
Abstract
Although vanA carrying Enterococcus faecium human clinical isolates have been rarely found in Hungary before 2012, they have been detected in continuously increasing numbers since then. To identify factors associated with their dissemination, we investigated the clonal relatedness and plasmids of 30 vanA carrying E. faecium isolates originating from different Hungarian healthcare institutions from 2012 to 2014. Molecular typing of the isolates (n = 30) was performed with pulsed-field gel electrophoresis (PFGE), multilocus sequence typing, Tn1546 polymerase chain reaction mapping, plasmid restriction fragment length polymorphism analysis, and sequencing. A single Tn1546 variant was detected in all of the isolates. It harbored IS1251 in the vanS-vanH intergenic region, had an entire deletion of the transposase gene and a partial deletion of the resolvase gene, and was located on a pRUM-like plasmid. Based on PFGE, the isolates could be grouped into 13 pulsotypes. Representative strains of these pulsotypes belonged to ST17, ST18, ST80, ST117, and ST203, which are known to be part of the hospital-adapted clades. The increase in the number of vanA carrying E. faecium clinical isolates in Hungary could be explained by the dissemination of pRUM-like vancomycin resistance plasmids in hospital-adapted clonal lineages.
Introduction
S
Preliminary studies performed at the National Center for Epidemiology indicated that 76.9% (n = 268) of the vancomycin-resistant E. faecium isolates, which were sent to the Reference Laboratory between 2012 and 2013, carried the vanA gene (B. Berta, 2013, personal communication). To identify the underlying factors associated with the change in resistance rates, we studied clonal relatedness and structure of the vanA operon and of the carrying plasmids of E. faecium strains isolated in Hungary between 2012 and 2014.
Materials and Methods
Antimicrobial susceptibility testing was performed in clinical microbiology laboratories of participating Hungarian healthcare institutions. Presumably, VRE isolates were submitted for confirmation and typing to the Reference Laboratory of the National Center for Epidemiology (renamed to National Public Health Institute in 2017). The identification of the isolates to species level and the detection of vanA, vanB, and vanC genes were performed with polymerase chain reaction (PCR) using primers based on the study by Dutka-Malen et al. 4 For molecular typing, pulsed-field gel electrophoresis (PFGE) was carried out as described elsewhere. 5 Pulsotypes and subtypes were defined as isolates with >80% and >95% similarity in banding patterns, respectively.
Thirty, nonduplicate vanA carrying E. faecium isolates were selected for further study. The selection was done as to be representative for the participating healthcare institutions, the time period, and the distribution of pulsotypes. Multilocus sequence typing (MLST) was performed for one isolate of each pulsotype and subtype. 6 Bayesian Analysis of Population Structure (BAPS) groups were defined according to study by Willems et al. 7 To detect Tn1546 structural variants, PCR mapping was carried out using primers based on the study by Arthur et al. 8 The first VanA-type E. faecium isolate was selected for Sanger sequencing of Tn1546. Based on the sequencing results, additional primers (p20–p23, Table 1) were designed to test for the integration of the IS1251-like element in the vanS-vanH region for all isolates.
Three isolates from different years, regions, and sequence types were selected for plasmid sequencing. Isolates were grown in a brain-heart infusion broth containing 10 mg/L vancomycin, and plasmid DNA was extracted with alkaline lysis method and purified with phenol–chloroform extraction. 9 Sequencing was performed on Illumina MiSeq with 2 × 250 bp paired ends. The quality of the data was assessed with FastQC 0.11.5 (Braham Institute, Cambridge, United Kingdom). PlasmidSPAdes 3.8.1 was used for de novo assembly. 10 De Bruijn graphs were visualized with Bandage 0.8.0. 11 Paths corresponding to plasmids were chosen according to depth and comparison to plasmid sequences in GenBank. The selection of the paths (order of the contigs) was verified with PCR (primers vp1–vp18b; Table 1) and restriction digestion of plasmid DNA with Bsu15I (ClaI).
Plasmids were annotated with GeneMark (Georgia Institute of Technology, Atlanta, GA), and their sequences were uploaded to GenBank (accession Nos. KY595962–KY595970). 12 Plasmid maps were created with SnapGeneViewer (GSL Biotech, LLC, Chicago, IL). Digestion of plasmid DNA with Bsu15I (ClaI) was performed for all isolates (plasmid restriction fragment length polymorphism [pRFLP]) and the results were evaluated for bands between 1,000 and 10,000 bp. Those isolates were considered to belong to pRFLP type A1, for which the characteristic restriction fragment pattern of pBUD102/pMIS102 (5,144, 4,717, 4,474, 3,462, 3,312, 2,361, 2,009, and 1,978 bp) was shown. Isolates with one to two band differences were designated type A2–A4. Isolates that differed in more than two bands were defined as type B–D. The presence of the pRUM replicon [rep(17)-F and rep(17)-R] and the axe-txe genes (Axe-Txe-F and Axe-Txe-R) were detected with PCR using primers based on the study by Freitas et al. 13
Results
Between 2012 and 2014, vanA carrying E. faecium was confirmed for 68% of the patients (n = 437), from whom presumably VRE isolates were submitted to the Reference Laboratory (Table 2). According to the results of PFGE, the vanA carrying E. faecium isolates were grouped into 77 pulsotypes, of which the most frequent one (designated ENTCO-002/a/c) enclosed 36% (n = 332) of the isolates and it could be subdivided into several subtypes (designated ENTCO-002, ENTCO-002a, and ENTCO-002c). Other pulsotypes represented a subtype at the same time as the similarity level was found to be >95% within those pulsotypes.
N/A, not applicable; VRE, vancomycin-resistant Enterococcus spp.
The results of MLST, Tn1546 typing and pRFLP are summarized in Table 3. According to the results of MLST, the isolates belonged to either BAPS2.1a (ST80 n = 1, ST117 n = 7, and ST203 n = 4) or BAPS3.3a (ST17 n = 2 and ST18 n = 2). The BAPS2.1a and BAPS3.3a isolates belonged to pulsotypes that included altogether 73% (22/30) and 27% (8/30) of the isolates, respectively.
Type F3 designated by Freitas et al. 13
MLST, multilocus sequence typing; NA, not analyzed; PFGE, pulsed-field gel electrophoresis; pRFLP, plasmid restriction fragment length polymorphism.
Typing of Tn1546 revealed that 26 isolates shared an identical variant with an ∼1,500 bp increase of p11p12 and a lack of the first 4 segments (p1p2-p7p8) (Table 4). Sanger sequencing showed integration of a copy of IS1251-like element in the vanS-vanH intergenic region (corresponding to additional DNA in p11p12), and total deletion of orf1 (transposase) and partial deletion of orf2 (resolvase), consistent with the absence of fragments p1p2-p7p8. This structure corresponded to “type F3” designated by Freitas et al. and detected in neighboring country (Serbia) in 2005. 13 Four isolates showed additional alterations, and their transposons were designated here subtypes F3.a-c. With primers p20–p23, the integration of the IS1251-like element in the vanS-vanH intergenic region was confirmed in all isolates.
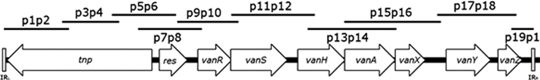

Size of the PCR product and the region of Tn1546 amplified was indicated for strain BM4147. 8
Size of the PCR product and the region of Tn1546 amplified was indicated for the isolate for which Sanger sequencing of Tn1546 was performed.
−, No PCR product was detected.
+, The size of the PCR product matched the indicated size.
++, The size of the PCR product was larger than the indicated size.
PCR, polymerase chain reaction.
The results of sequencing and assembly of plasmids are summarized in Table 5. The sequence quality (per base, per tile, and per sequence) of the short reads was good. The report indicated overrepresentation of certain sequences (warnings for “per sequence GC content,” “sequence duplication level,” and “overrepresented sequences”). Overrepresented sequences totaled upto <0.15% of total sequences and by BLAST search, they gave match with different E. faecium plasmid sequences (replication module protein, insertion sequence, hypothetical protein and noncoding sequence). The overrepresentation of certain sequences could be explained with the large variations in the copy number of plasmids.
For isolate 87, nodes with depth ≥100 × were given.
Match between in vitro and in silico results is indicated with (+). Discrepancy between in vitro and in silico results is indicated with (−).
The name (accession number) and statistics (“c” coverage, “id” identity) of the plasmid with the highest maximum score in BLAST search are indicated.
GC, guanine-cytosine content.
After removing dead ends and nodes with low depth (<50 × for isolate 1 and 81; <100 × for isolate 87) from the assembly graph, several nodes (contigs) interconnected through a central node (corresponding to IS1216) were visible for each of the isolates (Figs. 1–3).
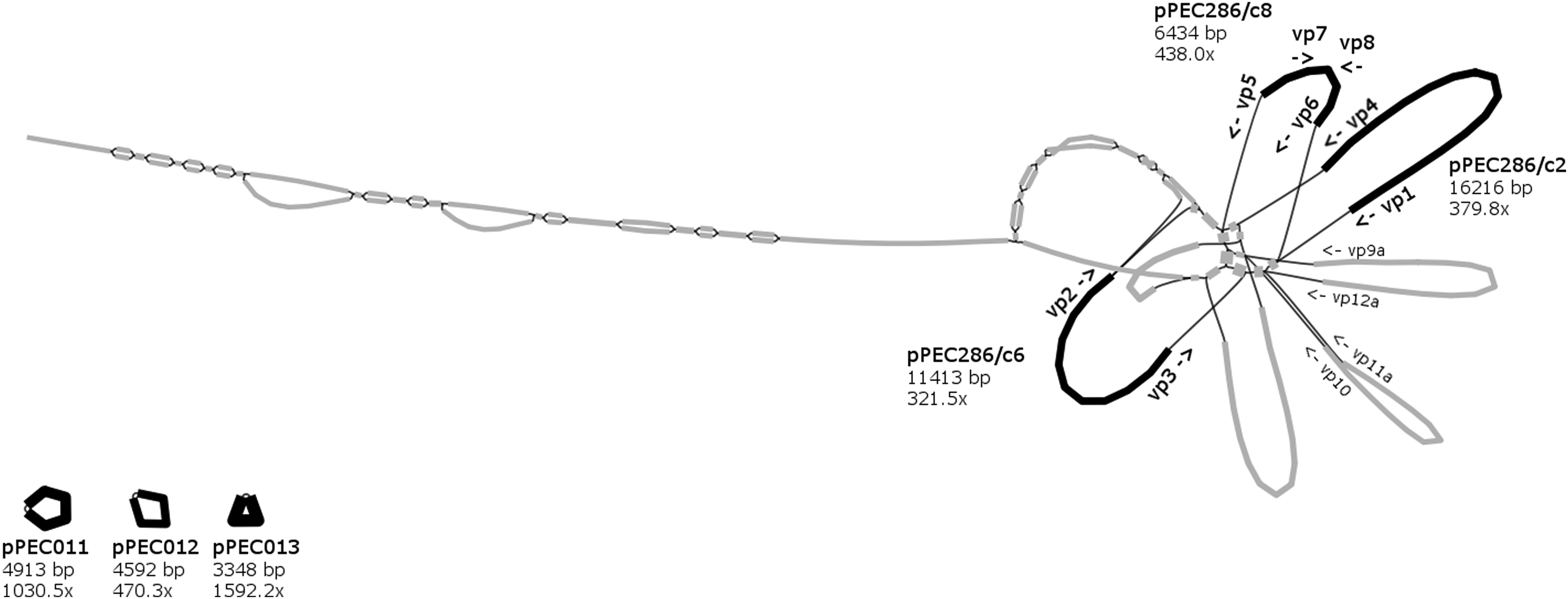
Assembly graph of isolate 1 showing ≥50 × depth contigs and binding sites of scaffolding primers. Nodes are represented by black thick lines (contigs assigned to plasmids) and gray thick lines (contigs not assigned to plasmids). Edges indicate overlaps between node sequences and are shown with black thin lines.

Assembly graph of isolate 81 showing ≥50 × depth contigs and binding sites of scaffolding primers. Nodes are represented by black thick lines (contigs assigned to plasmids) and gray thick lines (contigs not assigned to plasmids). Edges indicate overlaps between node sequences and are shown with black thin lines.
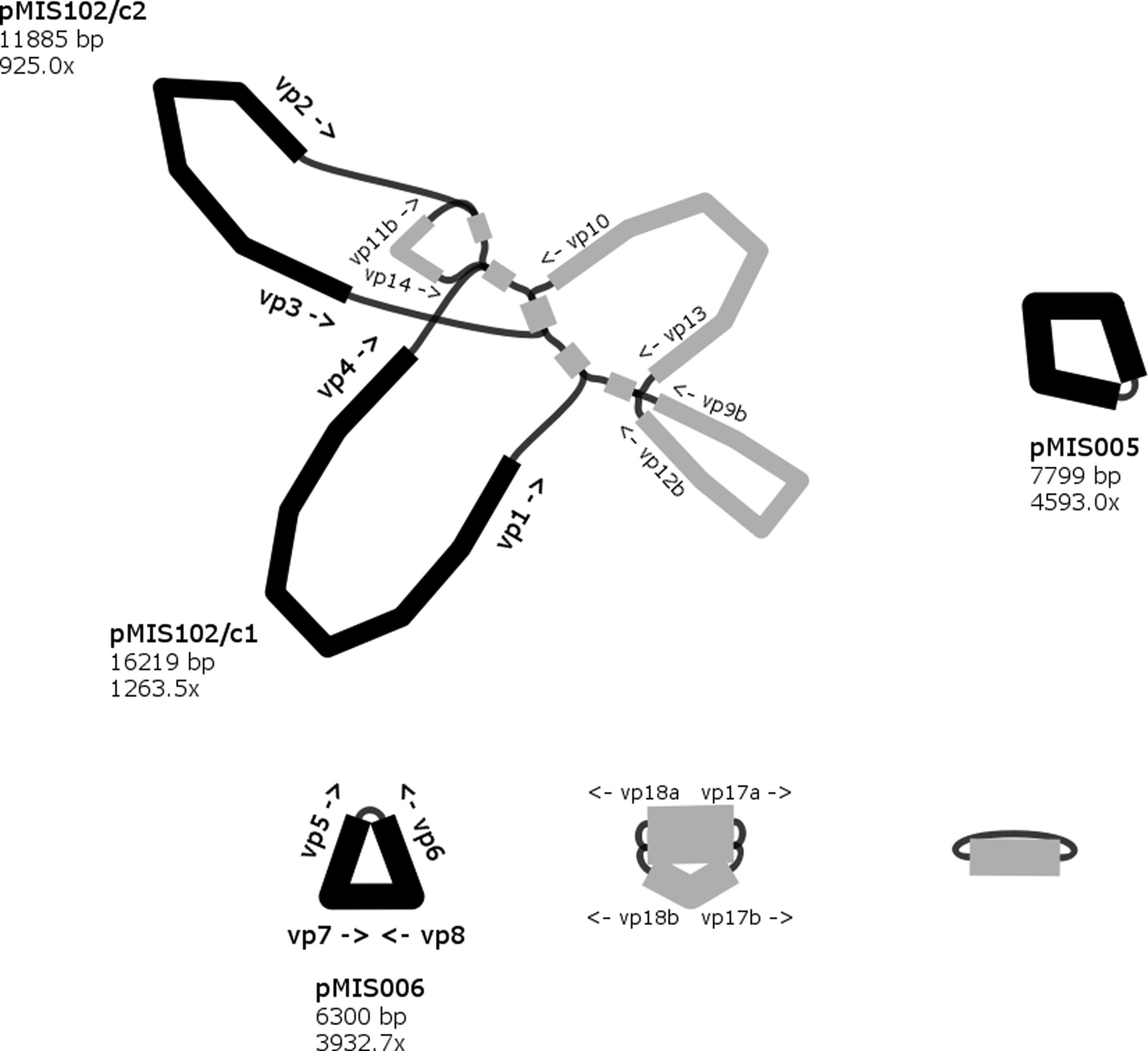
Assembly graph of isolate 87 showing ≥100 × depth contigs and binding sites of scaffolding primers. Nodes are represented by black thick lines (contigs assigned to plasmids) and gray thick lines (contigs not assigned to plasmids). Edges indicate overlaps between node sequences and are shown with black thin lines.
Altogether, 14 paths predicted to be distinct plasmids were selected. Scaffolding primers vp1–vp18b were designed to bind to the end of contigs (Figs. 1–3). For nine paths, the in vitro results of the scaffolding PCR matched the in silico predicted one (Table 5). The contigs of these closed plasmids are shown with black thick lines in Figs. 1–3. Five paths could not be circularized and their contigs (gray thick lines on Figs. 1–3) were not considered for further analysis. For all closed plasmids, the observed size of restriction fragments (Fig. 4) matched the in silico predicted one. This way the sequence of four, two, and three plasmids could be verified for isolate 1, 81, and 87, respectively (Table 5).
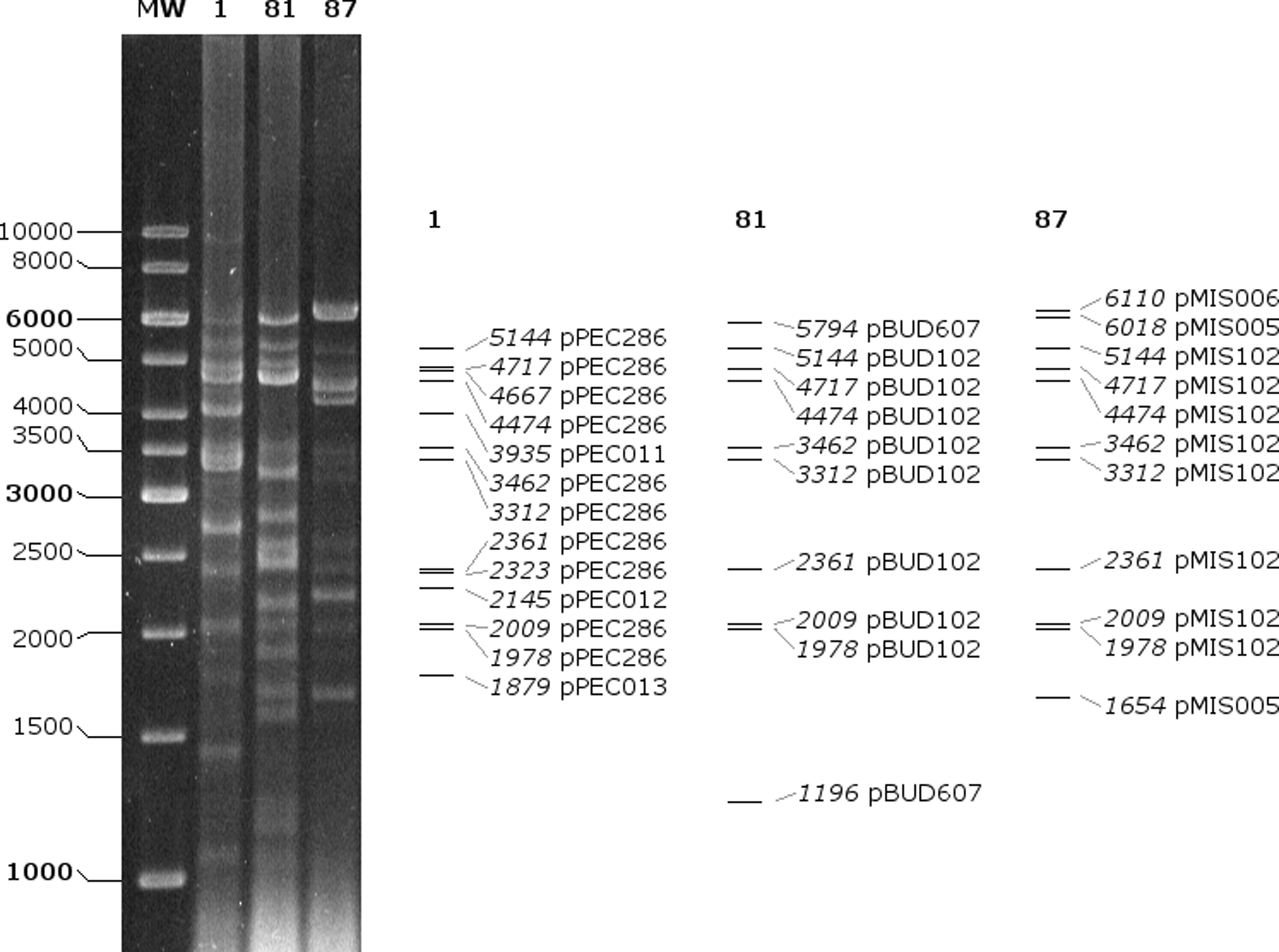
In vitro (left) and in silico (right) digestion of plasmids with Bsu15I (ClaI). Calculated fragment sizes (bp) and parent plasmids are indicated.
For the 9 plasmids, altogether 109 open reading frames and 61 unique proteins were predicted (Table 6). Maps of the vancomycin resistance plasmids pPEC286 and pBUD102 are shown on Figs. 5 and 6. Plasmids pPEC286, pBUD102, and pMIS102 harbored the Tn1546 variant-type F3 and shared 29,134 bp with each other (identity: 100%). The best match retrieved with BLAST search for this shared sequence was unnamed plasmid 3 of strain ATCC700221, which is a vancomycin resistance plasmid with a pRUM backbone. In addition, pPEC286 contained the complete sequence of the bacteriocin encoding plasmid pB82. The sequence of pB82 was also identified in the other two isolates on plasmids pBUD607 and pMIS006. Plasmids pPEC011, pPEC012, pPEC013, and pMIS005 encoded proteins with unknown functions and belonged to the Rep_3 family.
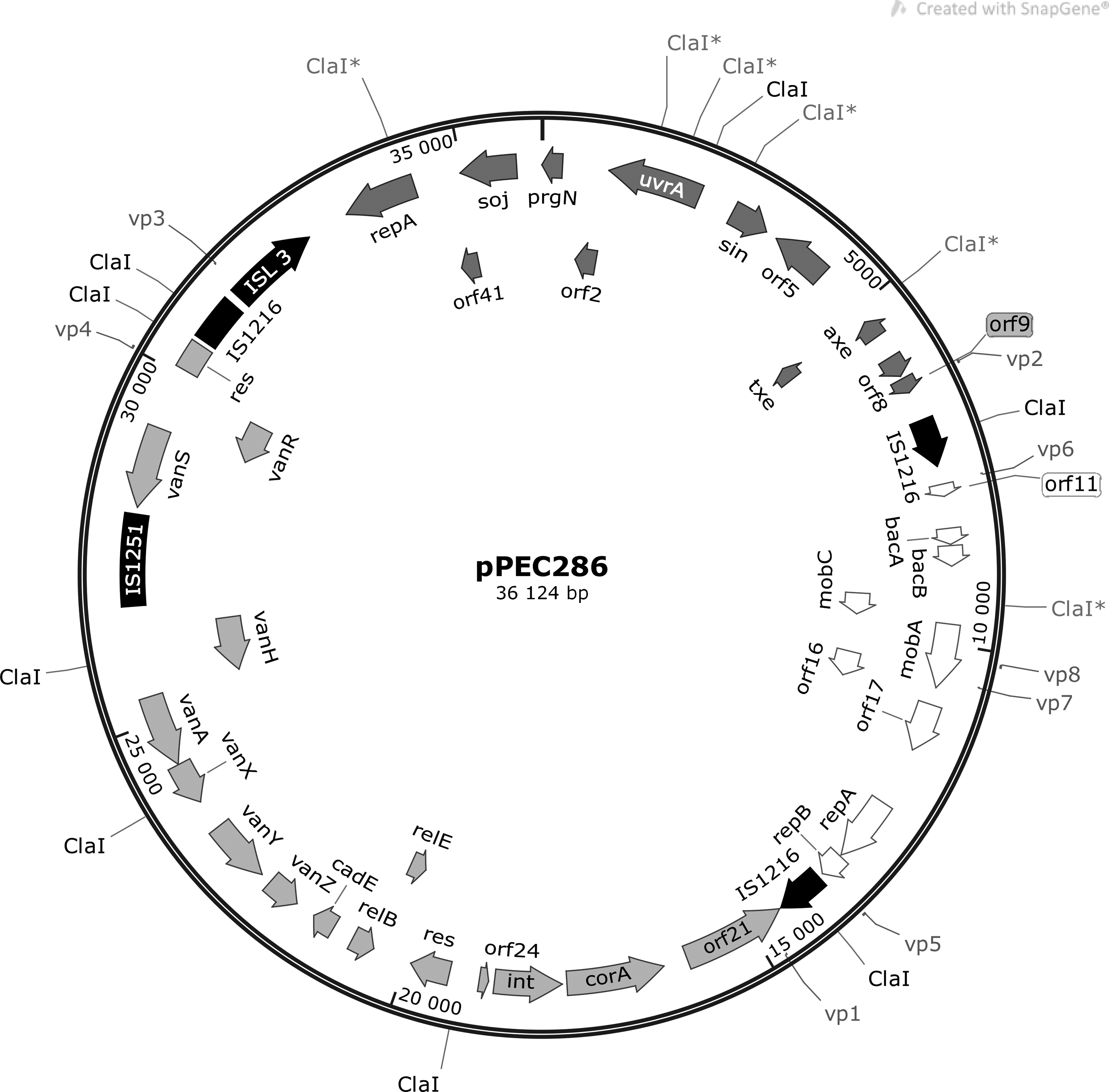
Structure of pPEC286. (IS elements—black; genes from pRUM—dark gray; genes from p3 of ATCC700221—light gray; genes from pB82—white; restriction sites and binding sites of scaffolding primers vp1–vp8 are also indicated.) IS, insertion sequence.
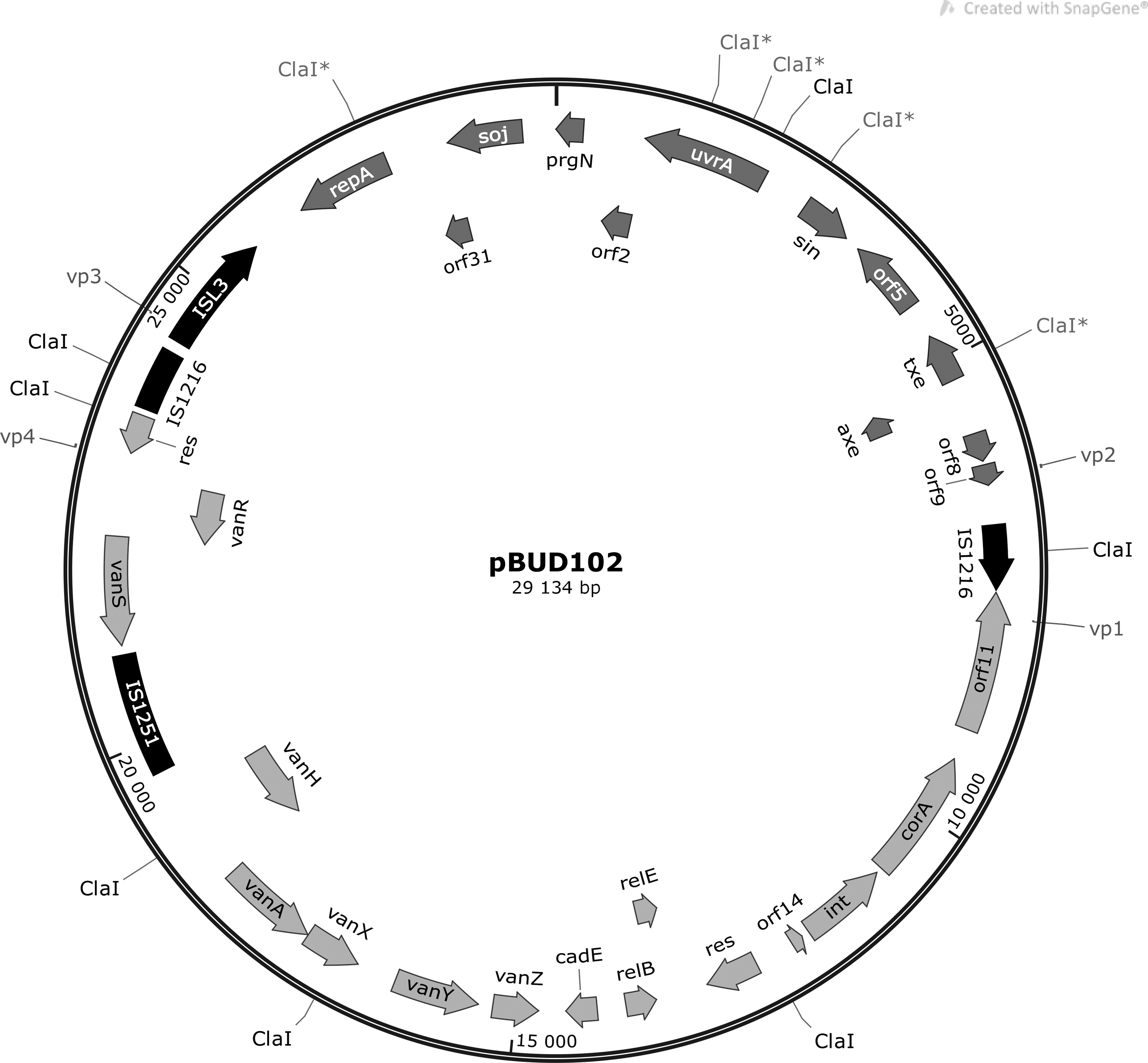
Structure of pBUD102. (IS elements—black; genes from pRUM—dark gray; genes from p3 of ATCC700221—light gray; restriction sites and binding sites of scaffolding primers vp1–vp8 are also indicated.)
c, Complementary strand; TA, toxin-antitoxin; UV, ultraviolet.
According to the results of pRFLP, the characteristic restriction fragment pattern of pBUD102/pMIS102 was shown for 13 isolates (type A1, Figs. 7 and 8). For 14 isolates, additional variations of type A1 were detected (type A2–A4). Types A2, A2a, and A2b had alteration of the 3,462 and 3,312 bp fragments. This could correspond to alterations in the repA restriction site. In type A2a, the 4,474 bp fragment carrying the vanH, vanS, and vanR genes seemed to be enlarged, which corresponds to the increment in p9p10 that was observed in Tn1546 PCR mapping (type F3a). In type A2b, an increment of the 4,717 bp fragment (relBE, vanZYX) was detected, which is in line with the enlargement of p15p16 (Tn1546 type F3.c). For type A3 and A4, the lack of 4,717 and 5,144 bp fragments was characteristic, respectively. Three isolates had unique pRFLP patterns (type B–D).
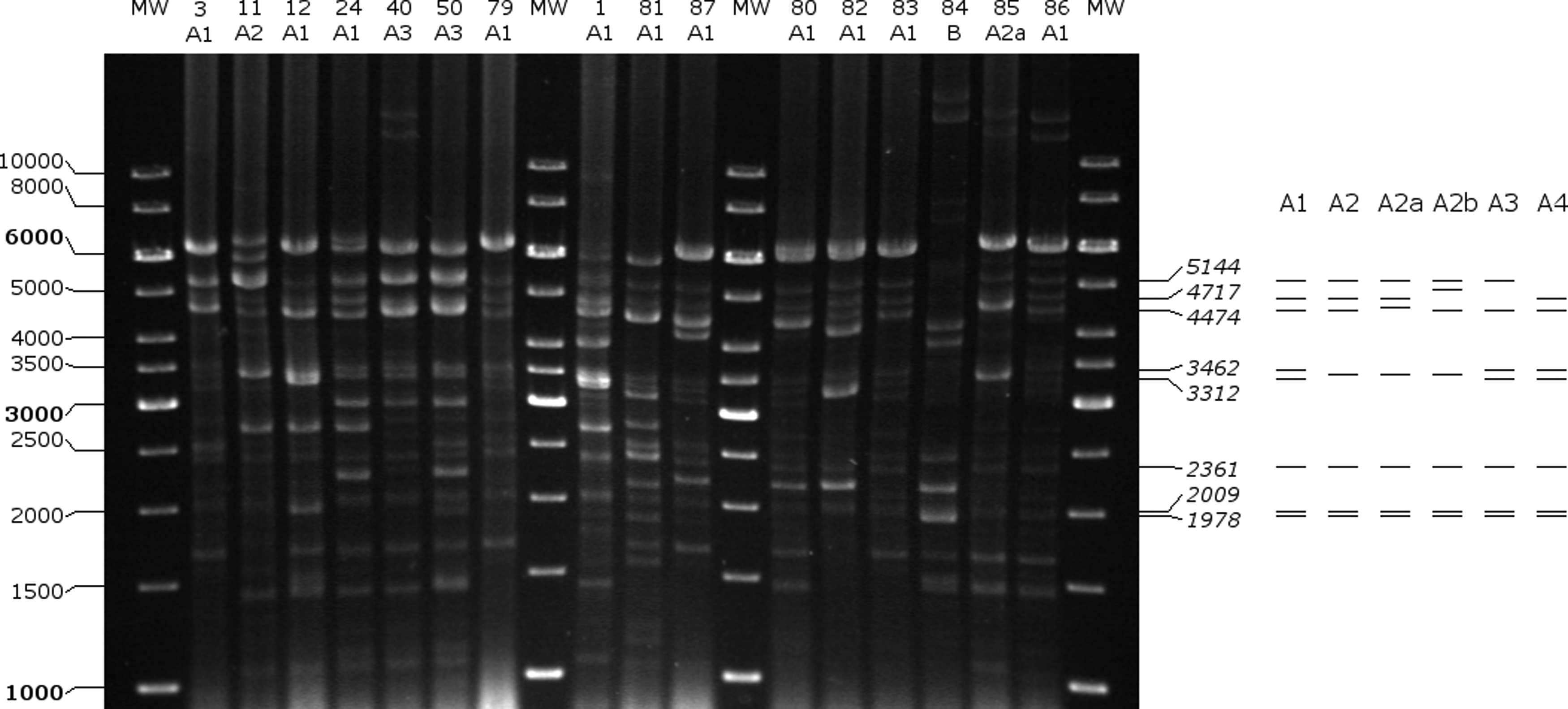
Restriction fragment analysis of plasmids with Bsu15I (ClaI) and pRFLP types of isolates 1–87. A schematic drawing of pRFLP types A1–A4 is shown on the right. pRFLP, plasmid restriction fragment length polymorphism.
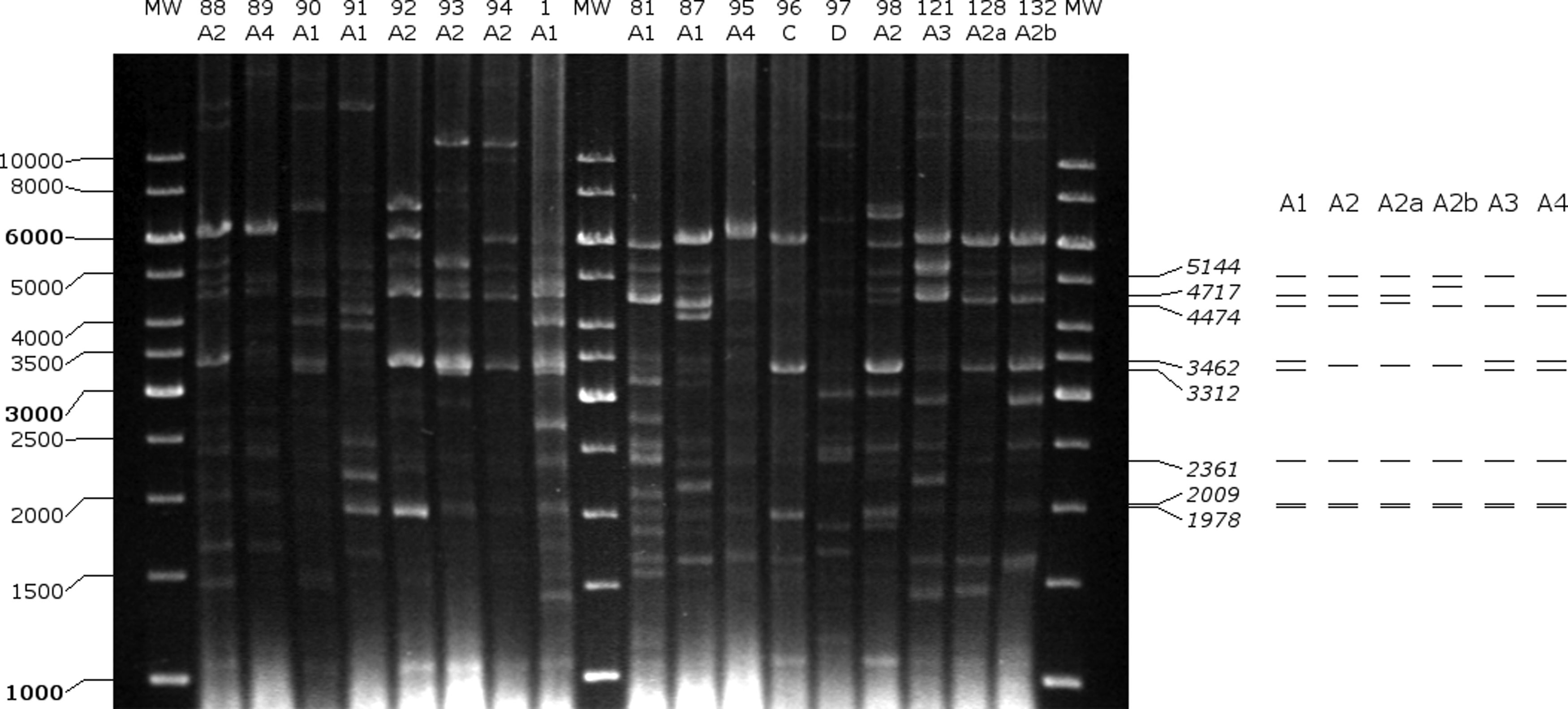
Restriction fragment analysis of plasmids with Bsu15I (ClaI) and pRFLP types of isolates 88–132. A schematic drawing of pRFLP types A1–A4 is shown on the right.
With primers rep(17)-F–rep(17)-R and Axe-Txe-F–Axe-Txe-R, the presence of the pRUM replicon and the Axe-Txe toxin-antitoxin system were shown for all isolates.
Discussion
To our knowledge, this was the first study to analyze the Tn1546 structure and vanA carrying plasmids of E. faecium human clinical isolates in Hungary. The results indicate that the widespread dissemination of VanA-type E. faecium in our country after 2012 can be attributed to the spread of pRUM-like plasmids in hospital-adapted clonal lineages (BAPS 2.1a and BAPS3.3a). These plasmids carried a Tn1546 variant “F” with insertion of IS1251 in the vanS-vanH intergenic region.
The pRUM plasmids belong to the RepA_N family, have a narrow host range, and show differences in their structure (replicon sequence, Tn1546 structure, and absence of Axe-Txe toxin-antitoxin system). 13 The structure of the vancomycin resistance plasmids (pPEC286, pBUD102, and pMIS102) identified in this study highly resembled the pRUM variant, which was originally widespread in the United States and was later found worldwide, including a neighboring country, Serbia.13,14
Although the aim of the plasmid next-generation sequencing was to identify the vancomycin resistance plasmids, additional small-sized Rep_3 plasmids (pBUD607, pMIS005, pMIS006, pPEC011, pPEC012, and pPEC013) could be assembled in each of the isolates. However plasmids belonging to Rep_3 family were shown to be prevalent in hospital-adapted E. faecium clonal lineages, their exact contribution to the dissemination has not yet been fully elucidated. 15 Besides, other plasmids might have remained unassembled, considering that several contigs could not be assigned to plasmids (Figs. 1–3) and multiple bands in pRFLP could not be associated with the assembled plasmids.
Although the plasmids of only three isolates were sequenced, we presume that the majority of the isolates also carried pRUM-like vancomycin resistance plasmids. This is supported by the following findings: (1) the replicon and the Axe-Txe toxin-antitoxin system of pRUM were shown with PCR for all isolates. (2) All isolates had the Tn1546 variant with insertion of IS1251 in the vanS-vanH intergenic region, and this variant was mostly identified on pRUM-like plasmids. 13 (3) The exact pRFLP pattern of pBUD102/pMIS102 (type A1, 13/30) or a pattern with slight differences (type A2–A4, 14/30) could be recognized altogether in 90% of the isolates. In the remaining isolates, the variations in the patterns could be explained by alteration through mutation or recombination; integration into the chromosome; or presence of different vancomycin resistance plasmids.
Several clonal lineages are known to contribute to the spread of VanA-type E. faecium isolates worldwide. 13 In Hungary, the BAPS3.3a group was detected in sporadic human isolates between 2004–2005. 2 In this study, most of the isolates belonged to BAPS2.1a and only a smaller portion clustered within BAPS3.3a. However, MLST was performed for a limited number of isolates; we presume that this distribution should highly resemble the sequence type and BAPS group distribution of all isolates, considering that (1) PFGE, MLST, and whole-genome sequencing (WGS) were shown to be highly concordant and PFGE at 88% similarity level supported the WGS subgrouping, 16 and (2) that in our study, the selection criteria for MLST were based on >95% similarity in PFGE banding patterns.
The distribution observed in this study might reflect the global tendency, which shows a shift from BAPS3.3 (predominant in 1990s) to BAPS2.1 (predominant after 2005). 7 The fact that the pRUM-like plasmids were identified in two separate BAPS groups indicates that not only clonal expansion but also horizontal transfer of plasmids was involved in the dissemination.
Although pRUM-like plasmids were identified in many countries since 1992, and in a neighboring country (Serbia) in 2005, their presence in Hungary was first indicated by this study.2,13 We suggest that these plasmids could have been imported to Hungary from abroad. Considering the difficulties in establishing phylogenetic relationships among plasmids, the exact source and time point of the importation could not be determined.
Several factors were detected on these plasmids that could have facilitated the long-term stability of the vanA operon in the E. faecium population in Hungary. The Axe-Txe addiction system present on the pRUM backbone ensures stable maintenance. 14 Bacteriocin 43, encoded on pPEC286, pBUD607, and pMIS102, has a broad spectrum of activity. The bacteriocin 43 coding genes were shown to be co-mobilized with vancomycin resistance plasmids, and it might play a role in colonization and invasion.17,18
Besides the aforementioned microbial features, environmental factors might influence the dissemination of VanA-type E. faecium in Hungary. Several studies showed that the incidence of Clostridium difficile infections (CDI) and VRE colonization are associated, and that oral vancomycin and oral metronidazole given for the treatment of CDI promote the overgrowth of VRE in the gut.19–21 In Hungary, the CDI incidence rate increased considerably recently (2008: 2/10,000 patient days; 2011–2012: 12.3/10,000 patient days).22,23 The increment was largely attributed to the spread of hypervirulent 027 PCR ribotype. 24 The unfavorable change in the epidemiology of C. difficile might contribute to the spread of VRE in our country.
The worldwide success of pRUM-like vancomycin resistance plasmids suggests that vancomycin resistance will continue to circulate among E. faecium strains in Hungary. Therefore, effective measures to prevent or dismiss colonization, or preclude the progression to infection should be enforced.
Footnotes
Acknowledgment
We thank P. Courvalin (Institut Pasteur, Paris, France) for providing strain BM4147.
Disclosure Statement
No competing financial interests exist.