
Abstract
This study aimed to evaluate the antibiogram and antibiotic resistance genes (ARGs) of Vibrio isolates recovered from a marine fish (Argyrosomus japonicus) and water samples from two commercial dusky kob aquaculture farms and the Kariega estuary, South Africa, and to evaluate these findings for their public health implications. A total of 277 molecularly confirmed Vibrio isolates consisting of 126 Vibrio fluvialis, 45 Vibrio vulnificus, 30 Vibrio Parahaemolyticus, and 76 vibrios belonging to species of the genus other than Vibrio cholerae were subjected to susceptibility testing to 15 antibiotics by the disc diffusion method. Multiple antibiotic resistance index (MARI) was used to determine the antibiotic resistance-associated health risk, while polymerase chain reaction was used to evaluate the presence of 14 ARGs for nonsusceptible strains. Highest resistances were recorded to amoxicillin (76.2%), ampicillin (67.5%), erythromycin (38.3%), and doxycycline (35.0%), while susceptibilities were highest to gentamicin (100%), followed by norfloxacin (97.8%), florfenicol (90.3%), tetracycline (87.7%), and chloramphenicol (87.4%). We recorded a 58.5% multidrug resistance (resistance to ≥2 antimicrobial classes). MARI did not vary significantly between sites (p > 0.05); however, values of >0.2 were recorded in 40% (108/277) of all strains tested. ARG markers, ampC, blaOXA, tetA, tetM, dfr1, sul1, sul2, ermB, nptII, strA, and SXT integrase, were detected in one or more strains with ermB (82.5%), sul2 (53.8%), strA (44%), dfr1 (42.3%), and tetM (38.3%) being the most abundant. Healthy marine finfish (dusky kob) and their environment can serve as reservoirs for antibiotic resistant vibrios and ARGs, which could be disseminated to humans and other susceptible bacteria and this therefore becomes a public health concern.
Introduction
A
In aquaculture, the control of antibiotic usage varies widely from country to country. 3 Most developed countries either have registered antibiotics for bacterial disease control, with well-defined regulations regarding their usage, or no longer permit the registration of products for nontherapeutic purposes such as growth promoters.4,5 However, about 90% of aquaculture industries are located in developing countries with weaker control strategies. Although antibiotics play an important role in the prevention and treatment of bacterial diseases both in humans and food animals, the indiscriminate use of these antimicrobials has led to the emergence of antibiotic-resistant bacteria and antibiotic resistance genes (ARGs) in the environment. Members of the genus Vibrio are abundant in the marine environment and may contaminate edible fish. Vibrio cholerae, Vibrio parahaemolyticus, Vibrio Vulnificus, and Vibrio fluvialis are the most significant human pathogens of the genus that may cause mild to potentially fatal foodborne illnesses, especially from consumption of contaminated seafood.6–8 The acquisition of ARGs by these pathogenic bacteria is of concern as these may cause treatment failure and difficulties in human disease management. Previous studies have reported antibiotic resistance or resistance genes in vibrios isolated from shrimps9,10 and other seafood.11,12
These pathogenic bacteria with acquired ARGs may serve as reservoirs of resistance genes to susceptible bacteria. The accumulation of antibiotic residues in edible fish tissues may also alter human intestinal flora and cause food poisoning or allergies. 13 Some of these residues may be detected from several months 14 up to a few years posttreatment with antimicrobials. 15 The World Health Organization considers the emergence of drug-resistant bacteria, particularly multidrug-resistant strains, to have critical consequences for human medicine. 16 Data on the actual supply, consumption concentrations, and volumes of antimicrobials utilized in South African animal health and aquaculture are scanty. It is imperative to continuously monitor the drug resistance pattern of potentially pathogenic bacteria species as to determine the risk they may pose to human health. We therefore evaluated the susceptibility patterns of Vibrio strains (isolated from water and fish from two commercial dusky kob aquaculture farms and the Kariega Estuary) to antibiotics commonly used in the treatment of fish and human vibriosis, as well as their associated ARG markers.
Materials and Methods
Bacterial isolates and growth
A total of 277 Vibrio isolates (126 V. fluvialis, 45 V. vulnificus, 30 V. parahaemolyticus, and 76 vibrios belonging to other species of the genus other than V. cholerae) randomly selected from a pool of 606 vibrios (193 V. fluvialis, 74 V. vulnificus, 33 V. parahaemolyticus, and 306 belonging to other species of the genus), isolated from 80 water samples and 120 dusky kobs in a previous study, 17 were used in this study. Forty-four isolates were from Kariega estuary (33°41′S, 24°44′E), 147 were from Farm 1 (13.0670°N, 59.5712°W), and 86 were from Farm 2 (32.9638°S, 27.8789°E), all located in the Eastern Cape province, South Africa. All strains were previously maintained in Tryptic Soy Broth supplemented with 25% glycerol and stored at −80°C following molecular identification of the isolates. The frozen cultures were resuscitated on brain–heart infusion agar plates, and the plates were incubated at 37°C for 18–24 hr before antibiotic susceptibility testing and antibiotic resistance gene detection.
Antibiotic susceptibility test
Antimicrobial susceptibility of Vibrio isolates to 15 antibiotics belonging to nine classes was performed according to the standard guidelines recommended by Clinical Laboratory Standards Institute (CLSI). 18 An inoculum for each isolate was prepared by emulsifying colonies from an 18-hr pure culture in 3 ml of sterile normal saline (0.85%), and the turbidity was adjusted to 0.5 McFarland standard (0.5 ml of 1% w/v BaCl2 and 99.5 ml of 1% v/v H2SO4) equivalent to 1.0 × 108 cfu/ml. The bacterial suspension was uniformly streaked on Mueller Hinton agar plates using sterile swabs and allowed for 3–5 min before introduction of antibiotic discs. A panel of 15 antibiotic-impregnated discs (Mast Diagnostics, United Kingdom), namely tetracycline (30 μg), erythromycin (15 μg), amoxicillin (15 μg), streptomycin (10 μg), gentamycin (10 μg), ciprofloxacin (5 μg), Chloramphenicol (30 μg), trimethoprim-sulfamethoxazole (1.25/23.75 μg), norfloxacin (10 μg), ampicillin (10 μg), doxycycline (30 μg), nalidixic acid (30 μg), florfenicol (30 μg), imipenem (15 μg), and cefoxitin (30 μg), were placed on inoculated plates with a mast disc dispenser (Mast Diagnostics). These antibiotics were chosen based on their use in the treatment of Vibrio infections in humans and animals, as well as their use in aquaculture. Plates were incubated at 37°C for 24 hr, and the diameters of the zones of inhibition were measured and results interpreted as susceptible (S), intermediate (I), or resistant (R) according to CLSI interpretive criteria.18,19 Resistance to florfenicol was interpreted as earlier reported. 20 A multiple antibiotic-resistant phenotype was defined as resistance to ≥2 antibiotic classes.
Determination of multiple antibiotic resistance index/multiple antibiotic resistance phenotype
Multiple antibiotic resistance index (MARI) is an important epidemiological tool used in assessing human health risk in relation to antibiotic resistance. This tool aids in differentiating high-risk (value of >0.2) from low-risk (value of <0.2) contamination with bacteria of potential hazard to humans in relation to antibiotic resistance. 21
MARI for an isolate was computed as:
MARI = a/b, where “a” represents the number of antibiotics to which that isolate was resistant and “b” the total number of antibiotics to which the isolate was exposed, 21 while the MARI for a given sample of several isolates for each site was also derived using the mathematical formula
MARI = a/b.c, where “a” is the aggregate antibiotic resistance of all the isolates, “b” is the number of antibiotics tested, and “c” is the number of isolates in the sample. 21
Detection of antibiotic resistance genes
For all cultures maintained at −80°C, genomic DNA was isolated by suspending isolated colonies from an 18-hr pure culture in 200 μl of sterile distilled water. Cells were lysed by boiling for 15 min in a digital Accu dri-block (Lasec, South Africa) and cell debris removed by centrifugation at 13,000 g for 5 min. The supernatant was used directly as template in the polymerase chain reaction (PCR) assay or stored in 50 μl aliquots at −20°C until use. Nonsusceptible strains to various antibiotics were subjected to detection of ARGs by PCR using specific primers in a C1000 Touch Thermal Cycler (Bio-Rad, Singapore, Malaysia). Table 1 shows the primer sequences and cycling conditions used for the detection of the ARGs. Each amplification was performed in a 25 μl reaction, consisting of 5 μl template DNA, 0.5 μl of each of the oligonucleotide (Inqaba Biotec, Pretoria, South Africa), and 12.5 μl of OneTaq Quick-Load 2 × PCR Master mix (New England BioLabs, Ipswich, United Kingdom) and an appropriate volume of nuclease-free water (New England BioLabs). PCR amplified products were separated in ethidium bromide (0.5 μg/ml) stained with 1% agarose gels by electrophoresis at 100 V for 45 min. Gels were visualized using a UV transilluminator (Alliance 4.7 XD-79; UVITEC, Cambridge, United Kingdom).
Statistical analysis
A chi-square test and Goodman and Kruskal's tau test were used to determine the relationships between resistances obtained for each antibiotic from different sampling sites. A p-value <0.05 was considered statistically significant.
Results
Antimicrobial resistance
Generally, the antibiotic susceptibilities of Vibrio isolates to various antibiotics varied considerably. Of the 277 isolates analyzed, 262 (94.6%) were resistant to at least one antibiotic. Overall, the antimicrobial susceptibility pattern revealed gentamicin (100%), norfloxacin (97.8%), florfenicol (90.3%), tetracycline (87.7%), chloramphenicol (87.4%), streptomycin (78.7%), imipenem (76.5%), sulfamethoxazole–trimethoprim (76.2%), and nalidixic acid (74.4%) as the most effective antibiotics against Vibrio strains tested (Table 2). In contrast, resistance to cefoxitin (28.9%), doxycycline (35.0%), and erythromycin (38.3%) was higher, with the highest resistance recorded to ampicillin (67.5%) and amoxicillin (76.2%). Although the total percentage of resistance recorded to ciprofloxacin was very low (1.8%), almost half (44.8%) of the nonsusceptible isolates showed an intermediary resistance pattern. Considering the three human pathogenic species, the high total resistance to amoxicillin was mostly contributed by V. vulnificus (84.4%) and V. parahaemolyticus (56.7%) with a much lower resistance (8.7%) of V. fluvialis to the drug. V. vulnificus resistance to erythromycin, streptomycin, trimethoprim–sulfamethoxazole, and cefoxitin was also significantly higher (p < 0.05) than those recorded for V. parahaemolyticus and V. fluvialis. Similarly, high resistance (35.6% for V. vulnificus and 46.7% for V. parahaemolyticus) was detected to doxycycline, with a lower resistance (13.5%) of V. fluvialis to the antibiotic. Table 2 shows the percentage susceptibility profiles of 277 Vibrio isolates to 15 antibiotics.
I, intermediate; R resistant; S, susceptible.
When we compared resistances among the isolates obtained from the three sites, there were marked differences in the antimicrobial resistance patterns observed from isolates of the different sites (Fig. 1). Vibrio isolates recovered from Farm 2 showed significantly higher (p < 0.05) resistance to tetracycline, doxycycline, erythromycin, trimethoprim–sulfamethoxazole, chloramphenicol, florfenicol, and imipenem with significantly lower (p < 0.05) resistances recorded to ampicillin, nalidixic acid, florfenicol, and cefoxitin than those from Farm 1 and Kariega estuary (Fig. 1). The only antimicrobial to which there was a significantly higher (p < 0.05) resistance in isolates from Kariega estuary compared to other sites was cefoxitin. None of the strains recovered from Kariega estuary were resistant to ciprofloxacin, norfloxacin, nor tetracycline. The percent resistances obtained for each antibiotic for Vibrio isolates recovered from the three sites are shown in Figure 1.
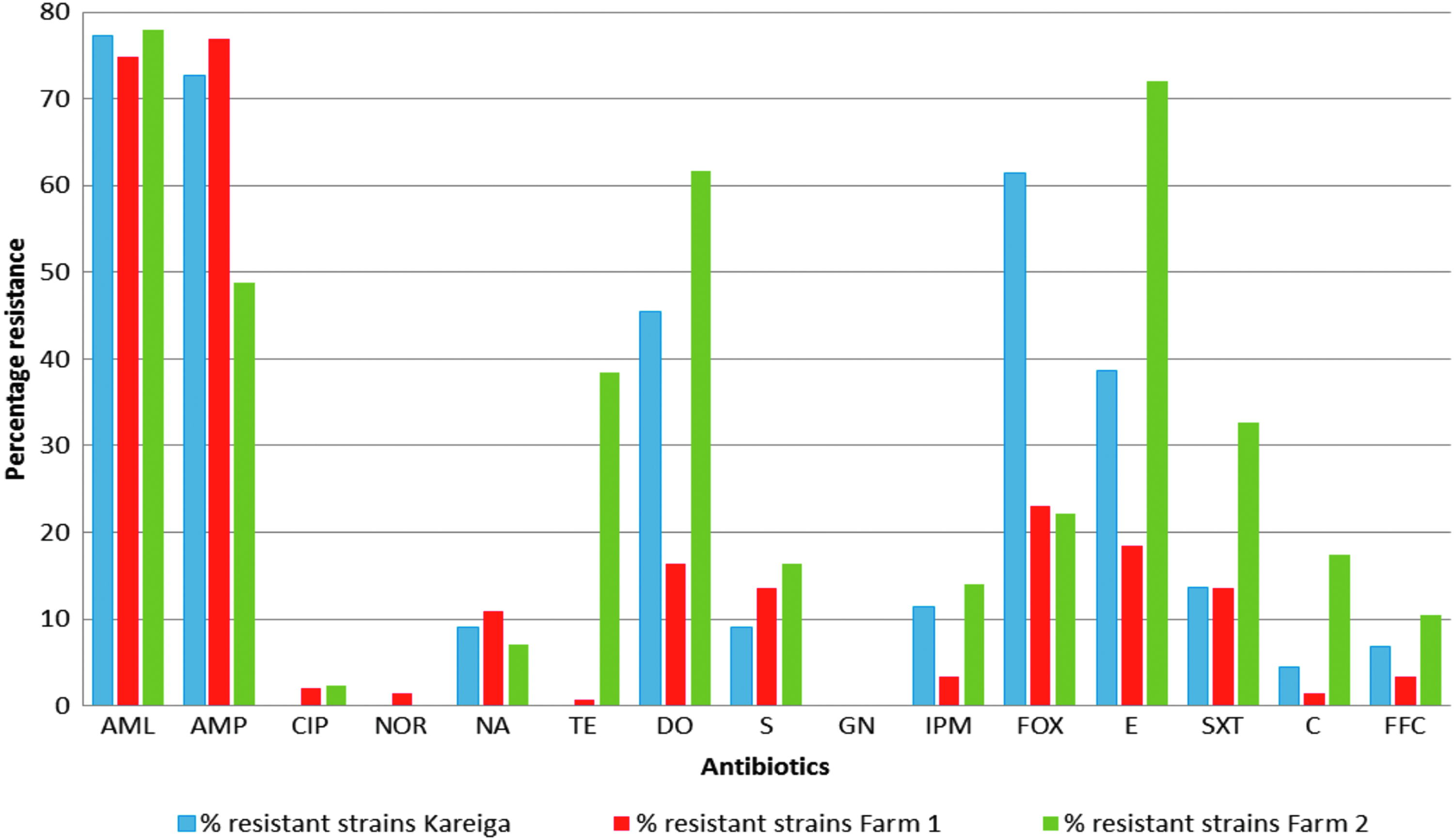
Percentage of antibiotic resistance of 277 Vibrio strains. AML, amoxicillin; AMP, ampicillin; C, chloramphenicol; CIP, ciprofloxacin; DO, doxycycline; E, erythromycin; FFC, florfenicol; FOX, cefoxitin; GN, gentamicin; IPM, imipenem; NA, nalidixic acid; NOR, norfloxacin; TE, tetracycline; S, streptomycin; SXT, trimethoprim–sulfamethoxazole.
Multiresistance/MARI/multiple antibiotic resistance phenotypes
Simultaneous resistance to various antimicrobials was high (84.5%; 234/277), with a 58.5% (162/277) multidrug resistance (defined as resistance to at least two classes of antimicrobials) recorded (Table 3).
ARG, antibiotic resistant gene.
As earlier described, 21 a MARI of >0.2 differentiates isolates between antibiotic resistance from low-risk and high-risk sources. In this study, the MARI ranged from 0.0 to 0.67. Forty percent (108/277) of isolates fell into a MARI of >0.2, while 42.2% (117/277) belonged to the <0.2 group. Fifty-two isolates (18.8%) had MARI = 0.2. Assessment of the aggregate individual resistances of isolates from each sampling location revealed that Kariega Estuary had the highest percentage of vibrios; this could possibly indicate areas of high antibiotic usage/contamination (MARI = 0.24), this was followed by Farm 2 (MARI = 0.23), and Farm 1 had the lowest percentage (MARI = 0.21). However, the differences in MARI between sampling locations were not significant (p > 0.05).
Considering multidrug-resistant isolates only, the compilation of the multiple antibiotic resistance phenotypes resulted in 23, 39, and 47 different patterns exhibited by isolates from Kariega, Farm 1, and Farm 2, respectively. One isolate from Farm 2 showed resistance to 10 of the 15 antimicrobials tested, exhibiting the phenotype AMPR-AMLR-TER-ER-SR-CR-SXTR-DOR-NAR-IPMR.
Prevalence of antimicrobial resistance genes
PCR assays carried out to detect the presence of antibiotic-resistant gene markers revealed that 11 (ampC, blaOXA, tetA, tetM, dfr1, sul1, sul2, ermB, nptII, strA, and SXT) of the 14 genes tested occurred in one or more Vibrio strains, with ermB (82.5%), sul2 (53.8%), strA (44%), dfr1 (42.3%), and tetM (38.3%) being the most abundant. Table 3 shows the prevalence of ARGs detected in the study.
Genes (dfr1, Sul1, and Sul2) coding for sulfonamide resistance were detected in 58.3% (7/12), 25% (3/12), and 50% (6/12) of the V. fluvialis isolates, respectively, while only dfr1 (33.3%; 4/12) and sul2 (66.7%; 8/12) genes were detected in the V. vulnificus isolates. None of these resistance genes were found in the nonsusceptible V. parahaemolyticus strains.
Among the genes that code for streptomycin (aminoglycosides) resistance, strA was the most commonly detected gene as it occurred in the V. vulnificus, V. parahaemolyticus, and V. fluvialis strains. In contrast, the nptII was detected only in V. vulnificus and V. fluvialis, while aacC2 was not found in any of the Vibrio strains. The gene coding for integrase of the SXT element was only found in V. parahaemolyticus and was detected in one out of the four resistant isolates. Of all the three genes tested which code for erythromycin (macrolide) resistance, namely, ermA, ermB, and ermC, only ermB was detected in all three Vibrio species at high frequencies. β-lactam resistance, encoded by the ampC and blaZ AGRs, was detected in all three Vibrio spp. For the genes coding for tetracycline resistance, tetM was detected in all the Vibrio species, while tetA was only detected in the resistant V. parahaemolyticus isolates. Figure 2 shows representative gels for some antibiotic resistance genes detected (combined from different PCR assays).
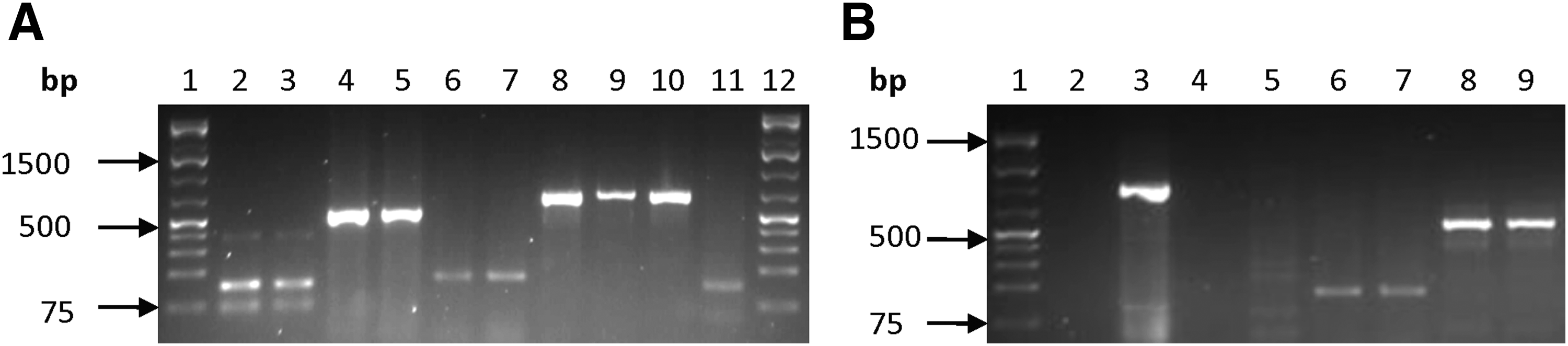
Representative gel showing some antibiotic resistant genes detected.
Discussion
Antibiotic sensitivity patterns among Vibrio isolates and MARI
It is necessary to continuously assess antibiotic resistance patterns of pathogenic bacteria in various foods and food environments as part of monitoring the spread and emergence of bacterial resistance. This helps in determining the health risk associated with subsequent infection when humans consume possibly contaminated food or water. Although intrinsic antibiotic resistance may occur, it is less likely to lead to high frequencies of resistance. An increase in bacterial resistance to an antimicrobial generally points to the widespread and indiscriminate use of these antimicrobials, while low frequency in bacterial resistance may be related to low contamination with antibiotics in the area. In this study, antimicrobial resistance results revealed that most (84.5%) Vibrio isolates were resistant to more than one antimicrobial, with over half of the total isolates (58.5%) being multidrug resistant. The high resistances revealed by Vibrio isolates and individual species to β-lactam antibiotics (ampicillin and amoxicillin) are similar to those reported in other studies.12,31–34 However, our results of <50% resistance of V. vulnificus to erythromycin and sulfamethoxazole, the resistance by V. parahaemolyticus to erythromycin and V. fluvialis to cotrimoxazole, erythromycin, and chloramphenicol are contrary to that reported by Okoh et al. 34 Although resistance to streptomycin was considered to be low, our findings revealed a higher percent resistance than has been reported before.12,35 The percentage of resistance recorded to ciprofloxacin might have been low (1.8%), but almost half (44.8%) of the nonsusceptible isolates showed an intermediary resistant pattern which possibly may be completely resistant to the antibiotic overtime.
In this study, gentamicin was the most effective antibiotic to vibrios as all isolates were susceptible to the drug. This result is similar to those earlier reported.12,32–34 The high susceptibility to gentamicin, together with a >90% susceptibility to norfloxacin and florfenicol, could be an indication of fairly low contamination of the aquatic environment and consequently its inhabitants by these antibiotics. In South Africa, florfenicol (AQUAFLOR, NUFLOR) is the only antibiotic currently registered for use in aquatic animals, although other antibiotics such as oxytetracycline hydrochloride and oxytetracycline dehydrate have been approved for extra-label uses against fish diseases under veterinarian supervision. 36 Low resistance (6.1%) to florfenicol is therefore encouraging. However, frequent monitoring and effective action to maintain the low levels of resistance to this antibiotic, particularly in aquaculture settings, are necessary to prevent further development and spread of resistance to this antibiotic.
Doxycycline, ciprofloxacin, and norfloxacin are among first line drugs administered against infections caused by vibrios. Based on our results, while ciprofloxacin and norfloxacin appear to be effective against the isolates tested, it is concerning to record resistances of 35.6% (V. vulnificus) and 46.3% (V. parahaemolyticus), respectively, to doxycycline. In addition, despite the absence of antibiotics in fish meals (as testified by the fish farmers of both Farm 1 and 2), the increased levels of antibiotic resistance to cefoxitin, doxycycline, erythromycin, ampicillin, and amoxicillin are an indication of antibiotic residues in the environment. This could possibly be from human and animal waste, runoffs from farmlands, or hospital waste that finally ends up in the estuaries or the residues could be from larval growth stages of fish in aquaculture. Although the culture of finfishes may not include antibiotics, most larval stages do include antibiotic treatment to increase survival rates from pathogenic bacteria. 37 The transfer of antibiotic-resistant bacteria's DNA from fish hatcheries to other pathogenic bacteria has been documented. 38 Subsequently, these are often associated with environmental and human health problems, including bacterial resistance and persistence of disease in the aquatic environment. 13 In turn, through the food chain or contact with contaminated water, humans become infected with resistant bacteria. Antibiotic resistance in food animals and the subsequent transfer of these resistant organisms to humans have been reported.1,3,39 Resistance to antimicrobials has also been reported in aquaculture products or environments, as well as in finfish,40–42 with residues detected from several months 14 up to a few years posttreatment with antimicrobials. 15
Generally, the MARI for each site was slightly beyond the threshold of low risk to humans. This result implies that locations from which samples were collected have been exposed to high antibiotic contamination. This conclusion was unexpected in the recirculatory fish farm systems (Farm 1 and Farm 2), as both farms reported using an antibiotic-free fish meals. These results might, therefore, indicate an incoming water source to the fish farms, which is already contaminated with antibiotic-resistant bacteria. The MARI obtained from isolates from Kariega Estuary was higher than that recorded in both farms, which we expected as the estuary is exposed to contaminants from various sources. Water bodies have been reported to be most contaminated, with resistant bacteria from human and animal waste, industries and hospitals, farmland runoff, municipal sewage, and poorly treated sewage.43–46 These resistant bacteria contaminate receiving water bodies and, therefore, aquatic life, including fish.47,48 This fact is reflected in many reports that have revealed the presence of antibiotic-resistant Vibrio in final effluents of wastewater treatment plants.34,49–51 It is unfortunate that most of these pathogens can eventually be transferred through the food chain or skin abrasions to humans, causing illnesses that are difficult to treat with regular antibiotics.
Prevalence of antibiotic resistance genes
The resistant genes ampC, blaZ, tetA, tetM, dfr1, sul1, sul2, ermB, nptII, and strA were detected in two or more isolates, while the SXT integrase was detected in one isolate. Other studies have reported resistant genes in Vibrio species, which could be located in the bacterial chromosome or plasmid mediated.11,31 Yano et al. 10 also detected β-lactamase and tetracycline-resistant genes in Vibrio species isolated from cultured shrimps in Thailand. The inappropriate use of antimicrobials could lead to the emergence of resistant bacteria and genes. Resistance genes are neither a respecter of phylogeny nor geographical or ecological borders 52 ; therefore, antibiotic-resistant bacteria or genes in the incoming waters might have been sources of ARGs to the farms. These resistant bacteria harboring resistant genes may transfer these genes to susceptible bacteria, possibly through plasmids.
Conclusion
Our findings represent the first report on antibiotic-resistant vibrios with circulating ARGs from finfish aquaculture in South Africa. The findings of this study support the hypothesis that marine fish and their environs may serve as vehicles of antibiotic-resistant vibrios and resistance genes to humans through the food chain or through contact with water, and this is a public health concern. Continuous monitoring of the antimicrobial-resistant patterns is therefore necessary to determine the emergence and spread of resistance, particularly in controlled environments such as aquaculture. This calls for better implementation of quality control strategies to prevent further transmission of resistant bacteria to humans and ARGs to susceptible bacteria.
Footnotes
Acknowledgments
The authors are grateful to the South African Institute for Aquatic Biodiversity (SAIAB) and NRF for their financial support. They are also grateful to the directors of the fish farms for provision of samples for the study.
Disclosure Statement
No competing financial interests exist.