
Abstract
Candida sp. are found as part of the commensal flora in humans but can cause invasive candidiasis in patients with severe underlying disease, especially cancer patients. These patients are frequently subjected to nonsurgical anticancer treatments such as ionizing radiation and anticancer drugs, which kill proliferating human cells by damaging DNA but also affect the microbiota of the patient. C. tropicalis, an emerging fungal pathogen, is associated with high mortality rates of cancer patients especially in tropical regions. In this study, we have investigated the in vitro susceptibility of 38 C. tropicalis clinical isolates from several Mexican hospitals to chronic treatments with several DNA damaging agents, including oxidizing compounds and anticancer drugs. C. tropicalis isolates displayed a high variability in their susceptibility to hydrogen peroxide (H2O2) while showing a high susceptibility to bleomycin (BLM), an anticancer drug that causes double-strand breaks in DNA. This contrasted with the moderate-to-high resistance exhibited by several C. albicans laboratory strains. At least for the C. tropicalis reference strain MYA3404, this susceptibility was hardly modified by the presence of serum. Our results open the possibility of using susceptibility to BLM to differentiate between C. tropicalis and C. albicans; however, analysis of a larger number of isolates is required. The use of BLM for prevention of C. tropicalis infections in neutropenic patients with cancer should be also evaluated. Finally, the variable susceptibility to H2O2 might be due to allelic variation of the histone acetyl-transferase complex which modulates the induction kinetics of H2O2-induced genes in C. tropicalis.
Introduction
T
The relative incidence of the individual NCAC species is variable and largely influenced by both latitude and environmental conditions of the regions where population-based studies were conducted. For instance, C. tropicalis is more prevalent in tropical regions where it ranges second in incidence, only overtaken, and not always, by C. albicans. Importantly, C. tropicalis has been reported as one of the most frequent causes of blood stream and urinary tract infections in the hospital.2–5 C. tropicalis is more common in oncology patients, in particular bone marrow transplantation recipients and those suffering hematological malignancies.6,7 Besides, C. tropicalis disseminates rapidly in the immunocompromised patient causing fungemia with mortality rates (40–70%) as least as high as C. albicans.
Among the conditions that trigger Candida spp. pathogenesis is the immunosuppression linked to nonsurgical cancer treatment, including ionizing radiation (IR) and treatment with drugs, such as bleomycin (BLM) and camptothecin (CPT), which damage human DNA and kill proliferating cells.
IR causes many lesions (mainly through the generation of reactive oxygen species or ROS), including a small number of lethal double-strand breaks (DSBs), oxidized bases, abasic (AP) sites, and single-strand breaks (SSBs) terminated by 3′-phosphoglycolate esters (3′-PG sites/5′P), which are frequently clustered and cause DSBs.8,9 BLM causes a more restricted set of oxidative lesions, mainly 4′-oxidized AP sites and DSBs with 3′-PG ends.10,11 Degradation by BLM is initiated by generating a free radical, in the presence of ferrous ion, in the deoxyribose resulting in two different types of DNA damage. At low oxygen tension, oxidized AP sites are favored, while at high oxygen tension SSBs and DSBs predominate. These alternative pathways lead to a mix of AP sites and strand breaks, occurring at a 1:1 ratio. 12
Importantly, ROS are also naturally produced by immune cells such as macrophages, dendritic cells, or neutrophils, and this substantially potentiates their killing abilities on invading pathogens, including C. albicans and other fungi. In response to this threat, Candida sp. have acquired defense against ROS. It is known that the inactivation of oxidative stress responses attenuates the virulence of C. albicans and its ability to counteract phagocytic killing. 13 However, there are no data concerning the response of other Candida sp. to hydrogen peroxide (H2O2) or other ROS producing systems.
In addition to their action on human cells, nonsurgical cancer treatments, such as IR and anticancer drugs, affect the microbiota of the patient. Therefore, it is important to know the extent to which commensal species that may thrive in the immunosuppressed patients are affected by each specific anticancer treatment.
Previously, we have reported the effect of several genotoxins on C. albicans. 14 In this work, we have characterized a group of 38 C. tropicalis isolates from different hospitals of the Mexican Republic in terms of susceptibility to DNA damaging agents, including oxidizing agents and anticancer drugs. We found that most, if not all, isolates were resistant to menadione (MND) but displayed a high variability in their susceptibility to H2O2. Importantly, all C. tropicalis isolates showed a high susceptibility to the anticancer drug BLM, which contrasted with the moderate-to-high resistance exhibited by several C. albicans laboratory strains. Since serum has been reported to largely reduce the BLM susceptibility of C. albicans, 15 we also investigated its effect on the BLM susceptibility of C. tropicalis.
Materials and Methods
Materials
BLM (Bleocin™) was from Calbiochem and CPT from MBL (International Corporation). MND and hydroxyurea (HU) were from Sigma-Aldrich (now Merck) and H2O2 from Panreac Chimica (Barcelona). Both types of bovine serum albumin, adult and calf serum iron fortified, were purchased from Sigma-Aldrich (Merck). BLM and MND were dissolved in sterile water and CPT in DMSO.
Strains
C. tropicalis MYA3404 (ATCC) was used as the reference strain. For comparison purposes, we have analyzed in parallel several C. albicans laboratory strains. Forty-one C. tropicalis strains were supplied by InDRE (Instituto de Diagnóstico y Referencia Epidemiológicos) (Mexico) (Table 1). When possible, the origin and characteristic of each isolate are included. Three strains (numbers 18, 25, and 32) could not be revived in our laboratory. All the C. tropicalis and C. albicans strains were maintained at −80°C in YPD supplemented with 16% glycerol and routinely propagated and grown in liquid or solid YPD medium (1% yeast extract, 2% peptone, 2% glucose, plus 2% agar when required). When indicated, YPD plates were supplemented with 5% serum, either bovine serum adult or calf serum iron fortified. Only the second one was able to induce germ tube formation by C. albicans at 37°C.
Ct, C. tropicalis.
Molecular diagnosis of C. tropicalis isolates
We took advantage of a species-specific PCR using oligonucleotides, CTR1 and CTR2, designed from the internal transcribed spacer (ITS) regions, ITS1 and ITS2, of the rDNA gene, as described. 16
Sensitivity to DNA-damaging agents
The effect of chronic exposure (i.e., always in the presence of the drug) to the several DNA-damaging compounds, including H2O2 (4–5 mM), MND (0.1–0.2 mM), HU (30 mM), CPT (20–40 μg/ml), and BLM (2–4 μg/ml), was determined using two assays.
In the first one (drop-test), the isolates were grown for the indicated time (12 and 20 hours for exponential and prestationary phase cells, respectively) in liquid YPD medium at 37°C. Then, they were diluted back to an OD600 = 1. Seven microliters of the initial cell suspension and serial fivefold dilutions were spotted onto YPD plates supplemented or not with the genotoxin. Following incubation at 37°C for 24–48 hours, plates were photographed. For a preliminary estimation of the relative susceptibilities, isolates were sorted according to the following criteria: R, isolates whose growth was not affected at all by the presence of the drug; S, isolates for which the number of dilutions showing growth was lower in the presence of the drug than in its absence; and I, isolates showing growth at all the dilutions but lower in the presence of the drug.
In the second assay, about 250–300 colony forming units (CFUs) from exponentially growing cells were plated on YPD supplemented with either H2O2 (4 mM) or BLM (2 μg/ml). All the plates were incubated in the dark at 37°C for 48 hours. Then, colonies were counted and the percentage of cell survival (CFU with drug/CFU without drug) was calculated. Under these conditions, we considered sensitive (S), intermediate (I), and resistant (R) isolates showing ≤45%, 46–84%, and ≥85% survivors, respectively. Two duplicate experiments were performed and results (means and standard errors) graphed in SigmaPlot.
Results
Molecular diagnosis of C. tropicalis isolates
As shown in Fig. 1, when PCR amplified using oligonucleotides CTR1 and CTR2, all the Mexican isolates as well as the reference strain yielded a single 357-bp amplicon specific of C. tropicalis. 16
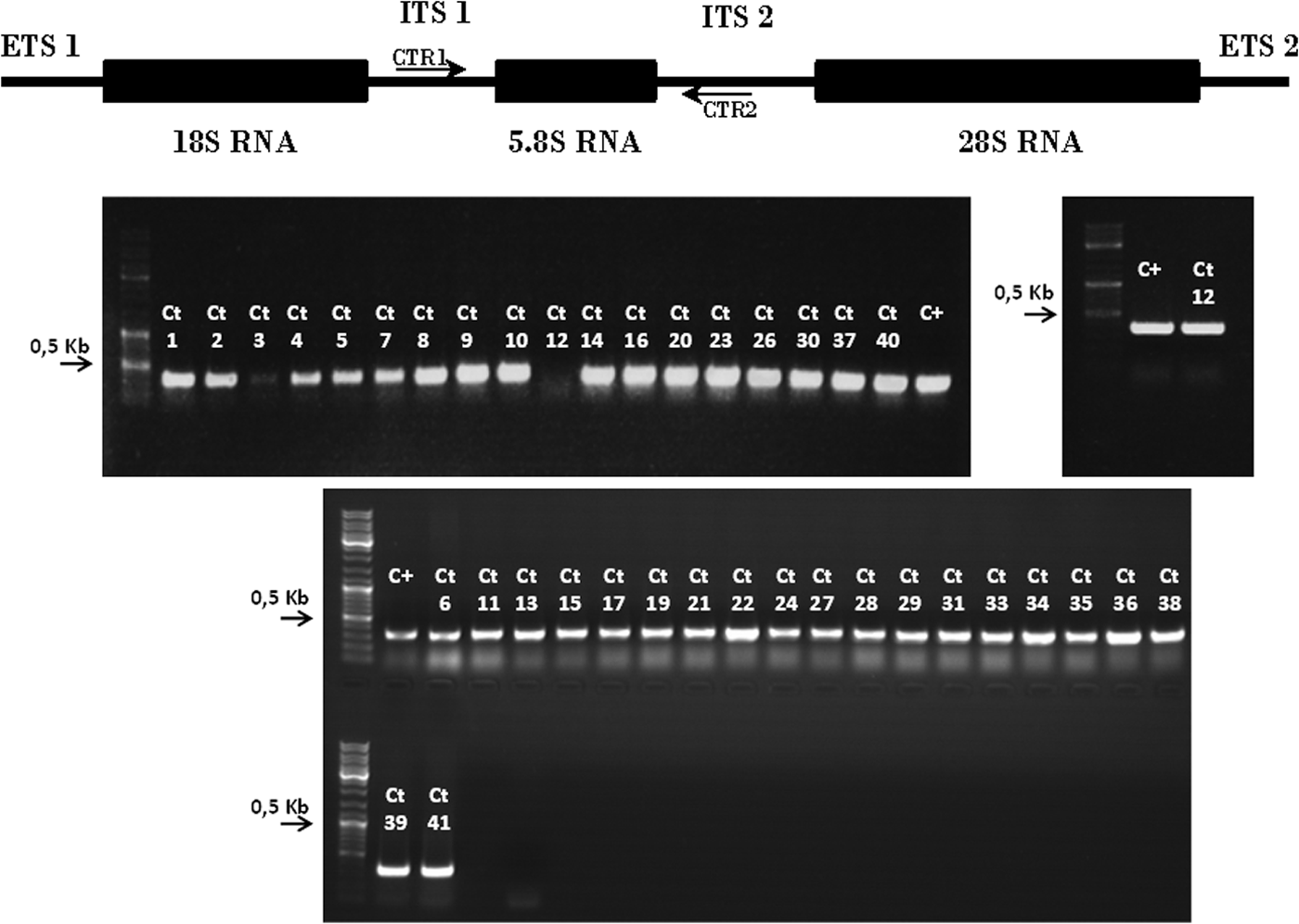
Molecular diagnosis of C. tropicalis isolates. PCR amplification bands from genomic DNA of the indicated C. tropicalis (Ct) isolates using oligonucleotides CTR1 and CTR2. 16 CTR1 and CTR2 are indicated in the diagram. All the clinical isolates as well as the MYA3404 reference strain showed a 357-bp band indicative of C. tropicalis. Molecular weight markers are on the left side of each gel. The arrow indicates the 500-bp marker.
Susceptibility of C. tropicalis to DNA-damaging agents in a drop-test assay
We first used a drop-test to estimate the susceptibility of the C. tropicalis isolates to several agents that impair DNA replication (HU) or cause DNA damage (BLM, H2O2, MND). Preliminary analyses were carried out to calculate the optimal concentration of each genotoxin discriminating between the several isolates.
Based on our previous results with C. albicans reference strain SC5314, 14 we tested the effect of 5 μg/ml BLM, 20 μg/ml and 40 μg/ml CPT, and 0.1 mM and 0.2 mM MND on two randomly selected C. tropicalis isolates (Ct3 and Ct5).
We could not detect growth on plates supplemented with 0.2 mM MND even at the highest cell concentration (3.7 × 106 cells/ml). However, 0.1 mM MND was appropriate to detect differential growth inhibition and the same was true for 5 μg/ml BLM. All C. tropicalis isolates were resistant to CPT at concentrations of 20 μg/ml and 40 μg/ml; therefore, this drug was not considered for further analysis. For HU and H2O2, we selected concentrations of 30 mM and 4–5 mM, respectively, as previously reported14,17 (not shown).
The effect of DNA-damaging agents may vary with the growth stage, since fast-replicating exponentially growing cells (OD ≈ 2–4) could be more affected than slow-replicating or nongrowing prestationary or stationary cells (OD ≈ 8–10). Therefore, we investigated the effect of the growth phase on the susceptibility of six C. tropicalis isolates (Ct1–Ct6) to the abovementioned genotoxins using the drug concentration calculated in the preliminary drop-test assays. For MND, BLM, and HU, we did not detect growth-phase-based differences in susceptibility and only for H2O2, exponentially growing cells were slightly more susceptible than stationary-phase cells (Fig. 2). On the basis of those results, the susceptibility of each isolate to the several genotoxins was determined exclusively with exponentially growing cultures.
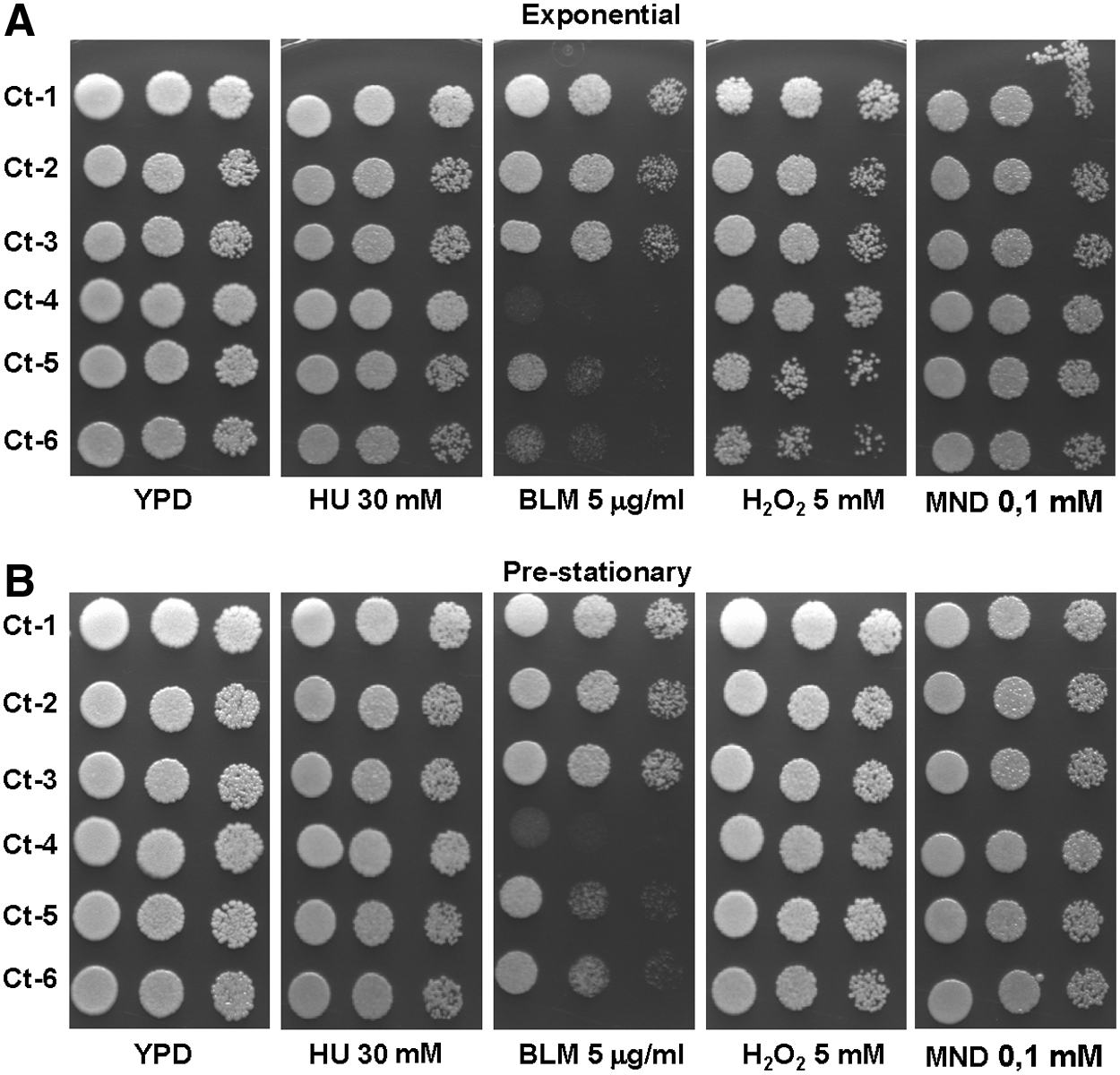
Effect of growth phase, exponentially
A summary of the susceptibility of exponentially growing C. tropicalis isolates to the several genotoxins in the drop-test assay is shown in Table 2 and the actual results yielded by several strains are shown in Fig. 3. We found that most isolates showed a high susceptibility to both BLM and H2O2 and only a few displayed a moderate susceptibility to one or both agents. In addition, most isolates displayed resistance or intermediate sensitivity to MND. Quite surprisingly, all the isolates were resistant to HU, a drug that depletes the ribonucleotide pools and causes cells to arrest at the S phase of the cell cycle.
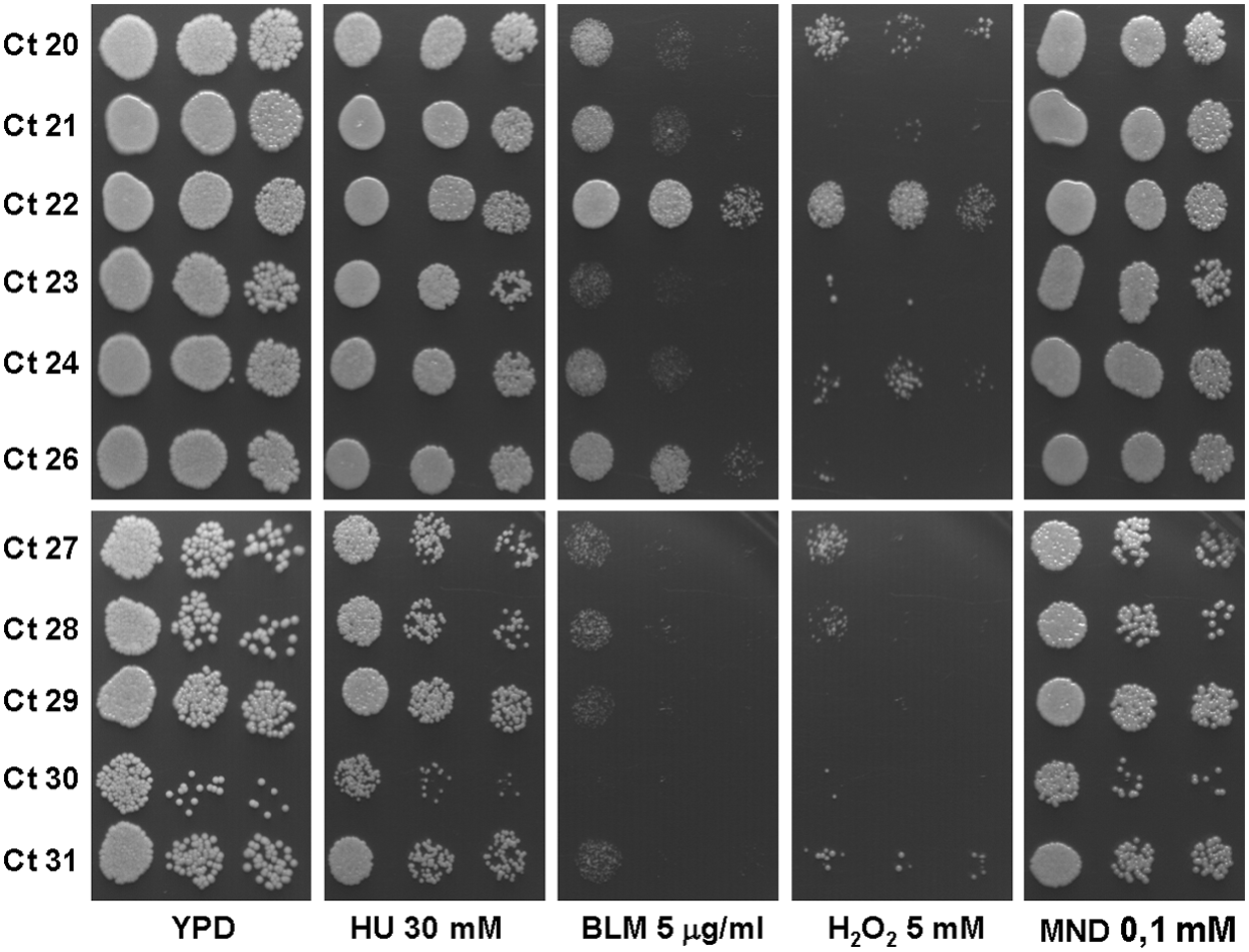
As indicated in the Materials and Methods section, text, S, I, and R, was an arbitrary classification that allows estimation of the relative susceptibilities of the isolates, as follows: R, isolates whose growth was not affected at all by the presence of the drug; S, isolates for which the number of dilutions showing growth was lower in the presence of the drug than in its absence; and I, isolates showing growth at all the dilutions but lower in the presence of the drug.
I, intermediate; R, resistant; S, susceptible.
Quantification of susceptibility of C. tropicalis isolates to H2O2 and BLM
Determination of drug susceptibility using the drop-test does not discriminate between the fungicidal effect and the growth retardation caused by the drug. To quantify the sensitivity of C. tropicalis isolates to BLM and H2O2, we used a quantitative assay based on determination of cell survival (ratio of CFUs in the presence vs. the absence of the drug) (see the Materials and Methods section). Preliminary experiments indicated that concentrations of BLM and H2O2 of 2 μg/ml and 4 mM, respectively, were optimal to discriminate between isolates (not shown).
We found that, quite strikingly, susceptibility to 4 mM H2O2 fluctuated widely among the C. tropicalis isolates; some of them (Ct8, Ct10) were very sensitive (0–1% survivability), whereas others yielded 89–100% survivors (i.e., Ct14, Ct23, Ct24). To evaluate the possibility that the assay was not reproducible enough because of the intrinsic instability of H2O2, we performed a fluctuation test using three independent colonies from each isolate. We found that within each isolate, fluctuation was rather low (Fig. 4A).
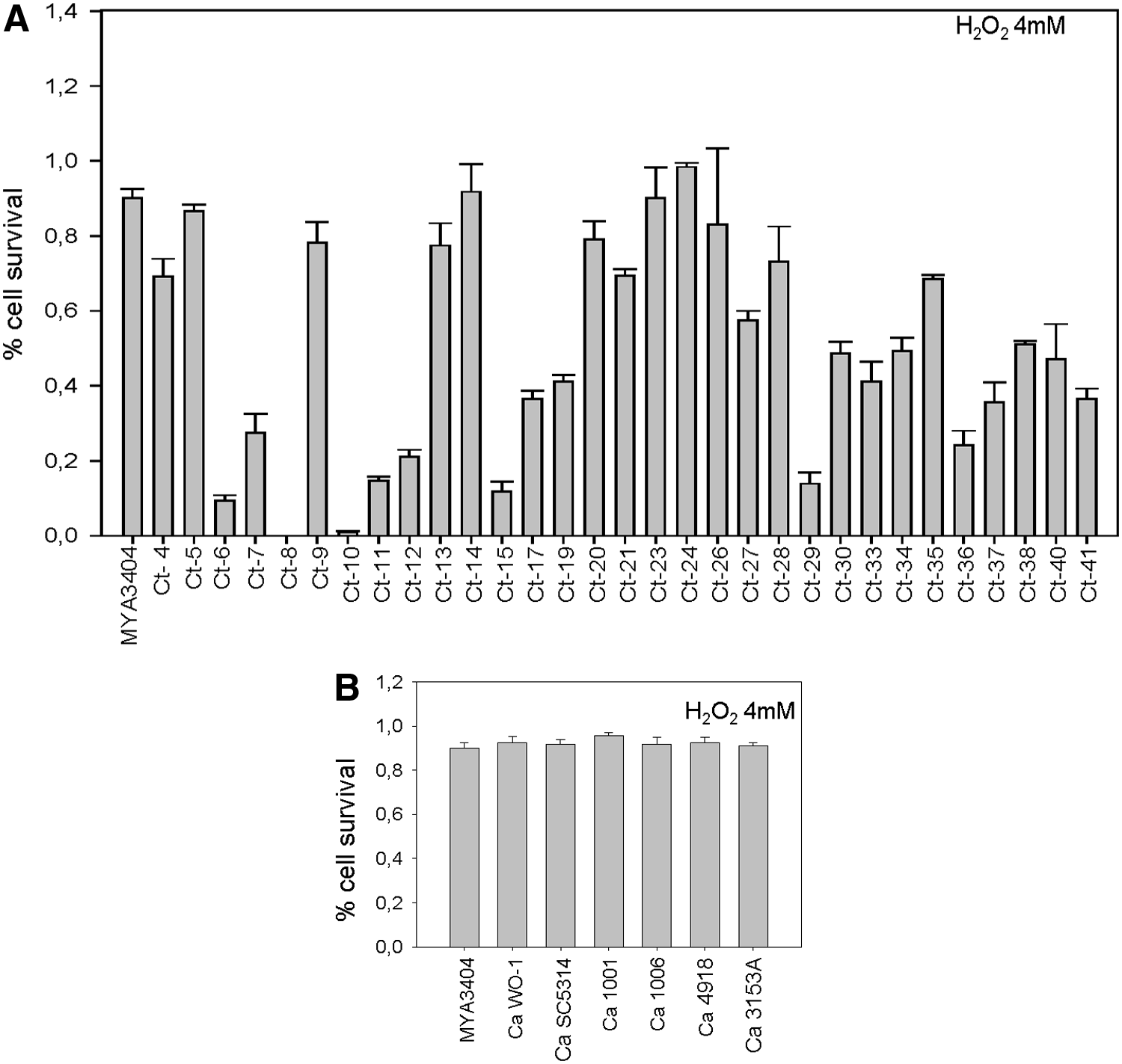
A comparison of susceptibilities of C. tropicalis clinical isolates
According to the criteria defined in the Materials and Methods section, 45% of the isolates were sensitive, 42% intermediate, and 13% resistant. By comparison, reference strains C. tropicalis MYA3404 and C. albicans SC5314 were rather resistant (80–90% survivability), and the same was true for several other C. albicans laboratory strains (WO-1, 1006, 1001, 3153A, and 4918) (Fig. 4B).
Quantitative tests confirmed the high susceptibility of C. tropicalis to BLM (survivability to 2 μg/ml BLM <13%) and further showed that within each isolate fluctuation was also low (Fig. 5A). This result contrasted with the high resistance (75% survivability) reported for C. albicans SC5314, 14 which extends to all its derivatives commonly used as laboratory strains (CAF2-1, CAI4, RM10, RM1000, BWP17, and SN strains) (not shown). Furthermore, other C. albicans isolates of different origins, also used as laboratory strains (Table 1), exhibited resistance to BLM (70–75% for strains WO-1, 3153A, 1006, and 1001, and around 50% for the most sensitive C. albicans isolate—strain 4918) (Fig. 5B).

A comparison of susceptibilities of C. tropicalis clinical isolates
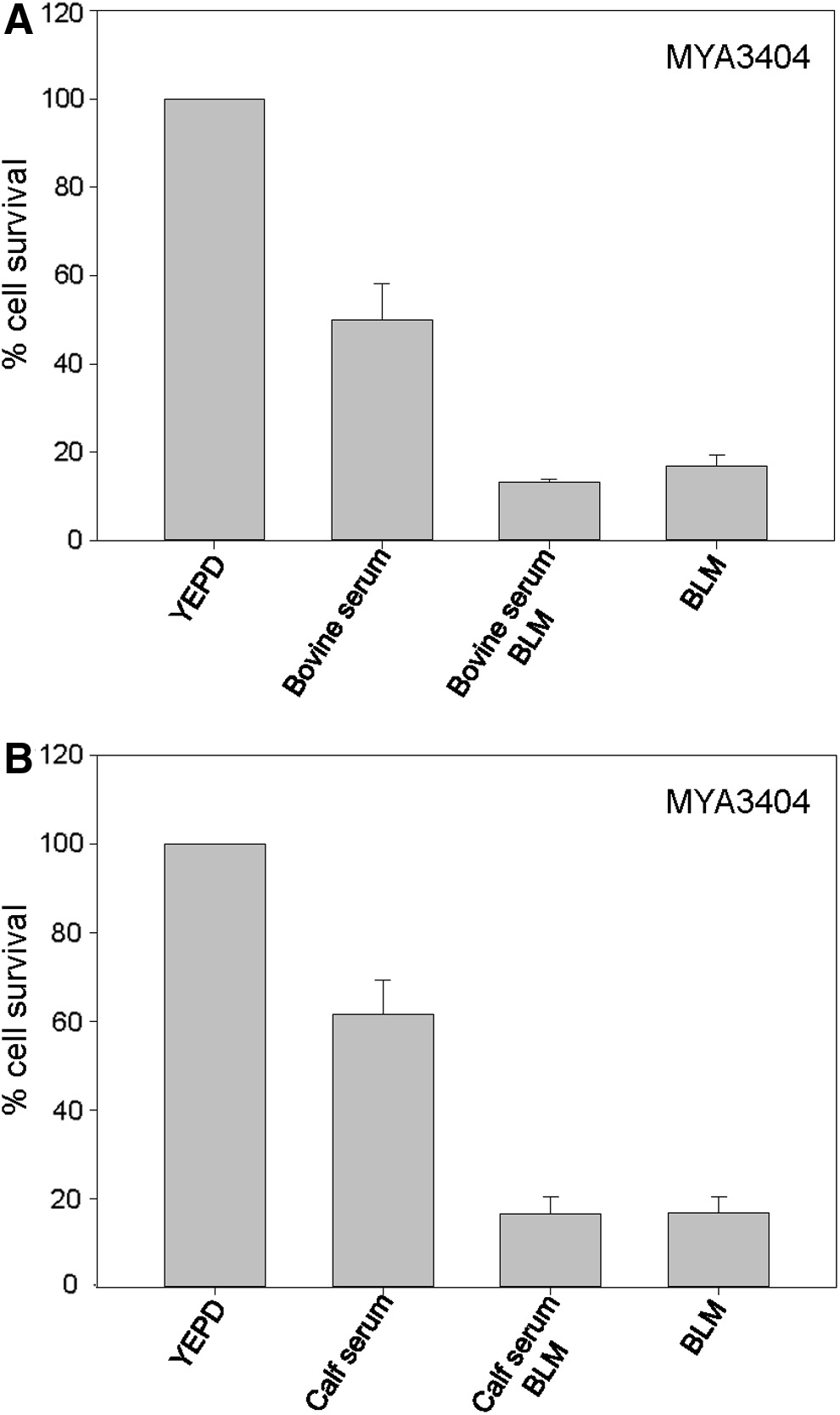
It has been reported that the MIC for BLM of a C. albicans isolate was markedly increased when determined in the presence of serum. 15 To extend this analysis to C. tropicalis, susceptibility of reference strain MYA3404 was analyzed on YPD plates supplemented or not with serum in a quantitative assay. It is well known that C. albicans generates germinative tubes when incubated with serum at 37°C. 18 We found that, under these conditions, only calf serum but not bovine serum induced formation of germinative tubes by C. albicans reference strain SC5314.
Unexpectedly, regardless the kind of serum, its presence lowered the CFU number of C. tropicalis by 40–50% (Fig. 6) (see the Discussion section). In addition, we found that the killing effect of BLM was lowered, but only in a moderate extent, by the presence of bovine serum (76 ± 6% in the presence vs. 87 ± 5% in the absence of serum) (Fig. 6A). This was independent of the ability of serum to induce germinative tubes since similar results were obtained when bovine serum was substituted by calf serum (Fig. 6B).
Discussion
In this study, we have determined the susceptibility of C. tropicalis to two genotoxins, BLM and H2O2. We found that, compared to 6 unrelated isolates of C. albicans, the 38 isolates from C. tropicalis displayed a high susceptibility to BLM. This observation points to the possibility that BLM susceptibility could be a species-specific trait of C. tropicalis. Obviously, a survey with a larger number of C. tropicalis (and other Candida spp.) strains is necessary before the test can be routinely used to differentiate between both species.
In a previous survey, no significant differences between C. albicans and C. tropicalis were found using the National Committee for Clinical Laboratory Standard method. However, in that survey, only one isolate from C. tropicalis and five from C. albicans were analyzed. 15 We have also found that C. albicans strain 4918 was more susceptible to BLM than the rest of C. albicans strains, suggesting that strain 4918 might belong to a different clade. Susceptibility to BLM is important because among the conditions that trigger Candida pathogenesis is the immunosuppression linked to nonsurgical cancer treatment.
BLMs are a family of glycopeptide antibiotics that have potent antitumor activity against a range of lymphomas, head and neck cancers, and germ cell tumors. 19 In this regard, our results predict that C. tropicalis should be infrequent in hematological patients treated with Blenoxane (Bristol-Myers Squibb), the clinically administered form of the drug. The molecular bases of differential BLM susceptibility between C. albicans and C. tropicalis are not known. One possibility is that C. albicans has more powerful DNA repair systems, but differences in BLM uptake (or efflux pump efficiency) or in the ability of sequestering the drug into the vacuole should be also considered. For instance, S. cerevisiae mutants deficient in BLM uptake or the endocytic pathway exhibited alterations in BLM sensitivity. 19
It should be pointed out that, in contrast to a previous report, 15 we found that the susceptibility of C. tropicalis MYA3404 to BLM was only moderately affected by serum. The inhibition caused by serum on the number of C. tropicalis CFUs was striking. One possibility is that serum induces the production of E,E-farnesol, which has been reported to cause growth inhibition on Candida sp., C. tropicalis being most strongly affected. 20
The variability in the susceptibility of C. tropicalis isolates to H2O2 was also an unexpected result. In addition to causing oxidative lesions on DNA, H2O2 may also oxidize thiol linkages of essential proteins further compromising cell viability. A Blast search using each peroxisomal (CTA1) and cytosolic (CTT1) S. cerevisiae catalase as queries identified a unique catalase in C. tropicalis, referred to as peroxisomal, with E values of 1.1.e−180 and 1.1.e−127, respectively. One possibility is that this enzyme is inactivated or repressed in the most susceptible isolates. However, although important, detoxification of H2O2 by catalase is only 1 of the 120 biochemical reactions implied in H2O2 metabolism in C. albicans. 13
Importantly, the histone acetyl-transferase (HAT) complex, which modulates chromatin assembly, is involved in the resistance to oxidative stress of several species from the Candida clade, including C. tropicalis, but not of other Saccharomycetaceae, such as S. cerevisiae or C. glabrata. This HAT complex modulates the induction kinetics of H2O2-induced genes. 21 Therefore, allelic variation of the HAT complex genes in C. tropicalis could be responsible for the observed variability in susceptibility to H2O2. It would be interesting to know whether C. tropicalis isolates from other worldwide geographical regions behave similarly to Mexican isolates. It is expected that C. tropicalis strains with high sensitivity to H2O2 exhibit less virulence in invasive and disseminated infections or are more easily cleared from blood stream where the fungus is exposed to competent neutrophils and macrophages.
Conclusions
This study provides for the first time evidence that C. tropicalis shows a significantly higher susceptibility than C. albicans to BLM. Many more isolates need to be analyzed to assess whether this differential susceptibility can be used as a test to discriminate between both Candida sp. Our results predict that cancer patients treated with BLM should be more refractory to suffer C. tropicalis invasion. In this regard, it would be interesting to analyze the susceptibility of clinically important Candida sp. to BLM or other anticancer drugs. Finally, although susceptibility of C. tropicalis to H2O2 is not a species-specific trait, it could be interesting to analyze the possible correlation between H2O2 susceptibility and virulence of different isolates.
Footnotes
Acknowledgments
The authors thank Belén Hermosa for technical support and Cudberto Contreras from InDRE (Mexico) for supplying the C. tropicalis clinical isolates.
Authors' Contributions
N.G.C., C. tropicalis drug susceptibility; A.B., C. albicans drug susceptibility; R.C., influence of serum; A.E.T., molecular characterization of C. tropicalis isolates; T.C. and G.L. conceived, designed, and supervised experiments and edited the article.
Disclosure Statement
No competing financial interests exist.