
Abstract
Streptococcus agalactiae is a causal agent of bovine mastitis and is treated by β-lactam antibiotics (BLAs). Compared to penicillin-resistant S. agalactiae from humans, resistant strains in bovine are rarely reported. In this study, we aimed to investigate BLA resistance and mutations in penicillin-binding proteins (PBPs) of S. agalactiae in central and northeast China. The minimum inhibitory concentrations (MICs) of 129 penicillin-resistant S. agalactiae isolates from cows with mastitis were determined, and the related PBP genes were detected and sequenced. All strains were unsusceptible to penicillin G and mostly resistant to ampicillin, cefalexin, and ceftiofur sodium. One hundred twenty-nine strains were divided into 4 clonal groups and 8 sequence types by multilocus sequence typing analysis. We found a set of new substitutions in PBP1B, PBP2B, and PBP2X from most strains isolated from three provinces. The strains with high PBP mutations showed a broader unsusceptible spectrum and higher MICs than those with few or single mutation. Our research indicates unpredicted mutations in the PBP genes of S. agalactiae isolated from cows with mastitis treated by BLAs. This screening is the first of S. agalactiae from cattle.
Background
S
Currently β-lactam antibiotics (BLAs), such as penicillin and cephalosporins, are extensively used in treatments of Streptococcus spp. However, Streptococcus pathogens with penicillin resistance are continually found worldwide,8,9 particularly in Japan and North America.10–15 This phenomenon is believed to be due to increasing clinical application of BLAs, 16 because the rate of penicillin-resistant strains in clinical samples was higher in regions where new generation BLAs were used for treatment. 17 As the target enzymes for BLAs, penicillin-binding proteins (PBPs) have been the theme of mass research regarding the penicillin resistance of S. agalactiae, For instance, penicillin-binding protein 2X (PBP2X) is highly mutational in clinical resistant isolates, and in particular two amino acid substitutions (V405A and/or Q557E) in PBP2X proved to be the leading cause of the penicillin-insusceptible phenotype.14,15,18 Moreover, various amino acids in PBP2X, PBP2B, and PBP1A were detected among penicillin-resistant S. agalactiae isolates.14,15,19 However, while all mutations of PBP2X, PBP2B, and PBP1A genes were found in S. agalactiae isolated from human host, only limited data are available on bovine S. agalactiae strains. 20
A high degree of distinctiveness was confirmed between the human and bovine S. agalactiae by ribotyping, PCR, DNA sequencing, multilocus sequence typing (MLST),21–23 and by taking into account the different environmental pressures on heterologous host-derived S. agalactiae.24,25 Furthermore, genetic diversity was demonstrated between S. agalactiae isolated from bovine in different Brazilian states. 26 Because of these factors, investigating penicillin resistance patterns of S. agalactiae isolates, specifically from bovines, was required. This research focused on unraveling the correlations between the specific mutations of PBP genes and penicillin insensitivity in S. agalactiae isolated from dairy cows with clinical mastitis.
Materials and Methods
S. agalactiae isolates
A total of 129 S. agalactiae isolates from 16 farms were collected from milk samples of cows with clinical mastitis. The cows were distributed in three provinces located in central and northeast China: 88 isolates were from 13 herds located in Henan Province (HN-1, HN-2, HN-3…HN-12, HN-13), 26 isolates were from 2 herds located in Heilongjiang Province (HLJ-1, HLJ-2), and 15 isolates were from 1 herd located in Liaoning Province (LN-1). All isolates were collected during 2014 and 2016, and confirmed as S. agalactiae via PCR by detecting the dltS gene. 27
Multilocus sequence typing
All 129 S. agalactiae isolates were typed using MLST by sequencing seven housekeeping genes (adh, pheS, atr, glnA, sdhA, glcK, and tkt), and fragments were amplified by PCR. The primers are available at the S. agalactiae MLST website (http://pubmlst.org/sagalactiae). For each isolate, the allele number and sequence types (STs) were defined by analysis of the allele sequence in the MLST database (http://pubmlst.org/sagalactiae). The eBURST V3 program (http://eburst.mlst.net) was used to determine clonal complexes (CCs) among these S. agalactiae strains. 28
Antibiotic susceptibility
The minimum inhibitory concentrations (MICs) were determined by a previously described broth dilution method and were interpreted by following Clinical and Laboratory Standards Institute (CLSI) guidelines. 29 As ceftiofur sodium is not listed in CLSI 2016, the same MIC interpretive standards for other broad-spectrum third generation cephalosporins (cefotaxime, ceftizoxime, and ceftriaxone) in CLSI 2016 were adopted. The MIC of each bovine S. agalactiae strain was tested five times to ensure reproducibility, and Streptococcus pneumoniae strain (ATCC 49619) was used as quality control. All drugs were purchased from the Bio-KONT company in China.
According to the recommendations of the CLSI, because of the rare occurrence of resistant strains of S. agalactiae, antimicrobial susceptibility of all strains were confirmed with the disk diffusion test referred to in the CLSI 29 by another laboratory in the Liao Ning Center for Animal Disease Control and Prevention.
Identification of PBP genes in bovine S. agalactiae
Bacterial genomic DNA was extracted using the EasyPure Bacteria Genomic DNA Kit (TRANSGEN BIOTECH, China). The presence of selected penicillin resistance genes was detected using previously described PCR conditions and primers for encoding putative PBP1A, PBP1B, PBP2A, PBP2B, and PBP2X of S. agalactiae. 15 For identification of the existence of PBP1A, two other sets of primers were used to amplify the fragments in the PBP1A region.14,18 The amplicons were recovered from the agarose using a gel extraction kit (Sangon, China). Then the purified products were directly sequenced in both directions using an ABI automated A373 sequencer (ABI). Sequences were further compared with existing sequences in databases using the BLAST search.
The representative sequence data of mutated PBP genes have been deposited in GenBank under accession numbers KX374346–KX374370. The EditSeq (DNASTAR) was used to profile the deduced amino acid sequences of PBPs.
Results
Isolates and BLA resistance
Four BLAs (penicillin, ampicillin, cefalexin, and ceftiofur sodium) that are officially approved and widely used in the treatment of mastitis were tested. MIC results of 129 S. agalactiae isolates from individual cows originating from 16 different farms in China were investigated in this study. According to the recommendations of the CLSI in 2016, 29 S. agalactiae was defined to be unsusceptible to penicillin when MICs were more than 0.12 (μg/ml), and no resistant interpretive criterion had been designated. In this research, all bovine S. agalactiae isolates were considered to be unsusceptible to penicillin G, as shown in Table 1, and no susceptible strain was isolated. The susceptibilities to ampicillin, cefalexin, and ceftiofur sodium were 9.3% (MICs ≤0.25 μg/ml), 14.7% (MICs ≤0.5 μg/ml), and 51.2% (MICs ≤0.5 μg/ml), respectively. The results of the disk diffusion test were consistent with the MIC results from the broth dilution method, which confirmed the BLA resistance of the S. agalactiae isolates.
GBS, group B streptococcus; MIC, minimum inhibitory concentrations.
MLST analysis
Among the 129 S. agalactiae strains, 8 unique STs (ST-61, ST-67, ST-103, ST-226, ST-301, ST-313, ST-314, and ST-568) and 4 CCs (CC64, CC67, CC103, and CC314) were identified. ST-568 (n = 50) and ST-103 (n = 47) were both clustered into CC103 and represented the largest segment of the strains detected in all three provinces. ST-301 (n = 10), ST-67 (n = 7), and ST-313 (n = 4) were classified into the CC67 group and were the second-largest segment of the strains that were only detected in the central province (Henan). ST-314 (n = 5) and ST-226 (n = 1), both grouped into CC314, were detected in the northeastern provinces (Heilongjiang and Jilin). ST-61, grouped into CC64, was detected in only one farm in Henan Province (Fig. 1). On the basis of the ST described, the comparison study of S. agalactiae strains between herds revealed that similar ST types (ST-67, ST-91, ST-103, ST-313, ST-314, and ST-568) were distributed in different herds, while herd-specific types (ST-61 and ST-226) were also observed during (Fig. 1). The S. agalactiae strains similarity analysis of each herd revealed that S. agalactiae strains exhibit either different types (in seven herds) or similar characteristics (in nine herds) within a same herd (Fig. 1).
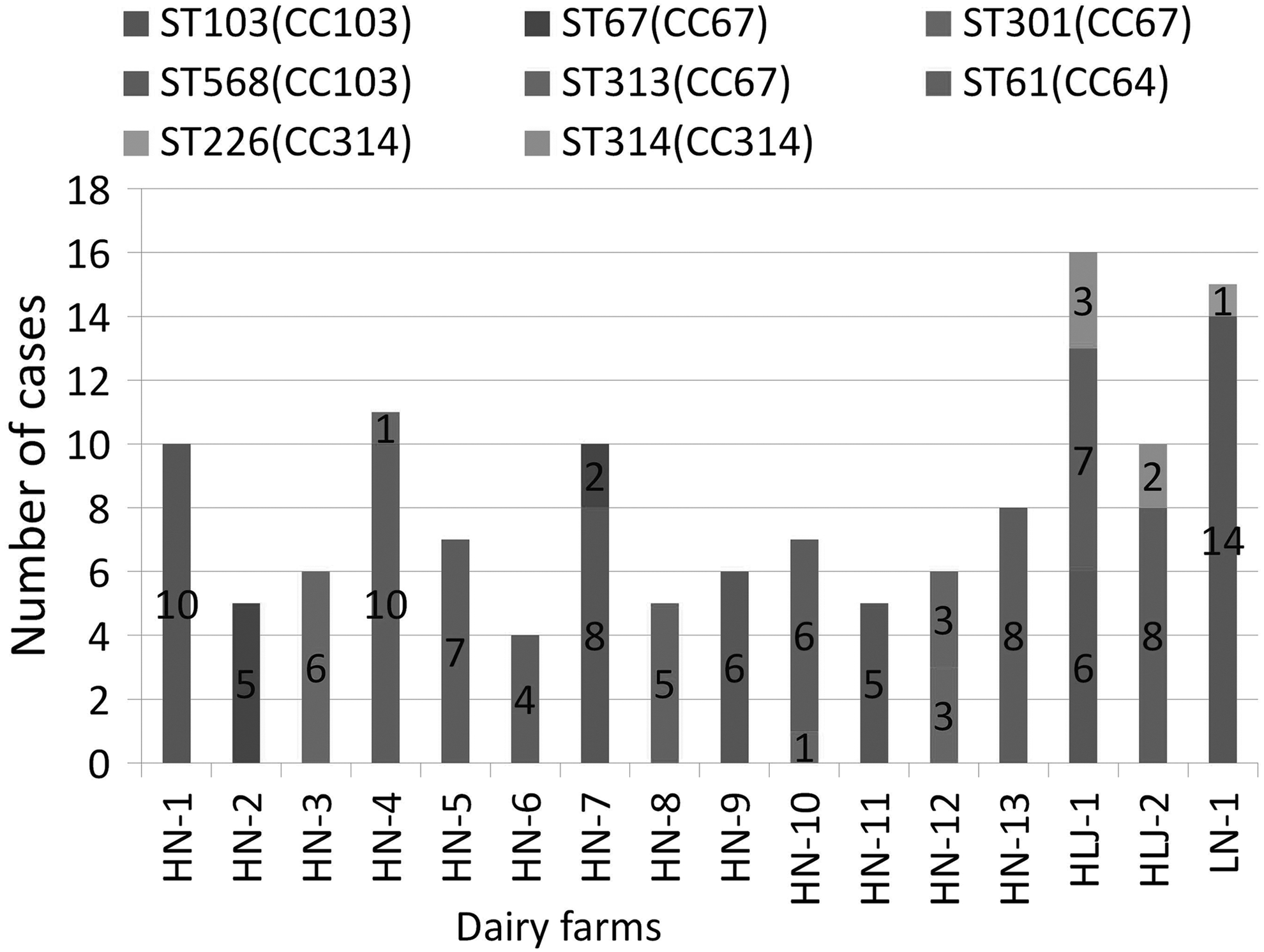
Typing of 129 Streptococcus agalactiae strains present in 16 dairy farms in China. The numbers of S. agalactiae are shown in columns. CC, clonal complexes; MLST, multilocus sequence typing; ST, MLST sequence type.
PBP mutations
Five PBP genes were detected by PCR. PBP1B, PBP2B, and PBP2X genes were identified in the penicillin-unsusceptible isolates, while none of the PBP1A genes was found in any bovine strains. Four nonsynonymous substitutions, L41S, A95D, K651E, and T461I were found in the PBP1B protein in 96 bovine strains, and the A95D substitution was detected in 85 strains. The other three substitutions were found in less than 5 strains. The PBP2A gene was detected in 32 bovine strains, but no nonsynonymous substitutions were found. The PBP2B gene was found in all 129 bovine strains, and the sequencing results revealed eight nonsynonymous substitutions in these strains: V80A, S147A, S160A, Y262N, A399E, G539E, T567I, and G613R. Among these substitutions, the positive rates of the first three substitutions were 66.7%, 38.0%, and 33.3%, respectively, and the rates of the remaining substitutions were under 5.0%. Y336F, I377V, V405A, K425M, and T720S in the PBP2X protein were found in 47, 55, 3, 38, and 19 strains, respectively.
The mutation types of bovine strains
In addition to the single nonsynonymous substitutions in PBP1B, PBP2B, and PBP2X in a few strains, 10 types of multisite nonsynonymous substitutions were found in more than half the strains, and representative isolates of each type are shown in Table 2. These strains were isolated from three provinces hundreds kilometers apart, however, the same multisite nonsynonymous substitutions were detected in all three provinces. In comparison to the single nonsynonymous strains, which showed insensitivity to two or three types of antibiotics and low MICs, all representative isolates of the 10 types were unsusceptible to the 4 BLAs. MICs of strains with two or three substitutions in PBPs were higher than strains with one substitution in PBPs. Most of these strains with multiple substitutions in PBPs were classified in ST-568 and ST-103, and in particular, the NY1512 strain (ST-103) showed the highest MICs to the four BLAs.
PBP, penicillin-binding protein.
Discussion
The high penicillin-resistant rate of bovine S. agalactiae found in our study was similar to previous work in Inner Mongolia. 20 Furthermore, the enterococci strains isolated from mastitis cows in China also show a high frequency of resistance to penicillin. 30 Although penicillin G serves as a primary antimicrobial drug for clinical mastitis in China and was used for decades in veterinary clinics, the high rate of resistance to BLAs in bovine S. agalactiae was still astonishing compared with the low rate of resistance to BLAs in human S. agalactiae.31,32 The results indicate that BLAs should be used discreetly in the treatment of bovine S. agalactiae infection unless a drug sensitive test is detected.
The MLST analysis revealed a distinct heterogeneity among the 129 S. agalactiae strains, which were divided into 8 STs and 4 CCs. All the STs identified during this work have been reported in previous research.26,33–35 ST-568 and ST-103 were predominant among the strains, and such a result is consistent with those reported in eastern China. It is worth noting that a majority of strains with higher MICs were classified in ST-568 and ST-103, which indicates that high-resistance strains have spread to a wider scope than what we measured in the study. Further work was needed to clarify the cause and current epidemic status of resistant strains in the CC103 group on a national scale. Meanwhile, ST-67, ST-301, and ST-313 identified during this work have been reported in Denmark and Brazil.26,35 This is partially due to the previous study focusing merely on herds in eastern China, and the cases in central and northeast China had not been included. 34 In addition, the distribution of STs in China is attributed to national and international animal trade. It sheds lights on why similar STs spring up among several farms in different provinces. In contrast to our results, all strains in several STs (ST-2, ST-23, ST-61, and ST-554) detected in Portugal were found to be sensitive to penicillin, 36 especially the same ST-61 type strains in our research, which were resistant to penicillin. This suggests that another factor, such as clinical drug use, enables change in the biological characteristics of the same ST strains. 12 For instance, PFGE, another typing method, demonstrated that strains grouped in the same ST could be divided into several different PFGE types. 34 Whether such a phenomenon exists in the strains in this work requires further investigation. How different S. agalactiae ST types emerged in same herd still remains unknown. 34 Dogan et al. considered that different sources of infection from humans might be a clue. 22
In the PBP1B protein, the L41S and A95D mutations were also detected in human S. agalactiae strains,14,15 but the high rate of A95D substitution was unexpected, considering that the hot spot regions of human S. agalactiae-related penicillin resistance were mainly in PBP2X, PBP2B, PBP2A, and PBP1A. 37 A95D substitution is a hydrophobic amino acid replacement by a hydrophilic amino acid; therefore, molecular genetic techniques should be used to estimate the role of this amino acid substitution in penicillin resistance of S. agalactiae. Although some substitutions were found in human S. agalactiae strains in PBP2A,14,15 nonsynonymous substitution was not detected. The function of the gene on drug resistance has not yet been reported.
In the PBP2B, V80A, Y262N, A399E, G539E, T567I, and G613R substitutions were also identified in the human S. agalactiae strain, 14 Among these six substitutions, T567I was detected many times in human S. agalactiae strains.14,15 The S147A and S160A substitutions, detected in more than a third of isolated samples, were first identified in penicillin-unsusceptible S. agalactiae strains. The significance of the two emerging amino acid substitutions in penicillin insensitive S. agalactiae requires further assessment in future studies.
In the PBP2X protein, V405 was one key substitution accounting for penicillin nonsusceptibility due to its closeness to the active-site motifs in the transpeptidase domain of PBP2X15,18; however, only three bovine strains with this substitution were detected. Another key substitution, Q557E substitution,14,15,18 was not found in bovine S. agalactiae. I377V was also detected in human S. agalactiae strains and was located in the transpeptidase domain adjacent to active site motifs (344STMK347, 402SSN404, and 552KSG554). 15 Fifty-five strains with the I377V substitution had the highest amounts of penicillin-unsusceptible S. agalactiae isolates with PBP2X in this study. These results warrant further investigation. It is worth noting that the substitution Y336F, near to the motif 344STMK347 and the eight bases close to the active transpeptidase region, was identified in 47 strains. This finding might have an impact on S. agalactiae. K425M was located in the transpeptidase domain, between the 402SSN404 and 552KSG554. That these previous four substitutions were in the transpeptidase domain reveals that the mutation hotspots might be in this region. Moreover, the T720S substitution located in the C-terminal domain was found in 19 penicillin-unsusceptible isolates. The S726L substitution was also reported within the C-terminal domain. 15 In addition, amino acid substitutions in PBP2X were identified as the leading cause for high cephalosporin resistance of S. agalactiae. 11
The same multisite nonsynonymous substitutions were found in various isolates in different areas, which demonstrate that the mechanism of penicillin unsusceptibility in bovine S. agalactiae strains might behave similarly under clinical drug pressure. It was suggested that the multisite nonsynonymous substitutions act synergistically in resistance to these drugs. Nagano et al. found that penicillin MICs of human S. agalactiae strains R7 and R8, each with a unique set of eight amino acid substitutions (two each in PBP2X and PBP2B, three in PBP1A, and one in PBP2A), were higher than other strains with fewer PBP substitutions. 14 These results indicate that amino acid substitutions in different PBPs enhance the penicillin resistance of S. agalactiae. Five new mutations in the PBPs were discovered in the S. agalactiae isolate from a patient with long-term penicillin therapy. 12 This suggested that continual therapy with BLAs used in clinics is a major cause of emerging substitutions in PBPs. According to the proposed classification of S. agalactiae-RBS based on the amino acid substitutions in PBPs, 37 strain SMX1564, strain NY1572, strain SQ1538, and strain SQ1512, which only contain amino acid substitutions in PBP2X, could be classified in Class I, strain NY1512 harbored substitutions in PBP2X, PBP1B, and PBP2B, but not in PBP1A or 2A; therefore, it should be classified in a new proposed Class V. The other four representative isolates with no substitutions in PBP2X, but with substitutions in PBP1B or PBP2B, should be classified in other new Classes under more detailed classification rules.
Conclusions
Emergence of penicillin insensitivity among S. agalactiae isolated from mastitis-afflicted cows appears to be frequent in China. The PBPs data showed remarkable mutations in the PBP1B, PBP2B, and PBP2X regions. Newly discovered amino acid substitutions were found in the majority of samples acquired from the three provinces, and high level of PBP mutations in these strains represents a broad unsusceptible spectrum and abnormal MICs. We suggested that PBP genes of bovine S. agalactiae were affected by the stress of antibiotic treatment for mastitis.
Footnotes
Acknowledgments
The disk diffusion test was performed in Liao Ning Center for Animal Disease Control and Prevention. We thank Shiyu Sun for technical assistance. This work was supported by grants from the Scientific and Technological Project of Henan Province (162102110046) and Nanyang Normal University (CN) (15081).
Disclosure Statement
No competing financial interests exist.