risk pathogenic K. pneumoniae lineages with tigecycline nonsusceptibility could spread among humans and dogs worldwide.
Among the tigecycline-nonsusceptible international high-risk clones, one ST11 isolate harbored six virulence genes, whereas three ST15 isolates and one ST147 isolate each harbored seven virulence genes. These isolates were sensitive to some antimicrobials, e.g., amikacin and carbapenems. 18 Two of the ST15 isolates and the ST147 isolate had been identified as ESBL positive and coresistant to an aminoglycoside, gentamicin. 18 Currently, human isolates of these STs exhibit resistance to these agents, and to colistin.39,43,44 These results indicate that these STs have an ability to accumulate antimicrobial resistance.
The risk to human health would be extremely high if these tigecycline-nonsusceptible strains acquired additional resistance mechanisms in the future. To prevent the worst possible scenario, i.e., the spread of human pathogenic multidrug-resistant K. pneumoniae strains with tigecycline resistance, appropriate antimicrobial use and continuous surveillance of tigecycline-nonsusceptible K. pneumoniae strains should become as important in the veterinary field as in the human clinical field.
Even though international high-risk clones were identified among tigecycline-nonsusceptible strains from companion animals, it should be noted that the isolation of these clones has been fewer reported in Japan.45,46 The human clinical Japanese K. pneumoniae isolates seem to show different susceptibility trends than international isolates, and may have different trends from the Japanese veterinary isolates. Nevertheless, MLST data for K. pneumoniae clinical isolates in Japan are insufficient to draw any definitive conclusions on the subject. Further studies are required to reveal the actual interaction of tigecycline-nonsusceptible K. pneumoniae from human and companion animals.
Conclusion
We found that companion animals harbor tigecycline-nonsusceptible K. pneumoniae strains, including international high-risk clones (ST11, ST15, and ST147) with potential pathogenicity to human. We also observed a strong association between tigecycline nonsusceptibility and fluoroquinolone nonsusceptibility of these microorganisms. Measures, such as antimicrobial stewardship, should be taken in the companion animal clinical field from the viewpoint of One Health approach to antimicrobial resistance, as well as in the human clinical field, to prevent the spread of human pathogenic tigecycline-nonsusceptible K. pneumoniae isolates and the development of multidrug resistance in these isolates.
Footnotes
Acknowledgments
Disclosure Statement
No competing financial interests exist.