
Abstract
Antibiotic resistant bacteria present a growing global healthcare challenge. Previous research demonstrates that wild birds harbor extended spectrum β-lactamase (ESBL)–producing Enterobacteriaceae and may contribute to their dissemination. We aimed to assess prevalence and temporal variation in the detection rate of ESBL-producing bacteria in urban wild birds and to evaluate methods regarding sample handling. Monthly fecal sampling was performed in 2013 at an urban pond in Sweden. ESBL-producing Escherichia coli and Klebsiella pneumoniae were analyzed by polymerase chain reaction targeting blaCTX-M. Subsets of samples were analyzed in multiple replicates and without previous freezing. Pond water samples were screened for 12 antibiotics. Out of 813 fecal samples, 47% grew ESBL-producing E. coli, a higher prevalence than in similar studies. Detection rate varied considerably between months, ranging from 4.2% in May to 84% in July, and was significantly higher during warm months. A majority of isolates harbored CTX-M-15 type ESBL. Detection rates were increased by duplicating samples and by avoiding freezing. No antibiotics were detected in pond water. This study demonstrates high prevalence and a previously undescribed temporal variation in detection rate of ESBL-producing Enterobacteriaceae in wild birds. The distribution of CTX-M genes corresponds well with Swedish human isolates, indicating communication between the genetic pools of ESBLs in humans and wild birds. Urban ponds may serve as important natural reservoirs for antimicrobial resistance.
Introduction
T
In 2005, an outbreak of ESBL-producing K. pneumoniae occurred at Uppsala University Hospital. The outbreak lasted throughout 2007, affecting 248 patients. Analysis revealed all isolates belonging to a single CTX-M-15-harboring clone of K. pneumoniae. 8 Antimicrobial resistance was conferred by the plasmid pUUH239.2, harboring the blaCTX-M-15 gene. In 19 of the 248 cases, the plasmid had been transferred to E. coli within the patient. 9
In the last decade, antibiotic resistant bacteria in natural reservoirs and wildlife have received increasing interest from the scientific community. Wild birds have been proposed as an important reservoir of resistant bacteria and as vectors for the transmission of resistant strains across continents. The potential of birds as vectors for zoonotic disease has been demonstrated in the context of West Nile Virus in the United States. 10 Evidence suggests bidirectional transmission of resistant bacteria between wild birds and humans.11–13 For instance, Franklin's Gulls (Leucophaeus pipixcan) share sequence types of ESBL-producing E. coli with human clinical isolates corresponding to both wintering and breeding grounds of the birds. 14 The detection rate of resistant bacteria in wild birds correlates with proximity to environments influenced by human activity, in particular agricultural and sewage treatment industries.11–13,15 Nevertheless, birds in remote areas with limited human presence have also been found to host antibiotic resistance.16,17
ESBL-producing E. coli has been isolated from a wide range of bird species across the world, and the highest prevalence has been recorded in birds of prey and in aquatic-associated species.11,12 Three studies have detected ESBL-producing E. coli in wild ducks in Europe: 13 out of 67 fecal samples from ducks (19%) were positive in a study from the Netherlands, 3 out of 86 sampled Mallards (3.5%) in a Polish study, and 0 out of 305 Mallards (0.0%) in a study from the Czech Republic.18–20
Regarding ESBL-producing bacteria in wild birds in Sweden, one study performed in 2008—at the same site as this study—found ESBL-producing E. coli in 0 out of 17 (0.0%) sampled Mallards 21 ; a study on gulls from nine European countries found ESBL-producing Enterobacteriaceae in 21% of Swedish samples 22 ; two further studies on Swedish gulls detected ESBL-producing E. coli in 2.0% and 6.4% of fecal samples, respectively23,24; and one study on Swedish Collared Flycatchers (Ficedula albicollis) found ESBL-producing bacteria in 0.0% of fecal samples. 25 Thus, while the results published in different articles vary, the occurrence of ESBL-producing bacteria has not been shown to exceed 21% in ducks sampled in Europe or in any birds sampled in Sweden. Rödiger et al. investigated a population of E. coli in wild Mallards in Germany over four consecutive winters and concluded that while the bacterial population was genetically dynamic and diverse, antibiotic resistance genes persisted within the population over time. 26
In this study, we assessed the detection rate of antimicrobial resistance in wild Mallards in an urban environment. The aim was to explore the overall prevalence and temporal variation of ESBL-producing E. coli and K. pneumoniae. This is novel as longitudinal studies on ESBL-producing bacteria in wild birds have previously not been published. This is also the largest study on ESBL-producing bacteria in Swedish wildlife to date. The large concentration of wild birds, urban location, vicinity to a large hospital, and frequent interactions between humans and birds add further relevance to an investigation of resistant bacteria, especially as such sites are common in cities globally. Given the sampling regime, we were able to assess methods for collection and storage, which are of importance to future studies on antimicrobial resistance in wild birds.
Materials and Methods
Collection of samples
Ethical approval for trapping and sampling was obtained from the Uppsala Animal Ethical Committee (Reference Number C228/12). A permit was obtained from the City of Uppsala to capture and a permit from Swedish Museum of Natural History to ring birds. Monthly sampling was performed at a pond in central Uppsala, Sweden, for 11 months; January to November of 2013. The pond, known as Svandammen (N59°51′16″ E17°38′25″), is located in the city center and in the direct vicinity of Uppsala University Hospital. Water is supplied to the pond from, and subsequently returned to, the nearby Fyris River. Throughout the year, numerous tourists and locals visit the pond, and many feed the ducks.
The number of Mallards present in the pond was estimated at each sampling. Fecal samples were collected both from captured Mallards (n = 52) and from fresh droppings (n = 761). In January, all samples were collected from captured birds; at the following samplings, samples from droppings were included due to difficulties in trapping birds. For a selection of droppings/birds sampled in February to April, duplicates (n = 123) and triplicates (n = 50), that is, multiple swabbings of the same dropping/bird were collected. In addition, bottom sediment from the pond was sampled monthly (n = 10). All samples were collected using sterile tipped applicators, put in lysogeny broth with glycerol, and stored at −80°C. At the January sampling, an additional sample replicate was collected for culturing without previous freezing (n = 20) using Amies Agar Gel Transport Swab with charcoal (Copan Italia S.P.A., Brescia, Italy).
Isolation of bacteria
Samples were enriched in brain–heart infusion broth with vancomycin 16 mg/L at 37°C for 16–24 hr and then spread on chromID ESBL plates (BioMérieux, Marcy l'Etoile, France). One colony each of presumed E. coli and K. pneumoniae was isolated. Species identity of isolates was primarily assessed using six biochemical tests: oxidation/fermentation glucose test, urease test, sulfide-indole-motility test (SIM), O-nitrophenyl-β-D-galactopyranoside test, Voges–Proskauer test, and culturing on cystine lactose electrolyte deficient agar plate. Isolates deviating from the expected profile were further analyzed using matrix-assisted laser desorption/ionization with time-of-flight mass spectrometry (MALDI-TOF) by the clinical microbiology department of Uppsala University Hospital using MALDI Biotyper (Bruker Daltonics, Billerica, MA).
Phenotypic analysis of antimicrobial resistance
In confirmed isolates of E. coli and K. pneumoniae, ESBL phenotype was assessed by disc diffusion testing on Müller-Hinton agar, using two discs containing cefpodoxime and cefpodoxime/clavulanic acid (Oxoid Ltd, Basingstoke, UK). 27 Only isolates susceptible to clavulanic acid, thus belonging to group 2be according to Bush et al., 28 were included in further analysis and are hereafter termed “ESBL.” In addition, susceptibility to carbapenems was analyzed by meropenem disc diffusion (Oxoid Ltd.), using the carbapenemase screening cutoff recommended by the European Committee on Antimicrobial Susceptibility Testing (EUCAST guideline on detection of resistance mechanisms v1.0, www.eucast.org/resistance_mechanisms).
Temperature-based grouping of samples
Considering conditions that enable Enterobacteriaceae to thrive, we grouped the samples based on the mean monthly air temperature in Uppsala 2013. Meteorological data were obtained from the Swedish Meteorological and Hydrological Institute. Accordingly, months were grouped into the six warmest (May to October 2013, mean temperature ≥7.4°C) and the six coldest (January to April and November to December 2013, mean temperature ≤4.5°C).
Statistical analysis
For statistical analysis, the two-tailed chi-square test was used with a cutoff value of p ≤ 0.05. GraphPad Prism software version 6.0e was used for all calculations.
Genotypic analysis of ESBL-producing isolates
ESBL-producing isolates were analyzed for CTX-M genes using previously described polymerase chain reaction (PCR) protocols. 29 Primarily, PCR was performed for genes belonging to the CTX-M-1 group; negative samples were subsequently tested for CTX-M-9, CTX-M-2, and CTX-M-8 group genes until a positive result was obtained. PCR products were cleaned using ExoSAP-IT (Affymetrix, Santa Clara, CA) and sequenced by Macrogen Europe, Amsterdam, The Netherlands. Samples found to harbor CTX-M-15 were screened for the pUUH239.2 plasmid using two singleplex PCRs as described earlier. 9 Isolates, which tested negative for all CTX-M genes, were further screened for the ESBL genes SHV, TEM, and OXA-1 using previously described multiplex PCRs. 30
Genotypic analysis of carbapenemases
Isolates with decreased susceptibility for meropenem were analyzed in two multiplex PCRs, using previously described methods 31 to screen for known carbapenemase genes. The annealing temperature was 57°C in the multiplex PCR screening for KPC, NDM, OXA, VIM, and IMP genes and 52°C in the multiplex PCR screening for AIM, GIM, SIM, and DIM genes.
Analysis of antibiotics in pond water
At all 11 monthly sampling occasions, 30 ml grab samples of surface water were obtained. They were frozen at −20°C and shipped on dry ice to the Department of Chemistry, Umeå University, Sweden. The samples were analyzed with a previously published multiresidue method based on online solid phase extraction liquid chromatography tandem mass spectrometry. 32 The method targets 105 pharmaceutical residues, including the following 12 antibiotics (limit of quantification is given in parentheses): azithromycin (40 ng/L), ciprofloxacin (10 ng/L), clarithromycin (3 ng/L), clindamycin (3 ng/L), erythromycin (20 ng/L), norfloxacin (20 ng/L), ofloxacin (3 ng/L), oxytetracycline (10 ng/L), roxithromycin (15 ng/L), sulfamethoxazole (15 ng/L), tetracycline (20 ng/L), and trimethoprim (3 ng/L).
Results
Distribution of collected samples
The number of Mallards residing by the pond at sampling occasions varied between 46 (in May) and 570 (in January). A total of 813 fecal samples were collected, the monthly number of samples ranging from 20 to 100, as outlined in Table 1.
Detection rate of ESBL-producing bacteria
Overall, phenotypical analysis revealed 411 ESBL-producing E. coli or K. pneumoniae in 386 of 813 fecal samples (47%). These results are detailed in Table 2 and Fig. 1. Note that, in regard to our samples, the term ESBL is reserved for enzymes of group 2be according to Bush et al. 28 ; our methods do not allow for detection of other types of ESBLs (such as pAmpC). In total, 386 isolates of ESBL-producing E. coli and 25 isolates of ESBL-producing K. pneumoniae were obtained from 386 samples; thus, ESBL-producing K. pneumoniae was not found in any sample without simultaneous presence of ESBL-producing E. coli. The monthly proportion of samples harboring ESBL-producing bacteria ranged from 4.2% in May to 84% in July. ESBL-producing K. pneumoniae was isolated in 3.1% of samples, and the highest detection rate was observed in July (15%). Grouping of samples according to mean air temperature revealed that 322 out of 493 (65%) fecal samples collected during the six warmest months (May to October) were positive for ESBL-producing E. coli. During the five coldest months (January to April and November), 64 out of 320 (20%) collected samples were positive for ESBL-producing E. coli. Statistical analysis revealed a significant difference (χ2 = 159.8, df = 1, p < 0.0001) between the two groups. There was also a significant difference in the detection rate of K. pneumoniae when comparing the warmest (4.6%) and the coldest months (0.6%) (χ2 = 10.63, df = 1, p = 0.0011).
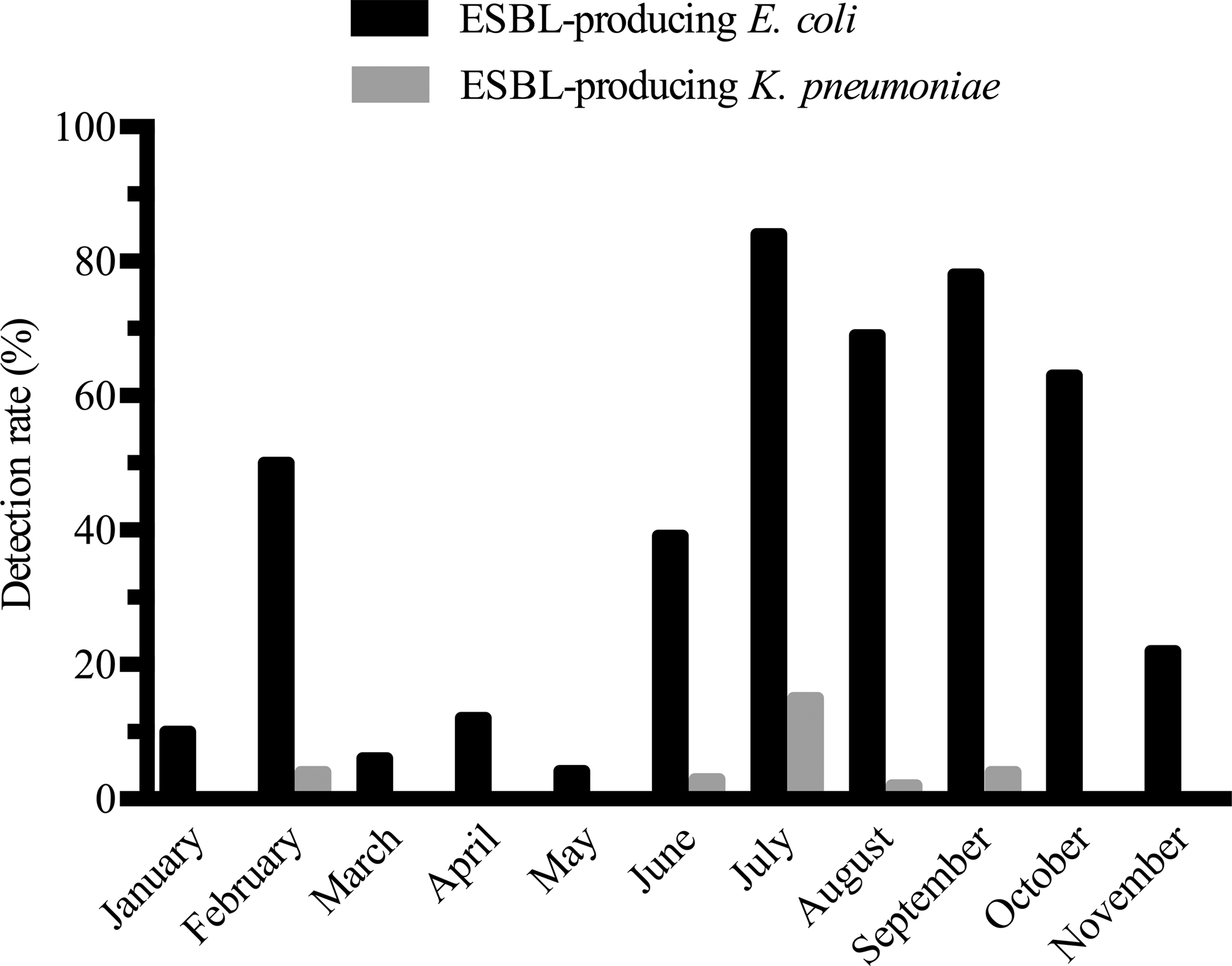
Monthly detection rate of ESBL-producing Enterobacteriaceae in fecal samples. ESBL, extended spectrum β-lactamase.
ESBL, extended spectrum β-lactamase.
Distribution of CTX-M genotypes
Among the 411 ESBL-producing isolates from fecal samples, six different CTX-M genotypes were found. The most prevalent was blaCTX-M-15, harbored by 295 (72%) of isolates, the monthly proportion ranging from 50% to 100%. Another 34 isolates (8.3%) harbored genes that we designated blaCTX-M-14-like (as our CTX-M-9 group PCR yielded products too short to separate blaCTX-M-14, blaCTX-M-17, blaCTX-M-18, blaCTX-M-24, blaCTX-M-27, blaCTX-M-38, and blaCTX-M-48) and 33 isolates (8.0%) harbored blaCTX-M-1. In addition, blaCTX-M-55, blaCTX-M-65, and blaCTX-M-3 were also detected. Seven isolates (1.7%) were found negative for all tested blaCTX-M gene groups. All seven CTX-M negative isolates were found to harbor blaSHV genes; in addition, four isolates harbored blaTEM and one isolate blaOXA-1. These isolates were not further characterized. The seasonal distribution of blaCTX-M genes is detailed in Table 3. The two most prevalent genes among the 25 K. pneumoniae isolates were blaCTX-M-15 detected in 12 isolates (48%) and blaCTX-M-65 detected in seven isolates (28%). In samples where ESBL-producing E. coli and K. pneumoniae coexisted, the resistance genes of the two isolates had identical genotype in 13 of the cases (52%). The outbreak plasmid pUUH239.2 was not identified in any of the blaCTX-M-15-harboring isolates.
Sediment samples
Five out of 10 (50%) sediment samples (from February, May, June, August, and September) were positive for ESBL-producing E. coli. Four of these isolates were found to harbor blaCTX-M-14-like genes, and one isolate was found to harbor the blaCTX-M-15 gene. No ESBL-producing strains of K. pneumoniae were identified in sediment samples.
Carbapenemase analysis
A decreased susceptibility to meropenem was demonstrated in 18 out of 391 (4.6%) ESBL-producing E. coli and 0 out of 25 (0.0%) ESBL-producing K. pneumoniae from fecal and sediment samples. All 18 isolates with decreased susceptibility to meropenem were negative for the tested carbapenemase genes (KPC, NDM, OXA, VIM, IMP, AIM, GIM, SIM, and DIM). None of the isolates fulfilled the definition of meropenem resistance as per EUCAST clinical breakpoints for bacteria v7.1.
Replicate analysis and impact of freezing
In the 173 samples where duplicates were collected, ESBL-producing E. coli or K. pneumoniae was isolated from 29 of the first replicates (17%). When the duplicates were analyzed, the number of ESBL-positive samples increased to 38 (22%), an increase by a factor of 1.3. In 20 of the samples, ESBL-producing bacteria were isolated from both replicates. Among the 173 double-tested samples, a third replicate was analyzed in a subset of 50 samples. In these 50 samples, analysis of the first and second replicates yielded 25 ESBL-producing isolates (50%), and the inclusion of a triplicate did not increase the number of ESBL-positive samples. ESBL-producing E. coli was isolated in 6 of the 20 samples (30%) analyzed without a freeze cycle. Replicates of the same samples, analyzed after long-term storage at −80°C, yielded two ESBL-producing isolates (10%). For one of the samples in which the frozen replicate was positive, the nonfrozen replicate was negative. The extra positive samples found by culturing replicates and nonfrozen samples were not included in analyses of detection rate or genotype.
Analysis of antibiotics in pond water
None of the 12 tested antibiotics were detected in any of the 11 pond water samples.
Discussion
Our study demonstrates that Mallards in central Uppsala, Sweden, are frequently colonized by ESBL-producing Enterobacteriaceae and that there is considerable temporal variation in the detection rate of resistant bacteria. Genotypic analysis of our isolates revealed a dominance of blaCTX-M-15. Interestingly, in samples where ESBL-producing E. coli and K. pneumoniae coexisted, the blaCTX-M genotypes were different in 48% of samples, demonstrating carriage of multiple ESBL genotypes within a host bird.
Previous research has demonstrated significant seasonality in human carriage of and community-acquired infections caused by Enterobacteriaceae, including ESBL-producing strains, such that rates increase at higher outdoor temperatures.33–37 We detected significantly higher rates of ESBL-producing E. coli during months with high mean air temperature than in months with low mean air temperature. For ESBL-producing K. pneumoniae, the pattern was similar. To our knowledge, this phenomenon has not been previously described in wildlife, and further research is required to determine whether this is an annual phenomenon. The sample sizes per season are borderline too small to draw conclusions regarding seasonal prevalence.
The mechanisms for increased dissemination of Enterobacteriaceae at higher outdoor temperatures are not known. The detection rate may be increased due to enhanced bacterial growth and improved survival of bacterial cells in the environment, facilitating transmission of bacteria between birds and/or increased recovery at sampling.
As demonstrated by the observed temporal variation, the result of a single sampling may not be representative for the overall prevalence of resistant bacteria within a bird population. Future studies on the prevalence of Enterobacteriaceae in bird populations must therefore take into account the possibility of seasonal variations.
The prevalence of 47% ESBL-producing E. coli in our study is drastically higher than in previous studies on birds in Sweden21,23–25 and on ducks in Europe.18–20 Results similar to ours have been reported from the Netherlands and Spain in a study on ESBL-producing Enterobacteriaceae in gulls from nine European countries. 22 Note that care should be taken when comparing results of different studies, especially with regard to the temporal variation found in our results. Interestingly, the only previous study on Mallards at our sampling site, performed in autumn of 2008, did not detect any ESBL-producing bacteria in 17 birds. 21 Although the sample size is small, this may suggest that the occurrence of ESBL-producing bacteria at the site is a more recent phenomenon.
Our method is selective for enzymes susceptible to clavulanic acid, and thus, the true prevalence of ESBLs, including plasmid-borne AmpC-type β-lactamases, may be even higher. The high detection rate in our study is remarkable, as Sweden is a country with comparatively low use of antibiotics in human and veterinary medicine and low rates of resistant bacteria in clinical patients.3,38 The average incidence of ESBL-producing Enterobacteriaceae reported in Sweden was 91 cases per 100,000 inhabitants in 2014, and Uppsala county ranked slightly below the national average. 3 However, rates are increasing, as demonstrated by studies on healthy preschool children in Uppsala where the prevalence of ESBL-producing Enterobacteriaceae rose from 2.9% in 2010 to 20% in 2016.6,7 The absence of carbapenemase-producing Enterobacteriaceae among our isolates is in accordance with the limited spread of such bacteria in Sweden, both among humans and in the environment.
Our findings reveal that urban Mallards in Sweden harbor ESBL-producing E. coli and K. pneumoniae with genetic characteristics similar to Swedish human isolates. Egervärn et al. report that in E. coli isolated from healthy humans, blaCTX-M-15 was the most common ESBL gene, followed by blaCTX-M-14 and blaCTX-M-1, which corresponds well to our results.3,5 They also found that there are three largely isolated populations of ESBL-producing E. coli in Sweden: one in imported foods; one in domestic foods, livestock, and poultry; and one shared by the human population, wastewater, river water, and wild gulls. Likewise, a study from France concludes that humans and wild gulls share populations of E. coli. 39 Our findings support the hypothesis that human and wildlife populations of Enterobacteriaceae communicate and highlight the need for efficient wastewater treatment, a setting where wild birds are exposed to human fecal material.
While our sampling site is a hub of human activity, it is located upstream of wastewater treatment facilities, and no antibiotic residues were found in the pond water. This suggests that resistant bacteria have been imported to, rather than selected for, at the site. While the possibility of direct communication between the bacterial populations of Uppsala University Hospital and the nearby sampling site has not been thoroughly investigated, we have not found any indications of such communication, as the pUUH239.2 plasmid could not be identified in our isolates. Phylogenetic analyses of isolated bacteria were not performed, and we cannot discern whether isolates from the same genotype mainly represent the same strain (indicating clonal spread of resistant bacteria between birds at the pond) or different strains (indicating multiple introductions of resistant bacteria to the pond). Regardless, the results clearly demonstrate that urban ponds can hold high concentrations of antibiotic resistant bacteria and, thus, may play an important role in the dissemination of such bacteria between birds, with the potential for geographic spread through bird migration. Similar environments are common in cities globally, and it is important to identify and monitor them. In addition, the possibility of transmission of resistant bacteria between birds and humans in such settings cannot be excluded, and further research asserting this aspect is of significance to determining the risk for public health.
Migrating birds have been suggested to act as vectors for the dissemination of antimicrobial resistance.11–17 Our sampling site accumulates large gatherings of Mallards during the postbreeding molt, the migration periods, and especially in wintertime. Data from ringed Mallards reveal that while most Swedish Mallards only perform regional movements over the year, a number of Mallards spending the summer in Sweden migrate to the North Sea coast in winter, whereas the wintering population includes Mallards breeding in Finland or western Russia. 40 The latter is illustrated in Fig. 2 (map and data were obtained by personal communication with Thord Fransson at the Swedish Bird Ringing Centre, Swedish Museum of Natural History). While this is presumably true also for the Mallards of Uppsala, it is unlikely that the influx and efflux of migratory birds alone explain the large variation in detection rate of resistant bacteria across the year. However, it highlights the risk of transmission of resistant bacteria from urban ponds to other environments through bird migration.
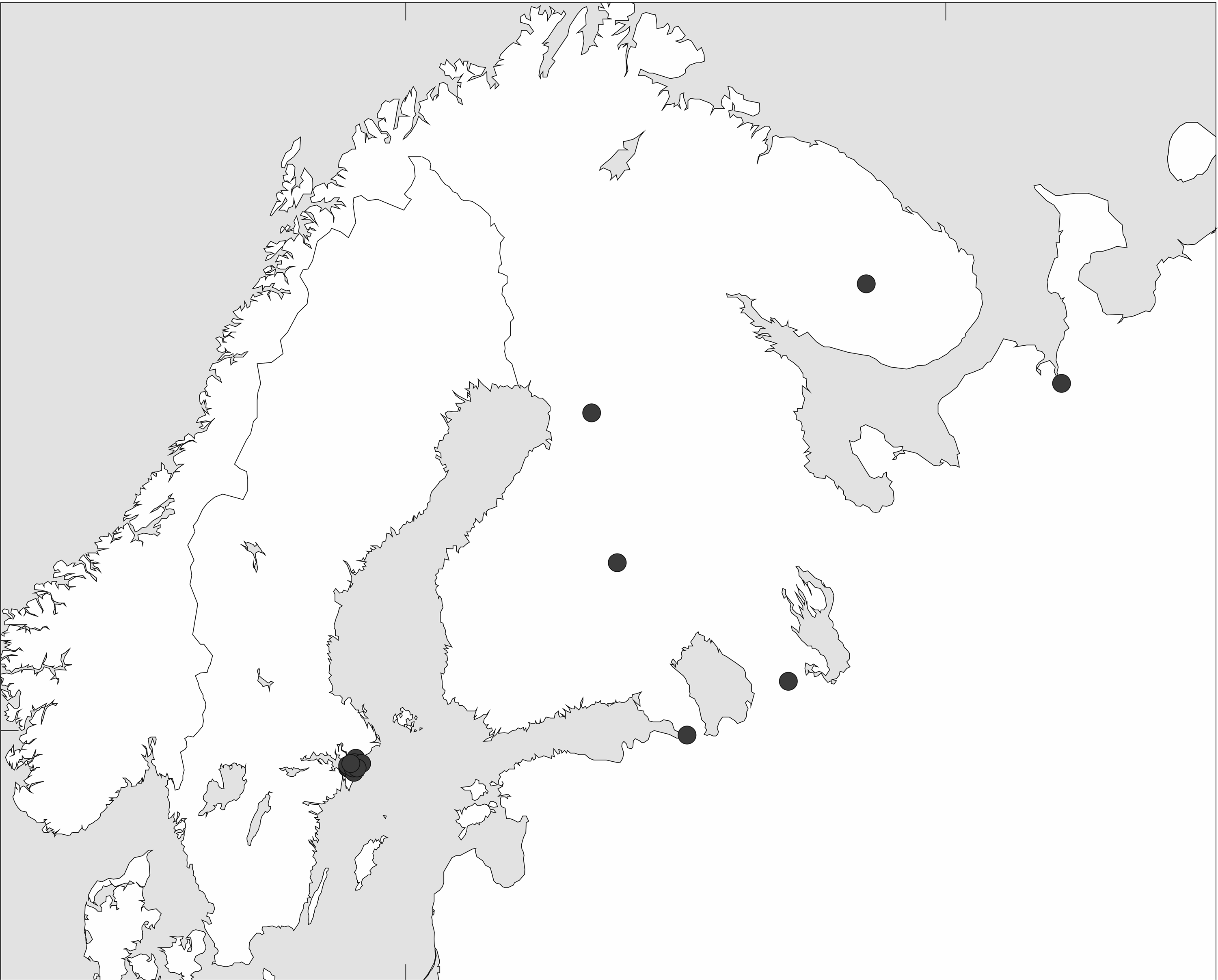
Recoveries of male Mallards, ringed in Stockholm, Sweden, during the period November to March and recovered during the period April to July (n = 34).
In this study, the analysis of duplicates of fecal samples increased the detection rate of ESBL-producing E. coli by a factor of 1.3. Analysis of triplicates did not increase the number of ESBL-positive samples further. Our findings demonstrate that, while use of a single sample could underestimate the prevalence of ESBL-producing E. coli, use of duplicates appears to be the optimal approach. To our knowledge, this methodological aspect has previously not been evaluated in regard to ESBL-producing Enterobacteriaceae in wild birds. In a comparison of sample replicates stored at −80°C and replicates analyzed without a freeze–thaw cycle, we observed a trend of increased detection of ESBL-producing E. coli in the nonfrozen replicates. While our findings should be interpreted with caution due to the limited number of samples, a decreased viability of E. coli after long-term storage at −80°C is suggested. Consequently, it is probable that the predominant approach in this study, as in many others, has underestimated the prevalence of ESBL-producing bacteria. We encourage researchers conducting future studies on Enterobacteriaceae in fecal samples to consider that duplicate samples and avoidance of a freeze–thaw cycle may increase the sensitivity of their methods.
A limitation of our study is the predominant collection of samples from fresh droppings. Compared to the sampling of captured Mallards, there is an increased risk for environmental contamination and we cannot know to what extent individual birds have been sampled multiple times at one sampling. We have sampled a large number of droppings, collected around the entire circumference of the pond and on its island, to reduce the risk of influence on our results by this limitation.
Conclusions
Our findings demonstrate a higher-than-expected prevalence of ESBL-producing E. coli and K. pneumoniae in wild Mallards in an urban Swedish environment. ESBL-producing bacteria were present throughout the year; however, temporal variation was profound, and the detection rate was significantly higher in the six warmest months of the year (May to October). Urban ponds may serve as important natural reservoirs for antimicrobial resistance. The distribution of CTX-M genes in our isolates corresponds well with Swedish human isolates of ESBL-producing Enterobacteriaceae, indicating communication between the genetic pools of ESBLs in humans and wild birds. However, the extent, direction, and mechanisms of transmission are not known, and further research is essential to understand the potential public health risks. Future studies on ESBL-producing Enterobacteriaceae in wild birds must take into account the possibility of temporal variation in prevalence; in addition, the use of duplicate or nonfrozen samples should be considered to produce more reliable results.
Footnotes
Acknowledgments
The authors are grateful to Swedish Research Council (Vetenskapsrådet, Grant No. 2016-02606) and Olle Engkvist Byggmästare's and Karin Korsner's Foundations for funding. The authors also thank Kristine Lindqvist and Carolina Stigwall who helped collect samples, as well as Badrul Hasan and Jenny Isaksson for assistance in the laboratory.
Disclosure Statement
No competing financial interests exist.