
Abstract
This study was conducted to assess emergence of daptomycin-nonsusceptible (DAP-NS) phenotype in DAP-naive patients with invasive Staphylococcus aureus (ISA) infections in Korea. A total of 208 S. aureus clinical isolates were selected from a previous prospective study on ISA infections and evaluated for DAP-NS. Although DAP has never been introduced in Korea, five DAP-NS S. aureus strains (2.4%) were identified among 208 S. aureus strains collected from ISA infections. The DAP-NS phenotype was observed only in methicillin-resistant S. aureus (MRSA) strains, but not in methicillin-susceptible S. aureus strains. One DAP-NS MRSA strain belonged to sequence type 72 (ST72) and four were ST5 MRSA strains, three of which were heteroresistant vancomycin (VAN)-intermediate S. aureus. All these five DAP-NS MRSA strains were from healthcare-associated infections without prior exposure to VAN within 30 days. While the ST72 MRSA strain exhibited DAP-NS phenotype via charge repulsion mechanism, four ST5 DAP-NS S. aureus strains had charge-independent DAP-NS mechanism. None of the five DAP-NS strains displayed significant increase in cell wall thickness, indicating that altered cell wall thickness was not associated with the observed DAP-NS phenotype.
Introduction
D
The detailed mechanisms involved in the loss of DAP susceptibility (DAP-S) have not been clarified yet. High burden of infecting organisms, suboptimal antibiotic exposure, and poor antibiotic penetration into infection sites have been suggested as factors associated with DAP-NS. 10 Single-nucleotide polymorphisms (SNPs) within multiple peptide resistance factor F (mprF) locus responsible for lysinylation of phosphatidylglycerol (PG) and thus increased surface positive charge in S. aureus are a plausible mechanism of DAP-NS in some clinical S. aureus strains isolated from patients with failed DAP therapy.11,12 However, the mechanism involved in the resistance to DAP in S. aureus strains isolated from patients without previous exposure to DAP remains unclear.
DAP has not been commercially available in Korea, where substantial numbers of invasive S. aureus (ISA) infections occur with high rate of methicillin resistance. 13 Previous studies have indicated that VAN treatment exposures in patients may be an important preconditioning event for subsequent emergence of DAP-NS S. aureus during DAP treatment.6,14 However, clinical MRSA strains collected during 2004–2009 at a tertiary referral medical center in Korea showed no DAP-NS, although the strains were screened for elevated DAP minimal inhibitory concentrations (MICs) according to their VAN MIC values (≤1 μg/ml vs. >1 μg/ml). 15
In the current study, we conducted a multicenter investigation in a large scale to assess emergence of DAP-NS among three groups of clinical S. aureus strains isolated from patients with ISA infections who had no prior exposure to DAP: (1) heteroresistant VAN-intermediate S. aureus (h-VISA); (2) S. aureus isolates with VAN MICs of >1.5 μg/ml; and (3) S. aureus isolates that caused persistent bacteremia ≥7 days. Furthermore, to characterize potential mechanisms involved in increased DAP resistance, SNPs within mprF, expression profiles of mprF and dltABCD transcription, surface positive charge, and cell wall thickness were assessed in DAP-NS MRSA strains identified.
Materials and Methods
Bacterial strains
Potential DAP-NS isolates were selected from S. aureus collection obtained in a previous prospective study on ISA infections performed from July 2009 to June 2011 at 10 hospitals located in Korea. 16 ISA infection was diagnosed when S. aureus was isolated from a normally sterile body fluid as defined previously. 17 Detailed clinical information on each ISA case was prospectively collected. Potential DAP-NS S. aureus isolates were defined as follows: (1) h-VISA confirmed by the modified population analysis profile-area under the curve methods as described in the previous study, 16 (2) S. aureus strain with VAN MIC more than 1.5 μg/ml by Etest (bioMérieux) or broth microdilution (BMD), and (3) persistent S. aureus bacteremia for 7 days or more.
S. aureus strains were grown in either Tryptic Soy Broth (TSB; Difco Laboratories, Detroit, MI) or Meuller–Hinton broth (MHB; Difco Laboratories) depending on individual experiments. Broth cultures were grown in Erlenmeyer flasks at 37°C with shaking (225 rpm) in a volume less than 10% of the flask volume.
Determination of VAN and DAP MICs
For all S. aureus isolates, VAN MICs were, respectively, determined by BMD method according to Clinical and Laboratory Standards Institute (CLSI) guidelines and by Etest (bioMérieux) according to the manufacturer's instructions.18,19 To identify potential DAP-resistant S. aureus isolates, DAP MICs were determined by using Etest (bioMérieux). Etest was performed using 0.5 McFarland standard inoculum streaked onto MHA plates. VAN and DAP Etest strips were placed on these plates. DAP-NS strains were defined when MIC was more than 1.0 μg/ml. S. aureus strains ATCC 29213 and Mu50 were used as quality control strains. Clinical and microbiological characteristics were collected for DAP-NS isolates.
DNA isolation and sequencing
Genomic DNA was isolated from S. aureus strains, using method as described previously. 20 PCR amplification of mprF open reading frames (ORFs) was performed using mprF gene-specific primer pair as described previously. 21 DNA sequencing of mprF ORFs was performed at Cosmo Genetech (Seoul, Korea). Multiple mprF sequence alignments were done using BoxShade Server (www.ch.embnet.org/software/BOX_form.html).
Molecular typing and sequencing
Multilocus sequence typing and staphylococcal cassette chromosome mec (SCCmec) typing were performed as described previously.22,23 Protein A gene (spa) type was also determined as described previously. 24 Sequencing of spa gene amplicon was performed at Cosmo Genetech. Sequence of the spa gene was analyzed using Ridom StaphType software (http://SpaServer.ridome.de).
RNA isolation and quantification of transcript levels by quantitative real-time PCR
For RNA isolation, fresh overnight cultures of S. aureus strains were used to inoculate TSB to optical density at 600 nm (OD600) of 0.1. Cell pellets were harvested during both exponential growth (2.5 h) and stationary phase (12 h). Total RNA was isolated from cell pellet using RNeasy Kit (Qiagen, Valencia, CA) and FASTPREP FP120 instrument (BIO 101, Vista, CA) according to manufacturer's recommended protocols.
Quantitative real-time PCR (qRT-PCR) assay was carried out as described previously.25,26 In brief, 1.5 μg of DNase-treated RNA was reverse transcribed to generate cDNAs using Moloney murine leukemia virus reverse transcriptase (New England Biolabs, Beverly, MA) and gene-specific reverse primers.21,27 Quantification of cDNA levels was performed in triplicate samples using Power SYBR Green Master Mix Kit (Applied Biosystems) with LineGene 9620 fluorescent quantitative detection system (BIOER technology). Transcript levels of mprF, dltA, and gyrB genes were detected using respective specific primers as described previously.27,28 Fold changes in expression level of mprF and dltA genes were determined in relationship to the housekeeping gene, gyrB. Transcriptional profiling was performed at both exponential and stationary growth phases. At least two independent runs in triplicate samples were performed on separate days.
Net positive surface charge
To assess the relative net positive surface charge of S. aureus strains, cytochrome c binding assay was performed as described previously.21,27,29 In brief, S. aureus cells were grown overnight in TSB, washed with 20 mM morpholinopropanesulfonic acid (MOPS) buffer (pH 7.0) four times, and resuspended in MOPS buffer to reach OD600 of 1.0. Next, cells were incubated with 0.5 mg/ml cytochrome c for 20 min and the amount of cytochrome c remaining in the supernatant was measured by determining OD530 (the absorption maximum of the prosthetic group). 30 Higher amount of unbound cytochrome c in the supernatant correlates with more positively charged bacterial surface. 30 At least three independent runs in triplicate samples were performed on separate days.
Transmission electron microscopy
To determine the cell wall thickness of the DAP-NS strains, S. aureus cells were prepared for transmission electron microscopy (TEM) analyses as described before. 31 Sample processing and TEM analyses were performed at the National Instrumentation Center for Environmental Management (NICEM), College of Agriculture and Life Sciences, Seoul National University. For each strain, 50 cell wall thickness measurements were taken from a minimum of 25 cells at × 200,000 magnification (model LIBRA 120; Carl Zeiss, Germany).
Statistical analysis
Kruskal–Wallis analysis of variance test with the Tukey post hoc correction was utilized for multiple comparisons. Statistical significance was considered when p < 0.05.
Results
Clinical and microbiological characteristics of DAP-NS S. aureus clinical isolates
A total of 1,289 S. aureus isolates from ISA infection cases at 10 different hospitals during 2 years of study period were screened. Among these, 208 S. aureus strains met the definition for potential DAP resistance, including 171 MRSA strains (82%) and 37 MSSA strains (18%). As shown in Fig. 1, among these 208 S. aureus strains that met the definition for potential DAP resistance, 124 (59%) exhibited VAN MICs of >1.5 μg/ml by Etest or BMD methods, 93 (44%) were from persistent bacteremia, and 42 (20%) were h-VISA.

Distribution of potential DAP-NS Staphylococcus aureus clinical isolates from patients with invasive infections. DAP-NS, daptomycin nonsusceptible; h-VISA, heteroresistant vancomycin-intermediate S. aureus; MICs, minimal inhibitory concentrations.
By Etest method, the mean, mode, and ranges of MICs of DAP were 0.30, 0.25, and 0.0019–2 μg/ml, respectively. DAP MIC values did not show any significant difference among the three potential DAP-NS groups (Table 1). Among 208 potential DAP-NS S. aureus strains, 5 (2.4%) MRSA strains from three hospitals showed DAP-NS phenotype (MIC >1 μg/ml). All DAP-NS strains were MRSA blood isolates from patients without previous exposure to VAN within 1 month before they were isolated. All these five DAP-NS strains were healthcare-associated MRSA (HA-MRSA) strains isolated from patients who underwent surgery within 1 year. All these patients but one had catheter-associated infections or surgical-site infections.
Some of these isolates overlapped in each group.
DAP, daptomycin; h-VISA, heteroresistant vancomycin-intermediate S. aureus; MIC, minimal inhibitory concentration; NS, nonsusceptible.
Although four of these five DAP-NS isolates showed sequence type (ST) 5-MRSA-SCCmec II genotype, they were not epidemiologically linked to each other (Table 2).
BJI, bone and joint infection; BMD, broth microdilution; CA-BSI, catheter-associated blood stream infection; CA-UTI, catheter-associated urinary tract infection; DM, diabetes mellitus; HA, healthcare associated; SSI, surgical-site infection; ST, sequence type.
mprF sequencing data
As shown in Table 3, mprF ORF sequencing revealed that mprF sequences of all four ST5 DAP-NS strains were identical to mprF sequence of S. aureus MW2 reference strain. In contrast, the mprF sequence of ST72 DAP-NS strain, 480, exhibited four amino acid substitutions (V26A, I376M, I461T, and I666I) compared with mprF sequence of S. aureus MW2, suggesting some degree of heterogeneity within mprF ORFs in ST72 type of S. aureus strains. These four mprF SNPs identified in 480 strain have not been reported previously in relationship to DAP-NS phenotype.
The mprF sequence from S. aureus MW2 was used as the consensus reference sequence.
MLST, multilocus sequence typing; mprF, multiple peptide resistance factor F; spa, staphylococcal protein A; SNPs, single-nucleotide polymorphisms.
Transcription of mprF and dltABCD
As shown in Fig. 2A and B, qRT-PCR analyses of mprF transcripts revealed that all four ST5 DAP-NS strains (481–484) did not exhibit enhanced expression of mprF compared with three ST5 DAP-S strains during exponential or stationary growth phase. In contrast, ST72 DAP-NS strain (480) exhibited significantly higher level of mprF expression than all three DAP-S strains during the stationary growth phase (Fig. 2B).

Relative transcription level of mprF during exponential
As shown in Fig. 3A, all DAP-NS strains exhibited significantly lower mean fold expression level of dltA than DAP-S group of strains during exponential growth phase. None of DAP-NS strains had significantly higher levels of dltA expression than all three DAP-S strains during the stationary growth phase (Fig. 3B).

Relative transcription level of dltA during exponential
Surface positive charge based on cytochrome c binding assay
Cytochrome c binding analyses of DAP-NS strains revealed that only one ST72 DAP-NS strain (480) had significantly enhanced surface positive charge compared with the three DAP-S strains (Fig. 4).
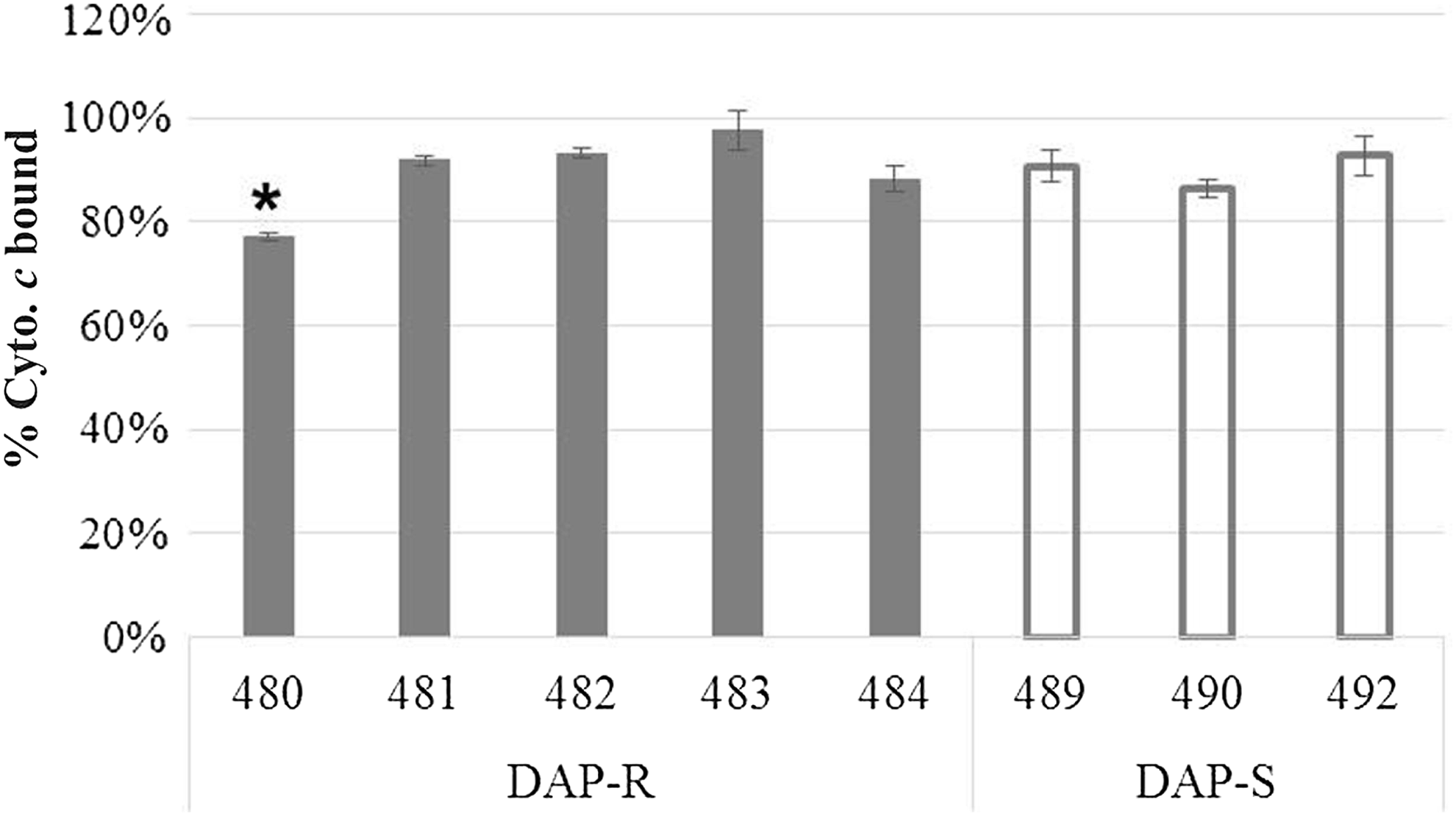
Binding of positively charged cytochrome c to Staphylococcus aureus whole cells. The graph shows percent of cytochrome c bound after 15 min of incubation with each study strain at room temperature. Three DAP-S ST5 MRSA strains were included for the assay. Data represent the means (±SD) from three independent experiments. *Lower than all the DAP-S strains tested (p < 0.01). MRSA, methicillin-resistant S. aureus; ST, sequence type.
Cell wall thickness
As shown in Fig. 5, all the five DAP-NS S. aureus strains did not exhibit increased cell wall thickness measurements compared with that of DAP-S strain (p > 0.5).

TEM analyses of the DAP-NS Staphylococcus aureus strains. The thickness of the cell walls was measured at × 200,000 magnification (mean ± SD nm). #A ST5 DAP-S MRSA strain (489) was included for the TEM analyses. TEM, transmission electron microscopy.
Discussion
VAN treatment failure or poorer outcome in invasive MRSA bacteremia caused by strains with decreased VAN susceptibility has been demonstrated in several studies.32–34 Cases of h-VISA infections have shown prolonged period of bacteremia with relatively higher mortality than VAN-susceptible S. aureus infections in our previous study. 16 DAP plays an important role as a primary therapy or a salvage strategy in treating invasive S. aureus infections, especially in cases with treatment failure. However, the possible association between reduced susceptibility to VAN and DAP-NS in S. aureus can limit its use in severe MRSA infections.6,7 Therefore, we surveyed clinical situations, in which DAP could have the highest benefit such as infections caused by h-VISA or S. aureus isolates with reduced susceptibility to VAN or persistent bacteremia cases. Results of this study revealed that DAP-NS in S. aureus isolates was not common (2.4%, 5/208) in Korea. All DAP-NS isolates showed MICs of 1.5–2 μg/ml. In regions where DAP has not been widely used, DAP can be a useful and safe choice for effective salvage treatment for invasive S. aureus infections without concern about DAP resistance.
While DAP-NS in S. aureus isolates is not a major clinical issue in Korea, the presence of DAP-NS among S. aureus isolates without any prior exposure to DAP confers a substantial importance. For the first time, we found DAP-NS clinical S. aureus isolates in Korea. We also demonstrated that DAP-NS might emerge without previous exposure to DAP. Moreover, none of those patients who was infected by DAP-NS S. aureus strains in this study had recent exposure to VAN before the diagnoses of infections at least ≤30 days, suggesting potential risk of DAP-NS development among S. aureus isolates without DAP or VAN exposure.8,9 It is possible that these MRSA isolates might be nosocomial strains present in hospitals with various antimicrobial pressure, including VAN, before infecting patients. In addition, we collected only the first S. aureus isolates for two DAP-NS strains (480 and 483) from persistent bacteremia cases. Therefore, we could not analyze the effects of long-term antimicrobial exposure in persistent S. aureus bacteremia cases, which may be potentially related to DAP-NS development. All five cases of DAP-NS S. aureus infections in this study were epidemiologically HA. Moreover, three of these five DAP-NS strains were h-VISA strains, indicating that the mechanism of DAP resistance might be associated with VAN heteroresistance in those strains. Further investigation is needed to clarify this hypothesis.
Two recent publications provided further insight into the possible mechanisms of DAP-NS phenotype among MRSA strains isolated from DAP-naive patients.35,36 Previously, it has been demonstrated that cross-resistance to DAP and host defense cationic antimicrobial peptides (HDPs) coemerge in MRSA strains isolated from patients who had never received DAP treatment. 35 In this study, two distinct DAP MIC populations of MRSA strains (<0.5 and 1 μg/ml) were segregated and were correlated with higher resistance to killing by thrombin-induced platelet microbicidal proteins (tPMPs). More recently, two MRSA isolates from rabbits who received no DAP therapy displayed substantially enhanced DAP MICs along with increased resistance to killing by two prototypical HDPs (tPMPs and hNP-1) compared with the parental MRSA strain. 36 These observations indicated that endogenous exposure of MRSA to specific HDPs in the human/animal bloodstream or target tissues can induce development of HDP resistance, as well as to DAP in the absence of prior exposure to DAP.
While four of five DAP-NS isolates belonged to ST5-MRSA-SCCmec II, the most predominant HA-MRSA, one isolate was found to be ST72-MRSA-SCCmec IV, is the most common community-associated MRSA (CA-MRSA) in Korea. 37 Antimicrobial susceptibility profiles have shown that ST72-MRSA-SCCmec IV strains are susceptible to most nonbeta-lactams in previous studies.37,38 However, these strains have increasingly resulted in HA-infections such as pneumonia and surgical-site infections as well as CA-infections in a recent study. 39 Considering profound difference in antimicrobial susceptibility between HA-MRSA and CA-MRSA strains in Korea, the mechanism of DAP-NS might be different based on their molecular characteristics, even though they have the same epidemiological background in terms of HA-MRSA.
Interestingly, in the current study, distinct SNPs within the mprF ORF were identified in ST72 DAP-NS S. aureus strain (480), resulting in multiple amino acid substitutions in both L-PG synthase domain and bifunctional domains.11,12,40 These four SNPs (V26A, I376M, I461T, and T666I) found in ST72 DAP-NS strain have not been described previously in association with increased lysil-phosphatidylglycerol synthesis/translocation or DAP-NS phenotype. Of note, this ST72 DAP-NS strain exhibited significant increase in surface positive charge compared with DAP-S strains without such SNPs, although these DAP-S strains were ST5 strains. Unlike strain 480, all four ST5 DAP-NS strains had no SNPs within mprF ORFs, suggesting that additional undefined mutations or genetic mechanisms might be involved in DAP-NS.
Previously, it has been shown that some DAP-NS S. aureus strains can express higher levels of mprF and/or dltABCD transcripts than their isogenic DAP-S parental strains during in vitro growth.12,21,28,41 However, none of the four ST5 DAP-NS strains in this study showed significant increase in levels of mprF or dltA transcription compared with ST5 DAP-S strains during both exponential and stationary phases (Figs. 2 and 3). This indicates that the four ST5 DAP-NS strains used in this investigation have no dysregulation in mprF or dltABCD expression associated with their DAP-NS phenotype. Only the ST72 DAP-NS strain, 480, showed significantly enhanced expression of mprF during stationary growth phase (Fig. 2B), correlating with enhanced surface positive charge in cytochrome c binding analyses (Fig. 4), implying that the DAP-NS phonotype in this strain was mediated at least, in part, by charge repulsion mechanism. The strain, 480, exhibited equivalent growth curve kinetics and growth yields over a 24-h period compared with ST5 DAP-S and DAP-NS strains (data not shown), indicating that alteration of biofitness is not likely involved in the DAP-NS phenotype of the strain. Although emergence of DAP-NS MRSA in some DAP-naive patients has been associated with increased cell wall thickness, no substantial change in cell wall thickness or morphology was found among the five DAP-NS MRSA strains (Fig. 5).
In summary, results of this multicenter study suggest that (1) DAP-NS phenotype among MRSA strains isolated from ISA infections can emerge independent of prior exposure to DAP or VAN; (2) Development of DAP-NS phenotype among S. aureus strains from DAP-naive patients is more prominent in MRSA strains compared with that in MSSA strains in Korea; (3) h-VISA phenotype might be associated with increased DAP resistance in some DAP-NS MRSA strains; (4) Increased DAP resistance in the four ST5 DAP-NS strains is independent of alterations in net surface positive charge; and (5) Cell wall thickening is not associated with the observed DAP-NS phenotype in the five MRSA strains. Current studies are in progress to further examine the emergence of DAP-NS phenotype among MRSA strains collected earlier years as well as more recent collection of MRSA strains.
Footnotes
Acknowledgments
This work was presented as an abstract (No. 2011) in the ID Week, New Orleans, LA, October 26–30, 2016. This study was approved by the Institutional Review Board of Seoul National University Bundang Hospital (IRB No. B-1107/132-301). We thank members of the Korea Infectious Diseases (KIND) Study Group and associated staff for their cooperation in this study. Collaborators in the KIND study group were as follows: H.-C.J., K.-H.P., and S.-I.J., Department of Internal Medicine, Chonnam National University Medical School, Gwangju; N.Y. and D.-M.K., Department of Internal Medicine, Chosun University Hospital, Gwangju; Jeong-Hwan Hwang and C.-S.L., Department of Internal Medicine, Chonbuk National University Medical School, Jeonju; J.H.L., Department of Internal Medicine, Wonkwang University Hospital, Iksan; Y.P., Department of Internal Medicine, National Health Insurance Corporation Ilsan Hospital, Goyang; K.S.L., Department of Internal Medicine, Myongji Hospital, Goyang; Y.G.K., Department of Internal Medicine, Inje University Ilsan Paik Hospital, Goyang; S.Y.P., Department of Internal Medicine, and Dongguk University Ilsan Hospital, Goyang, Korea. This work was supported by a grant from Seoul National University Bundang Hospital (grant number 02-2011-017).
Disclosure Statement
No competing financial interests exist.