
Abstract
The inactivated mgrB gene and the mcr-1 gene are important mechanisms of colistin resistance in Klebsiella pneumoniae and they are threats to the clinical use of colistin. In this study, mcr-1 gene was cloned into K. pneumoniae strains (XH209 and KP10) and their derived strains (XH209 M and KP10 M), which showed high-level resistance to colistin. The acquisition of the mcr-1 gene led to colistin resistance in XH209 and KP10, but the addition of mcr-1 gene did not cause change of colistin minimum inhibitory concentrations in the XH209 M and KP10 M. In addition, the impact of mcr-1 gene on growth rate showed strain specific in K. pneumoniae. In conclusion, the mcr-1 gene does not cause the same level of colistin resistance as the inactivated mgrB gene in K. pneumoniae. The mcr-1 gene has no effect on colisitin resistance when it coexists with inactivated mgrB gene in K. pneumoniae.
Introduction
M
Materials and Methods
Bacterial strains
All the bacterial strains in this study were isolated from patients with bloodstream infection. The E. coli strain 20COE13 carried mcr-1 gene and was resistant to colistin. 9 The K. pneumoniae strains XH209 and KP10 were resistant to carbapenems. 10
Antimicrobial susceptibility testing
The colistin minimum inhibitory concentrations (MICs) were determined by broth microdilution according to the Clinical and Laboratory Standards Institute (CLSI) recommendation, and the results were interpreted in accordance with the European Committee on Antimicrobial Susceptibility Testing (EUCAST) breakpoints (www.eucast.org/clinical_breakpoints).
PCR amplification and sequencing
The colistin resistance-related genes in K. pneumoniae were amplified and sequenced, which included two-component systems phoP/phoQ, pmrA/pmrB, crrA/crrB, the mgrB gene and mcr-1 gene. The primers have been reported in previous studies.5,6,11 The presence of blaKPC-2 gene was also confirmed by PCR. 12 The nucleotide and deduced protein sequences were analyzed at the National Center for Biotechnology Information website (www.ncbi.nlm.nih.gov) using the Basic Local Alignment Search Tool program. We did multilocus sequence typing (MLST) with the primers and protocol specified in the online MLST database for K. pneumoniae.
DNA manipulations
The mcr-1 gene and part of its flanking sequence were amplified from 20COE13 using the following primers: F-5′- TTATCGCGTCGAGTCTGAAC-3′ and R-5′-AAGCTTGCTTGGCAACCATC-3′ (GenBank, accession number KY012274). The purified PCR products were cloned into the pCR2.1 vector using T4 DNA ligase (Invitrogen, Shanghai, China). Then, the recombinant plasmid pMCR was introduced into the competent E. coli DH5α for amplification. The plasmid pMCR was transformed into strain recipient bacterium through electroporation. The plasmid pCR2.1 was also introduced into the strains as blank controls. Transformants were selected on Müller-Hinton (MH) agar plates containing 50 mg/L kanamycin and confirmed by PCR amplification and sequencing.
Population analysis profiling
Population analysis profiling (PAP) was performed using the spread plate techniques to obtain the derived colistin-resistant isolates of the original strains.13–15 The XH209 and KP10 were cultured overnight and were diluted to a gradient of colony-forming units (CFUs), as 108, 107, 106, and 105 CFU/mL. A 50 μL volume of each dilution was spread on each MH agar plates with a gradient of colistin concentrations, which were set as 0, 1, 2, 4, 8, 16, 32, and 64 mg/L. Then they were cultured for 24 hr. The colonies found in 4 mg/L or higher concentrations were regarded as derived colistin-resistant isolates. The colistin MICs of the selected isolates were reassessed after serial daily subcultures on antibiotic-free medium for 3 days.13,14
Growth rate determination
The growth curves of bacteria were measured in MH broth at 37°C by taking OD600 measurements every 5 min using the Bioscreen C reader (Oy Growth Curves Ab Ltd., Finland) for 16 hr. Four independent cultures of all strains were grown overnight and were diluted 1:1,000 with the MH broth containing kanamycin. They were aliquoted into a flat-bottom 96-well plate in triplicate (0.2 mL/well). The growth rates were calculated using an R script (https://github.com/decarpen/growth-curves).
Result
The XH209 carried a blaIPM-4 gene and belonged to ST17, whereas the KP10 carried a blaKPC-2 gene and belonged to ST11. They both showed resistance to carbapenems and were sensitive to colistin (Table 1). The strain XH209 M and KP10 M were obtained in PAP, and they both showed a change of colistin MIC from 0.06 to 128 mg/L (Table 2). The genes related to colistin resistance were further identified by PCR analyses (two-component systems pmrA/pmrB, phoP/phoQ, crrA/crrB, and the mgrB gene). Inactivated mgrB gene was detected in the strain XH209 M and KP10 M. Compared with XH209, the XH209 M showed insertional inactivation of the mgrB gene. The insertion sequence IS26 belonged to the IS6 family and it is located at nucleotide 68 of the mgrB gene. Similarly, the KP10 M also owned an inactivated mgrB gene. The insertion sequence ISKpn14 belonged to the IS1 family and it is located at nucleotide 97 of the mgrB gene. The mcr-1 gene derived from E. coli strain 20COE13 also caused colistin resistance in K. pneumoniae. The XH209-pMCR showed a colistin MIC of 32 mg/L, whereas the colistin MIC of KP10-pMCR was 16 mg/L. But the strains with inactivated mgrB gene showed the same colistin MIC with or without the recombinant plasmid pMCR (Table 2).
MIC, minimum inhibitory concentration; N., not found.
N, not resistant to colistin.
Growth rates of the strains were determined by performing calculations on the growth curves. Compared with the strain XH209-pCR2.1, the XH209-pMCR showed a lower growth rate (p < 0.05). Similarly, XH209 M-pMCR showed a lower growth rate than XH209 M-pCR2.1 (p < 0.05). But, the growth rates of the strain KP10-pMCR was higher than KP10-pCR2.1 (p < 0.05). The growth rates of KP10 M-pCR2.1 and KP10 M-pMCR showed no significant difference (p > 0.05) (Fig. 1).
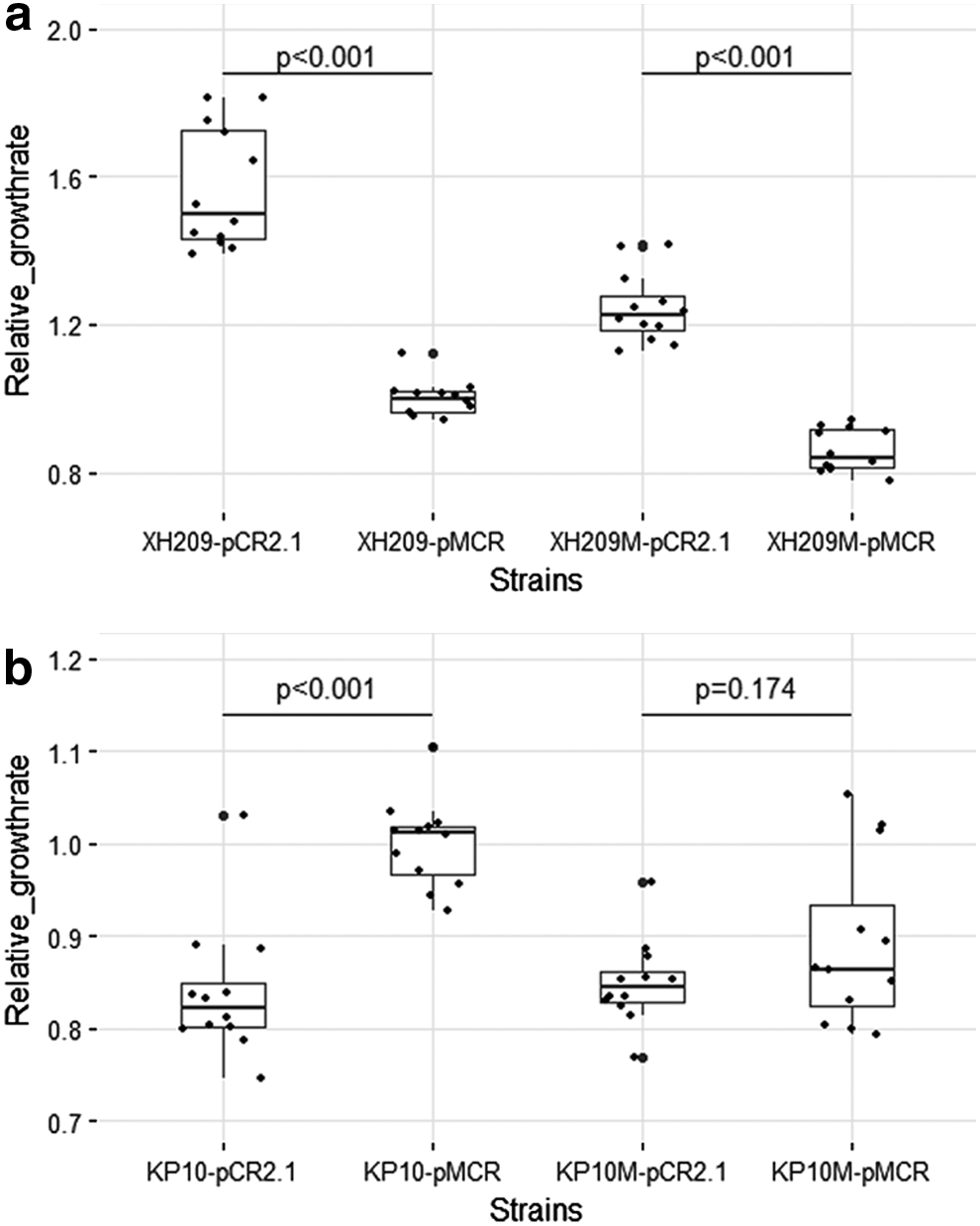
Relative growth rates of all the strains.
Discussion
As important mechanisms leading to colistin resistance in K. pneumoniae, mcr-1 gene and inactivated mgrB gene have different characteristics. They both contribute to the severe situation of colistin resistance worldwide. The mcr-1 gene causes resistance to colistin by PEtN modification with lipid A. Chromosomally, the mgrB gene plays an important role in K. pneumoniae's acquired resistance to colistin, especially in the carbapenem-resistant K. pneumoniae strains, for which there are very few therapeutic options.3,16
The K. pneumoniae XH209 and KP10 strains were both resistant to carbapenem. More importantly, the KP10 belonged to ST11, which was the key KPC-producing strain in China, and carried a blaKPC-2 gene, which was the most dominant carbapenemase gene type in K. pneumoniae from China. 17 The XH209 and KP10 developed high-level resistance to colistin in vitro immediately and rapidly. The inactivation of mgrB gene caused colistin resistance in XH209 M and KP10 M. The transformation of mcr-1 gene also caused colistin resistance, but the MICs of colistin was relatively lower than the XH209 M and KP10 M. The MIC results indicated that the mcr-1 gene and inactivated mgrB gene showed no synergistic effect in colistin MIC value in K. pneumoniae. The addition of mcr-1 gene did not cause change of colistin MICs in the XH209 M and KP10 M. When the mcr-1 gene and the inactivated mgrB gene coexisted in K. pneumoniae, it showed the same colistin MIC with the strain owing inactivated mgrB gene alone. This illustrated the leading role of inactivated mgrB gene in lipid A modification when it coexisted with the mcr-1 gene. The mcr-1 gene and the inactivated mgrB gene both cause colistin resistance by modification of the lipid A. The mcr-1 leads to the addition of PEtN to lipid A, whereas the inactivated mgrB gene ultimately results in modification of lipid A by the addition of L-Ara4 N and PEtN.3,5 Obviously, no synergistic effect was found between mcr-1 and inactivated mgrB gene in K. pneumoniae, and the mcr-1 gene did not cause the same level of colistin resistance as the inactivated mgrB gene in K. pneumoniae. The function of mcr-1 gene might be covered up by the inactivated mgrB gene.
The growth rates of the strains were measured as a proxy for fitness, so as to evaluate the impact of mcr-1 gene in K. pneumoniae. The growth rates of XH209-pMCR and XH209 M-pMCR were significantly reduced compared with the strains without the mcr-1 gene. But in the KP10 series, mcr-1 gene did not lead to any fitness cost. The KP10-pMCR even showed a higher growth rate than the strain KP10-pCR2.1. Taken together, the impact of mcr-1 gene on growth rate showed strain specific in K. pneumoniae. Fitness cost of mcr-1 gene in K. pneumoniae was not stabilized. In different strains it would cause different results. The sequence types might have an effect on the fitness cost, but further studies are needed.
The mcr-1 gene is an important colistin resistance mechanism in Enterobacteriaceae. It is the first reported plasmid-mediated mechanism and it owns the ability of horizontal gene transfer. But in K. pneumoniae, its chromosomally mediated colistin resistance mechanisms, especially the mgrB inactivation, should not be ignored. They could cause resistance to colistin without exogenous genes, and more importantly, the mgrB inactivation has been regarded as an important mechanism of colistin resistance in the carbapenem-resistant K. pneumoniae strains. While the carbapenem-resistant K. pneumoniae carrying mcr-1 gene is rare and only isolated case reports have described them. In conclusion, no synergistic effect was found between mcr-1 and inactivated mgrB gene in K. pneumoniae. The mcr-1 gene played a role in lipid A modification, but it does not confer the same level of colistin resistance as the inactivated mgrB gene in K. pneumoniae. When the mcr-1 gene coexists with the inactivated mgrB gene in K. pneumoniae, it has no effect on colisitin resistance. Further investigations are necessary to understand the specific mechanisms modulated in bacteria carrying mcr-1 and inactivated mgrB gene.
Footnotes
Acknowledgments
This work was supported by the Medical and Health Research Project (2017KY224), and the Natural Science Young Foundation of Zhejiang Province, China (LQ17H190006).
Disclosure Statement
No competing financial interests exist.