
Abstract
The predominance of Klebsiella pneumoniae carbapenemase (KPC)-producing K. pneumoniae was caused by the spread of ST258 clone. In Latin America, KPC was reported in 2006, with the isolation of genetically unrelated K. pneumoniae in Colombia. Since then, the expansion of blaKPC in either K. pneumoniae ST258 or other Enterobacteriaceae (ETB) species was increasingly reported. In this study, we characterized 89 KPC-producing Escherichia coli, Klebsiella oxytoca, Serratia marcescens, and Citrobacter freundii that were received between 2010 and 2014. The results revealed that all isolates harbored blaKPC-2. Moreover, the dissemination of KPC by non-K. pneumoniae was mainly caused by the dispersion of ETB mostly genetically unrelated. E. coli is a community pathogen that may serve as the vehicle for the spread of KPC into community settings. Recently, KPC was detected in E. coli ST131, an international epidemic and multidrug-resistant clone. We found that 5/29 KPC-producing E. coli belonged to ST131 and four were blaCTXM-15 producers. The detection of blaKPC in ST131 should be closely monitored to prevent further dissemination. The blaKPC is generally located within Tn4401 transposon capable of mobilization through transposition found in plasmids in ST258. Less is known about the diversity of blaKPC genetic elements that disseminate horizontally among other species of ETB. We found that 16/29 E. coli and 2/18 S. marcescens harbored blaKPC-2 in Tn4401a. In 71 isolates, blaKPC-2 was located amidst diverse Tn3-derived genetic elements bearing non-Tn4401 structure. Further studies on the plasmids that encode blaKPC-2 in these clinical isolates may provide additional insight into its transmission mechanisms.
Introduction
C
The worldwide dissemination of KPC has been caused mainly by the spread of a single successful clone of K. pneumoniae, sequence type 258 (ST258). 3 KPC in Latin America (LA) was first reported in 2006, with the isolation of genetically unrelated K. pneumoniae isolates in Medellin, Colombia. 4 Since then, the expansion of blaKPC harbored by K. pneumoniae ST258 in LA hospitals has been increasingly reported in Brazil,5,6 Argentina,7,8 Uruguay, 9 Puerto Rico,10,11 and Cuba. 12
KPC enzymes have been detected in other ETB, including Salmonella spp., Enterobacter spp., Klebsiella oxytoca, Serratia marcescens, Citrobacter freundii, Escherichia coli, and Proteus mirabilis. 13 Nevertheless, KPC production in these species is less prevalent, but is increasingly reported in our continent, presumably as blaKPC-carrying plasmids are acquired from K. pneumoniae by these species and then disseminated. 14
As mentioned, KPC-producing E. coli are not yet frequent. Nevertheless, the intercontinental dissemination of E. coli ST131 has contributed immensely to the worldwide emergence of fluoroquinolone-resistant and CTX-M-producing E. coli. 15 Several countries in different continents (Europe, America, Asia, etc.) have reported and alerted the detection of KPC in E. coli ST131.16–18 As a consequence, ST131 poses a major threat to public health because it has the potential to cause widespread resistance to carbapenems in a community pathogen. 19 In Argentina, the occurrence of E. coli ST131 was reported in 2012, but this clone was never associated to blaKPC 20 .
blaKPC genes are typically transposon encoded in Tn4401 and blaKPC-bearing non-Tn4401 elements (NTEKPC), named like this to simplify the nomenclature. 21 These elements are carried on conjugative plasmids, which increase its potential for dissemination. 21 The transfer of mobile genetic elements may serve as a vector for interspecies transmission and thus poses a serious challenge for infection control measures. 22
In Argentina, health institutions submit clinical strains for characterization to the National Reference Laboratory in Antimicrobial Resistance (NRLAR). In this study, we describe the molecular characteristics of KPC-producing E. coli, K. oxytoca, S. marcescens, and C. freundii detected between 2010 and 2014, and analyze the diverse genetic environment of blaKPC-2 found in these isolates.
Materials and Methods
Isolates
From June 2010 to March 2014, 89 nonduplicate KPC-producing non-K. pneumoniae ETB were referred to the NRLAR from 9 cities across Argentina (Supplementary Table S1 Supplementary Data are available online at www.liebertpub.com/mdr) due to nonsusceptibility (NS) to ertapenem and positive synergy between a carbapenem and boronic acid disk as follows: E. coli (n = 29), K. oxytoca (n = 22), C. freundii (n = 20), and S. marcescens (n = 18). Isolates were identified using matrix-assisted laser desorption ionization-time of flight mass spectrometry (Bruker, Germany). The isolates were obtained from different anatomical locations (see Results section and Supplementary Table S1), some were rectal screening samples and one from an environmental source.
Susceptibility testing
Minimum inhibitory concentrations (MICs) were obtained by agar dilution to gentamicin, amikacin, ciprofloxacin, minocycline, colistin, fosfomycin, and tigecycline and interpreted according to the Clinical and Laboratory Standards Institute guidelines (CLSI). Imipenem and meropenem MICs were performed by broth microdilution (CLSI). Fosfomycin, colistin, and tigecycline MIC results were interpreted following EUCAST recommendations a .
Molecular characterization
ESBL screening was performed using specific primers listed in Supplementary Table S2 for blaPER-2, and specific for blaCTX-M-2, blaCTX-M-1/15, blaCTX-M-8/25, and blaCTX-M-9/14 groups. Sequencing of amplicons was done on both strands (Applied Biosystems/Perkin Elmer, Foster City, CA). blaKPC was confirmed by polymerase chain reaction (PCR) and sequencing. The genetic surroundings of blaKPC were done as previously described. 8 Sequence analysis was performed using BLAST (www.ncbi.nim.nih.gov). Multiple-sequence alignments were done with Clustall Omega (www.ebi.ac.uk/Tools/msa/clustalo) and BioEdit version 7.0.0. The insertion sequences were searched in the insertion sequence (IS) finder (www-is.biotoul.fr/blast.php).
Pulsed-field gel electrophoresis (PFGE) of Xba-I digested genomic DNA was performed in a Chef-DR® III System (Bio-Rad, Hercules, CA) as previously reported by Gomez et al. 8 Salmonella enterica serotype Braenderup (H9812) restricted with XbaI was used as a molecular weight standard. PFGE gels were analyzed and interpreted according to Tenover et al. 23 Plasmid content and estimated sizes were determined by S1 endonuclease digestion of genomic DNA followed by PFGE (S1-PFGE). 24
Preliminary screening by PCR targeting O25b gene of E. coli ST131 was done as proposed previously 25 and confirmed with E. coli MLST scheme b .
Results
Epidemiological data
The isolates were received from 46 hospitals (9 cities) around Argentina. The median age was 45 years (range 2 months–94 years old) with or without underlying conditions. Blood stream infections were the most prevalent (21%), followed by urinary tract infections (18%), rectal screening (15%), abdominal drainage (9%), skin and soft tissue (6%), and other sites (38%). One K. oxytoca isolate was an environmental sample obtained from dialysis water. The admission of patients was mostly in the intensive care units (35%) and the rest to different wards. The epidemiological data can be found in Supplementary Table S1.
Identification of blaKPC allele, antimicrobial susceptibility, and ESBL content
All isolates harbored blaKPC-2. E. coli and S. marcescens were resistant to imipenem and meropenem (Table 1). K. oxytoca showed 95% non-susceptibility (intermediate plus resistant phenotype) while C. freundii showed 95% and 90% non-susceptibility against imipenem and meropenem, respectively. The evaluation of aminoglycosides rendered 32% of the isolates nonsusceptible to amikacin, while 57% were nonsusceptible to gentamicin. Non-susceptibility to ciprofloxacin was observed in 69% of the isolates. Minocycline showed 38% non-susceptibility, while fosfomycin and colistin were the most active drugs: 2% and 3% non-susceptibility, respectively. S. marcescens is naturally resistant to colistin; therefore it was excluded in this analysis. In addition, we screened for locally prevalent ESBLs (Supplementary Table S1): blaPER and blaCTX-M. 26 blaPER-2 was detected in 12% of the isolates distributed as follows: 32% K. oxytoca, 15% C. freundii, and 5% S. marcescens, whereas blaCTX-M was detected in 19% of the isolates and mostly produced by E. coli 27%. The CTX-M groups detected were blaCTX-M-2 (n = 4), blaCTX-M-1/15 (n = 6), blaCTX-M-9/14 (n = 5), and blaCTX-M-8/25 (n = 2). CTX-M-9/14 was mostly detected in S. marcescens, while CTX-M-1/15 was detected in E. coli.
MICs and range are expressed in μg/ml.
%NS, percentage of nonsusceptibility (intermediate plus resistant); MICs, minimum inhibitory concentrations; nd, not determined.
Genetic relatedness
PFGE analysis showed the absence of major clones; only some related patterns were observed in groups of two to four isolates per species (Supplementary Table S1). In some cases, these related isolates were recovered in a short period of time within the same institution, for example S. marcescens clones A, B, and C and C. freundii clone D. In other cases, related band patterns were seen in isolates recovered >6 months apart in different institutions without evident epidemiological link. Examples are clones G and D of K. oxytoca, or clone E of C. freundii.
In this study, five isolates were confirmed to belong to ST131 (Supplementary Table S1). These isolates had no epidemiological link between them (different years, cities, and institutions), and four PFGE patterns were observed. The two isolates with similar PFGE band pattern “J” were isolated in 2011 and 2014, respectively, in different institutions. All E. coli ST131 were resistant to fluoroquinolones and four out of five harbored blaCTX-M-15.
The number of isolates increased progressively from 2010 to 2013, as well as the number of new hospitals referring isolates (Supplementary Fig. S2). This increment was primarily due to new strains because the genetically related isolates were not detected again for over 1 year either in the same hospital or the country.
Plasmids and genetic platform characterization
All species harbored between one and six plasmids ranging from <48 to ∼360 kb, with an average of two plasmids in C. freundii and K. oxytoca, and three plasmids in E. coli and S. marcescens (Supplementary Table S1). The genetic analysis of the immediate environment of blaKPC-2 revealed that 16 of 29 E. coli (55%) and 2 of 18 S. marcescens (11%) harbored blaKPC-2 in Tn4401a, representing 20% of the isolates studied. In the remaining 71 isolates, we detected a common backbone where blaKPC-2 was flanked by ISKpn8 (also known as ISKpn27 by IS finder) and ISKpn6-like, with or without truncated blaTEM and deletions of different lengths (Fig. 1).
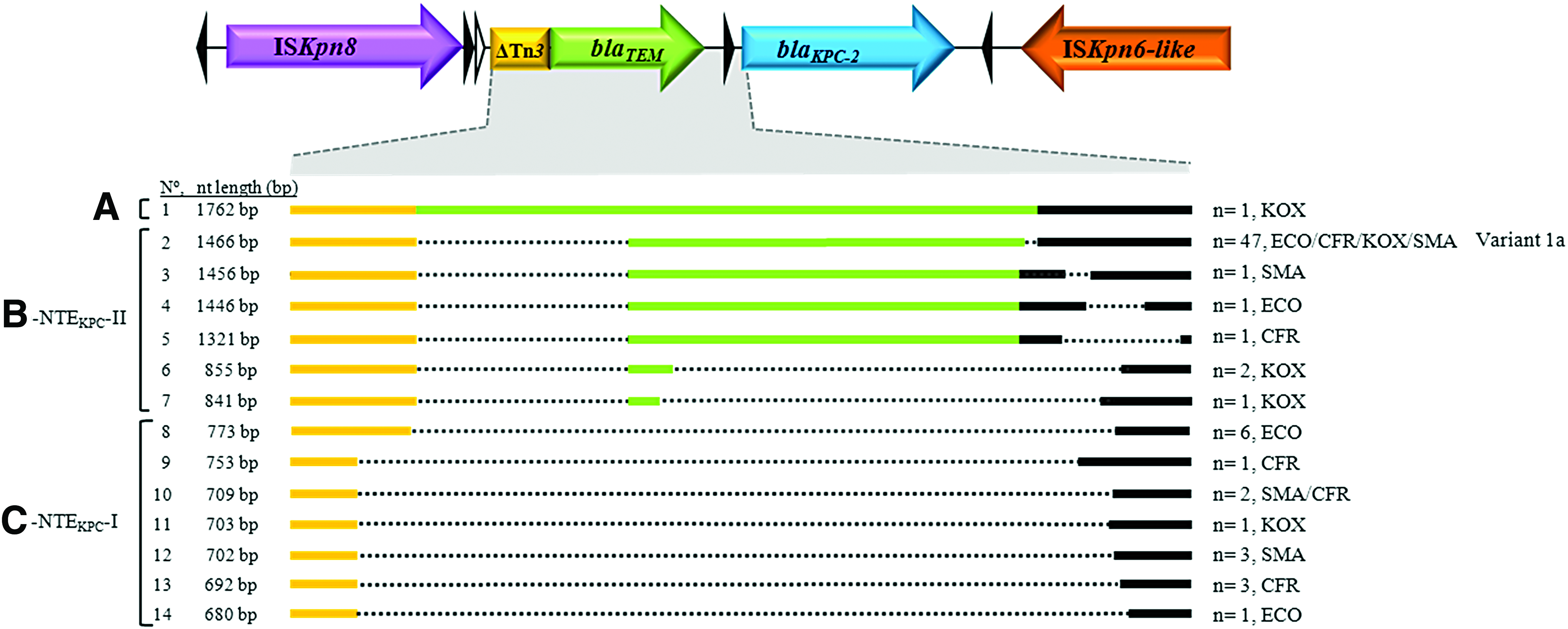
Schematic representation of blaKPC-2 genetic environment in ETB from Argentina. The schematic figure represents the longest sequence obtained and it was used as reference for the alignment shown. The alignment shown is centered on the variations detected upstream of blaKPC-2 (
The sequence alignment of all non-Tn4401 elements (NTEKPC) detected revealed great variability in the length of the region upstream of blaKPC as depicted in Fig. 1. To analyze the genetic structure of these NTEKPC, we separated them in three groups according to the absence or presence of blaTEM. The first is a one-member group composed of a K. oxytoca isolate with a complete blaTEM-1 and gene arrangement: ISKpn8-ΔTn3- blaTEM-1-blaKPC-2-ISKpn6-like (Fig. 1, A, No. 1). The second group includes all the variants detected that have a truncated blaTEM with the following gene array: ISKpn8-ΔblaTEM-blaKPC-2-ISKpn6-like. The major representative of this group was a genetic structure previously called Variant 1a, detected in 47 isolates (66%) (Fig. 1, B, No. 2). This structure seems to be the most disseminated NTEKPC in Argentina.8,27 Interestingly, we also detected different ΔblaTEM truncation lengths that harbor an amino acid mutation, A217T, (according to Lahey nomenclature) not described at that position in any of the TEM enzymes known (www.lahey.org/studies/temtable.asp, last accessed 07/04/2017) (Fig. 1, B, No. 2–7). Finally, a third genetic structure without ΔblaTEM was detected with the following gene order: ISKpn8- ΔTn3- blaKPC-2-ISKpn6-like (Fig. 1, C, No. 8–14). This arrangement was found in 17 isolates (19%) and we observed 7 different variable regions defined between ISKpn8 and blaKPC-2. Moreover, this structure is similar to the sequence originally reported by Shen et al. (GenBank FJ628167.1). 27
In silico analysis of the blaKPC promoter region sequence in NTEKPC isolates revealed the presence of promoter P1 already described in KPC located in Tn4401. 22 The exception was one C. freundii where the P1 promoter was absent, only showing the ribosome binding site and one E. coli without the −35 sequence (Fig. 1, B-NTEKPC-II, line 4). The imipenem MIC of both isolates was 4 μg/mL.
Discussion
Infections caused by multidrug-resistant bacteria constitute the leading cause of serious healthcare-associated infections and are responsible for extended periods of hospital stay, severe illness, mortality, and increased economic burden. 28 Moreover, the attributed mortality to infections due to KPC-producing strains is very high (50% or more). 28 We have reported the emergence of blaKPC in Enterobacteriaceae in Argentina in 2006. Since 2008, we have used an algorithm to implement active surveillance of carbapenemases among laboratories that participate in the National Quality Control Program in Bacteriology (Argentinean Ministry of Health). 8 Briefly, isolates showing the following: (1) imipenem inhibition zones ≤21 mm, (2) NS to expanded-spectrum cephalosporins, and (3) synergy between carbapenems and 3-aminophenyl-boronic acid disks were suspected of KPC production and sent to the NRLAR. In this work, we presented a description of 89 non-K. pneumoniae KPC-2 producers isolated in Argentinean hospitals.
blaKPC-2 is the only allele ever detected in Argentina since the first finding. 7 In LA, blaKPC-3 has been reported in Colombia and blaKPC-2 in southern countries. 29 Nevertheless, this geographical difference is not exclusive: Puerto Rico has reported the novel variant blaKPC-8 and blaKPC-2, and Cuba has reported the emergence of blaKPC-2.10,12
In our study, no major clones were detected, demonstrating that the spread of KPC-2 in non-K. pneumoniae isolates in Argentina is not driven by clonal dissemination, unlike K. pneumoniae ST258. 8 Previous reports, although infrequent, have documented the emergence of KPC enzymes in other ETB different from K. pneumoniae, including Salmonella enterica, K. oxytoca, Enterobacter species, E. coli, C. freundii, and S. marcescens.30,31 In a report from Brazil, two distinct PFGE pattern groups were observed among six E. cloacae and three S. marcescens blaKPC-2 producers. 32 Other reports are from China, where the pattern of KPC dissemination seems to be similar to that of Argentina: many ETB species disseminating in a nonclonal manner, mostly due to horizontal transfer of blaKPC-2-harboring plasmids.
Carbapenem resistance in E. coli in extraintestinal infections is often life-threatening and one of the most important nosocomial pathogens that may serve as a vehicle to spread KPC-mediated resistance into community settings. 14 Nevertheless, data on the molecular epidemiology of KPC-producing E. coli remain relatively scarce. 3 In this study, we show that 5/29 E. coli isolates belong to the international, hyperepidemic, and multidrug-resistant E. coli ST131 clone. This clone has been characterized as highly virulent and has been reported all over the world, representing a major public health problem. To the best of our knowledge, this is the first report of KPC in ST131 in Argentina.
Many resistance genes are located on plasmids that can be transferred between different bacterial strains or species, thus facilitating horizontal gene transfer (HGT). 33 In our study, the plasmids detected vary in number, size, and replicon type among the different strains. This was observed in previous reports, where we and others demonstrated the in vivo and in vitro transference of blaKPC-harboring plasmids between different ETB with diverse genetic backgrounds.8,30,34 Some authors consider that the role of HGT in short-term evolution is still unclear, as capturing the processes in real time is challenging and outbreaks in healthcare settings are usually dominated by clonal transmission. 35 Furthermore, resistance gene mobility can be enhanced by integration into transposable elements, which can autonomously mobilize between different genomic locations. 36 The characterization of the immediate genetic environment of blaKPC-2 in our isolates revealed that Tn4401a was detected in 55% of E. coli isolates referred to the NRLAR. The reasons why Tn4401a moved into E. coli and not to the other ETB species is a question that needs to be answered. The emergence of E. coli producing KPC carried by Tn4401 has been reported in other regions such as USA, 37 Israel, 38 and Greece. 39 In these countries, where the prevalence of KPC-producing ST258-K. pneumoniae is high, the dissemination of KPC-producing E. coli has been mostly nonclonal, as observed herein. Interestingly, results from our study reveal that all isolates harboring Tn4401a came from hospitals located in the metropolitan area of Buenos Aires, while non-Tn4401a isolates were recovered throughout the country. When analyzing blaKPC genetic platforms in reports from LA (Brazil, Colombia, Venezuela, Trinidad and Tobago, Perú, Ecuador, Chile, Paraguay, and Uruguay), most countries report that both blaKPC-2 and blaKPC-3 almost exclusively harbored on Tn4401a or b variants and in diverse ETB species, although K. pneumoniae is the majoritarian species reported, as expected.5,40–43 As an exception, a report from Chile describes the finding of 11 out of 17 blaKPC-2 K. pneumoniae isolates harbored in Variant 1a, and in the rest of the isolates in Tn4401a. They also find 1 blaKPC-24 and 16 blaKPC-2. 43 Reports from the rest of the American Continent suggest the tendency is very similar to LA where Tn4401 is the majoritarian genetic platform detected.11,44–46 The curious finding was that of Puerto Rico where they detected blaKPC on a novel truncated version of Tn4401e located in the chromosome of an Acinetobacter baumanii. 41 In our study, we are reporting a different situation where blaKPC-2 is mostly harbored (80%) in NTEKPC.
In 2009, a novel genetic background of the blaKPC-2 gene was described in China 27 on the plasmid pKP048. blaKPC-2 flanking regions contained a Tn3-based transposon and a partial Tn4401 segment. The genetic environment of the blaKPC-2 gene of 71 isolates analyzed in our study was similar to the genetic structure described by Shen et al. Overall, 66% (47/71) of the NTEKPC isolates harbored the same region around blaKPC-2 (Fig. 1, b-NTEKPC-II, line 2). In addition, other variants were identified in this study (Fig. 1A–C). A similar scenario was reported recently in China where the spread of blaKPC-2 gene between different Enterobacteriaceae species seems to be mediated by HGT of Tn3-based transposons and not by blaKPC-2-carrying plasmids. 30 Further studies are required to determine whether these NTEKPC are a consequence of transposon rearrangements together with HGT of blaKPC-2-harboring plasmids or a different disseminating mechanism of NTEKPC between species.
Conclusions
In this work, we show that the dissemination of non-K. pneumoniae species producing blaKPC-2 in Argentina is not vertical and blaKPC is located on diverse variants of Tn3-derived NTEKPC genetic platforms. We believe that this is the first report of KPC-producing E. coli ST131 in the region. KPC producing gram-negative pathogens, together with other carbapenemases, are significantly changing the global behavior of resistance, similar to other infections that have altered the course of humanity. The reasons are the increasing incidence of multidrug-resistant bacteria, together with the lack of effective antimicrobial stewardship, infection control policies, design of new antimicrobials- and safe approaches to protect the efficacy of the current last-resort drugs. As a consequence, there is compelling need to understand the dissemination mechanisms of these pathogens. Undoubtedly, understanding the genetic determinants that contribute to the dispersion of these bacteria as well as their associated plasmids will lead to improved tracking of resistance and the better control of its spread.
Footnotes
Acknowledgments
Part of this work was previously presented at the 53rd ICAAC meeting in Washington DC as a poster (C-074). D.F. and S.A.G. are members of the Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) research career, and D.D.B. has a doctoral fellowship from CONICET. We are indebted to Roberto Melano, PhD, Shannon Hayes, PhD, for the critical reading of the article. Institutions and professionals who referred the clinical isolates for characterization to NRLAR: Clínica De Los Virreyes, Dra. María Silvia Montone: Clínica Modelo Morón, Leticia Bardi; Clínica Privada Vélez Sarsfield, Lidia Wolf; Clínica Y Maternidad Suizo Argentina, Dra. Silvia Vicario/Dra. Adriana Pecio; Fleni, Dra. Nora Orellana, H. Area Cipolletti, Dra. Roncallo Mariela/Pinoche Laura; H. “Dr Julio Perrando,” Dra. Isabel Marques; H. 4 De Junio–Dr. Ramon Carrillo (Chaco), Dra. Norma Lodeiro/Norma Esther Cech; H. Alvarez, Dra. Mariela Schijman; H. Central De Mendoza, Dra. Selva Ruffolo; H. Cesar Milstein, Nancy Zugasti–Jorge Herrera; H. Cetrangolo (Vte. Lopez), Dra. Silvia Guaycochea; H. De Niños Dr. Ricardo Gutierrez, Dra. Adriana Procopio/Miryam Vazquez; H. Del Niño Jesus (Tucumán), Dra. Ana Villagra De Trejo; H. Evita De Lanus, Dra. Ana Togneri; H. Gral De Agudos “Dr I Pirovano,” Dra. Claudia Garbasz; H. Gral De Agudos “J A Fernández,” Dra. Sara Kaufman; H. Gral.De Agudos Dr. Cosme Argerich, Dra. Marcela Badia; H. Guillermo Rawson (Córdoba), Dra. Ana Littvik; H. Heller, Dra. Gladys González; H. Infantil Municipal De Córdoba, Dra. Liliana González; H. Italiano (Caba), Dra Sandra Mariela Mangiaterra/Dr. Edgardo Moretti; H. Juan Pablo Ii (Corrientes), Bioq. Sandra Pierlorenzi; H. Misericordia (Córdoba), Dra. Carmen Nobile; H. Municipal De Urgencia (Cdba), Dra. Elena D’ Andrea; H. Nacional Prof. Dr. Alejandro Posadas, Dra. Adriana Di Bella; H. Pablo Soria, Dra. María S. Royo De Weibel; H. Provincial Neuquén “Dr. Castro Rendón,” Elizabeth Kummerlen; H. San Roque (Córdoba), Dra. Gladis Pino De Muñoz; H. Vélez Sarsfield, Dra. Manganello Silvana; H. Zonal Bariloche, Dr. Néstor Blazquez; Higa “Dr. José Penna” (Bahia Blanca), Dra. María Luz Benvenutti; Higa San Martin De La Plata, Dra Rosana Padlog Mais; Hospital Britanico, Dra Marta Beatriz Giovanakis; Htal. Arturo Oñativia, Dra. Mirta Bergami; HZGA. Dr. Carlos Bocalandro, Dr. Miguel Andrade; ICCYC, Inst. De Cardiología y Cirugía Card. Fund. Favaloro, Dra. Analía Fernández; Instituto Alexander Fleming, Dra. Sandra Valle; Lab. Priv. Tucumán Norma Cudmani, Lab. Stamboulián, Adriana Súcari; Policlínico Bancario, Dra. Graciela Miranda; Policlínico Central UOM, Pablo Saul; Sanatorio Anchorena, Dra. Nancy Giudice; Sanatorio Mitre, Dra. Lorena Bottinelli; SASJ, Dra. Rochman Zulema B.
Funding
This work was supported by Préstamo BID-PICT-2012-0145 to S.A.G. from ANPCYT and by the regular federal budget of the National Ministry of Health of Argentina.
Authors Contribution
D.D.B. performed most experimental work. C.L., M.R., D.F., and E.A., contributed with experimental work. All authors contributed with the design, analysis, and revision, gave final approval, and agreed to be accountable for all aspects of the work of the finished article. A.C. and S.A.G. did major design of the work, result interpretation, and writing of the article.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.