
Abstract
Rifampicin is one of the major drugs used on its own and also in combination to treat numerous infections sustained by methicillin-resistant Staphylococcus aureus (MRSA). In Italy, rifampicin resistance (RIF-R) is increasing in multidrug-resistant-MRSA isolates (16.4%), with respect to Europe (5.7%). In our study, the relationship between clones, rpoB mutations, and susceptibility profiles in 50 RIF-R MRSA isolated from hospitalized patients was evaluated. Antimicrobial susceptibility testing was performed by the broth microdilution method. Isolates were typed by MLST/SCCmec/spa-typing. The rpoB gene was analyzed by PCR and sequence analysis. RIF-R isolates were 60% heterogeneous vancomycin-intermediate S. aureus (hVISA) and 22% daptomycin nonsusceptible and belonged to the major MRSA clones: ST228-SCCmec I (44%), ST8-SCCmec IV (18%), ST239-SCCmec III (16%), ST5-SCCmec II (14%), and ST22-SCCmec IVh (4%). Thirteen diverse RpoB amino acid substitutions were identified. Half of the strains harbored the H481N substitution, conferring low-level resistance. Different single mutations at the equivalent locus (H481D; H481Y) or in other loci, and multiple mutations conferred high-level resistance. In conclusion, this study investigated the nature of RIF-R in Italy among RIF-R-MRSA strains, finding a prevalence of ST228, strongly associated with reduced susceptibility to glycopeptides (hVISA). The spread of RIF-R strains in clinical settings represents a serious threat, due to their complex resistance nature even to new anti-Gram-positive drugs, making these infections particularly difficult to treat.
Introduction
S
In recent years, the first choice therapy of glycopeptides has been hampered by the occurrence of strains showing different nonsusceptibility mechanisms such as heterogeneous vancomycin-intermediate S. aureus (hVISA) and/or VISA and daptomycin-nonsusceptible (DNS) phenotypes, underlying the need for new therapeutic options. 3 Dalbavancin, telavancin, and oritavancin antibiotics, together with fifth generation cephalosporins, have entered the market in the last 5–10 years, offering new strategic possibilities for the treatment of staphylococcal infections. 4
During a 3-month prevalence surveillance study on S. aureus performed in Italy in 2012, 5 an increasing rifampicin resistance (RIF-R) trait in MRSA isolates (16.4%) was found. This figure was considered worrisome if compared with the European trend (5.7%), as reported by the annual European Centre for Disease Prevention and Control (ECDC) surveillance report. 6
Rifampicin is one of the drugs used in combination 4 and mechanisms of resistance are almost mutational, accumulating in an extremely conserved region of the RNA polymerase beta-subunit, called “rifampicin resistance-determining region” (RRDR), spanning approximately from amino acid (aa) 462 to 550. 7 In particular, in this sequence, two clusters are closely associated with RIF-R: Cluster I (aa 462–488) and Cluster II (aa 515–530). 8
In addition, some authors demonstrated that some mutations in rpoB are associated with the alteration of vancomycin and daptomycin susceptibility giving a VISA and DNS phenotype. In particular, the H481Y mutation in the Mu50 strain provided a great contribution for the acquisition of the complete VISA phenotype with a mechanism far from being completely elucidated.3,9–11
The purpose of this study was to determine the rpoB gene mutations conferring RIF-R in MRSA strains isolated in Italy and their correlation with the main circulating clones.
Materials and Methods
The study sample consisted of 50 RIF-R strains isolated from hospitalized patients with documented bloodstream infections (BSIs), low-respiratory tract infections (LRTIs), and skin and soft-tissue infections (SSTIs), selected from a large collection of 640 MRSA strains collected in an Italian national survey during 2012. 5
The 50 strains were not geographically and epidemiologically related, belonging to 21 hospitals in 15 different cities.
Antimicrobial susceptibility tests
All strains had already been identified as previously reported. 5
Minimum inhibitory concentration (MIC) testing for the main anti-Gram positive agents: teicoplanin (Aventis, West Malling, United Kingdom), vancomycin (Sigma Chemical, St. Louis, MO), daptomycin (Novartis, Basel, Switzerland), tigecycline (Pfizer, Inc., New York, NY), linezolid (Pfizer, Inc.), ceftobiprole (Basilea Pharmaceutica Ltd., Basel, Switzerland), dalbavancin (Allergan Pharmaceuticals International Ltd., Dublin, Ireland) and rifampicin (Sigma-Aldrich, Milan, Italy), was carried out by the broth microdilution method, following standard criteria. 12
The antimicrobial susceptibility profiles of the MRSA isolates were evaluated according to EUCAST guidelines, and vancomycin-intermediate isolates were defined according to Clinical and Laboratory Standard Institute (CLSI).12,13
Tolerance, defined as a minimum bactericidal concentration (MBC)/MIC ratio ≥32 or ≥16 when the MBC was greater than or equal to the breakpoint for resistance, was evaluated only for dalbavancin not-susceptible strains.14,15
Detection of VISA/hVISA phenotype
For the screening of VISA and hVISA phenotypes the MIC Test Strip glycopeptide resistance detection (GRD) was used, comprising a double-sided predefined gradient of vancomycin (VA) and teicoplanin (TEC), following the instructions provided by the manufacturer (Liofilchem®, Italy). Interpretation criteria: hVISA was defined as GRD either VA or TEC ≥8 mg/L, with VA MIC <4 mg/L; VISA was defined as GRD either VA or TEC ≥8 mg/L, with VA MIC ≥4 mg/L. The results obtained were confirmed by using the population analysis profile/area under the curve (PAP/AUC) as a standard method, as already published. 16
Molecular characterization
All isolates were molecularly characterized for SCCmec typing, Panton-Valentine leukocidin and Arginine catabolic mobile element locus, MLST, spa-type, and mutations responsible for linezolid resistance, as previously described.17–21
Analysis of rpoB gene mutations
The rpoB gene was amplified by PCR using the primers rpobSP1F-5′TTATGCTGCACCTTCGTG3′; rpobSP5R-5′CAAGTGCCCATACCTCCCATC3′, designed to cover the entire region of interest. The constituents of the reaction mixture, in a final volume of 25 μl, were used in the following final concentrations: 1 μl of DNA, 0.5 pM of primer (up/down), 0.2 mM dNTP, 1 × PCR buffer, 1 U of Taq-DNA-polymerase. Amplification was carried out with a thermocycler Biometra T3000 Thermal Cycler (M-Medical Srl, Italy) according to the following program: a step of predenaturation for 5 min at 95°C; 30 cycles of annealing at 53°C; extension for 1 min at 72°C, and denaturation for 1 min at 95°C, followed by a final extension step of 10 min at 72°C.
The 1,024 bp PCR products obtained were purified and sequenced. The sequences of the rpoB gene were corrected and analyzed using the Chromas Lite 2.1 program; the sequences were exported in FASTA format. Sequence alignments were performed by using BLAST (Basic Local Alignment Search Tool;https://blast.ncbi.nlm.nih.gov/Blast.cgi) and UniProt programs (www.uniprot.org/-UniProtKB/Swiss-Prot UniProt release 2017_02).
Modeling study of RpoB
The bioinformatic program Swiss-Model ExPASy (http://swissmodel.expasy.org) was used to obtain the Protein Data Bank (PDB) format of RpoB protein from FASTA format. The PDB file was of three-dimensional (3D) structures, determined experimentally, of biological macromolecules, which contains atomic coordinates, information on the primary and secondary structure, and other information, collected during the process of X-ray crystallography.
To display the PDB file we used PyMol (https://www.pymol.org/), an open source software 3D graphics program used for the representation of biomolecules in biochemistry, bioinformatics, and structural biology. 22 Common data formats enable PyMol to run on different operating systems (multi-platform), with high-quality graphics rendering. PyMol was used for the calculation and the visualization of the molecular surface of the protein and the mutated amino acid residues of interest.
Results
Epidemiology of RIF-R MRSA
The 50 RIF-R isolates, analyzed in this study, belonged to diverse sources (SSTIs, n = 25; BSIs, n = 13; LRTIs, n = 12) and were all methicillin resistant; the hVISA phenotype was found in 30 RIF-R strains (60%) and 11 isolates were also DNS (22%).
All isolates belonged to the major multidrug-resistant (MDR)-MRSA clones circulating in Italy: n = 22 ST228-SCCmec I (spa-types t001-t041); n = 9 ST8-SCCmec IV/IVc (spa-types t008-t121-t2450); n = 8 ST239/241-SCCmec III (spa-types t030-t037); n = 7 ST5-SCCmec II (spa-types t2154-t3217-t002); n = 2 ST22/SCCmec IVh (spa-types t020-t902); n = 2 sporadic clones (ST247-SCCmec IA spa-type t051; ST63-SCCmec IV-spa-type t209). The most common clonal type observed among these strains was associated with the spread of ST228-SCCmec I, among which 86.3% were hVISA and 18.2% DNS (Table 1).
hVISA, heterogeneous vancomycin-intermediate Staphylococcus aureus; MIC, minimum inhibitory concentration; NS, nonsusceptible.
As expected, all strains were resistant to non-anti-staphylococcal cephalosporins (cefoxitin and ceftriaxone). Moreover, the majority of strains were susceptible to trimethoprim-sulfamethoxazole except for all ST239/241-SCCmec III strains, and two strains belonging to ST8 and ST247. All strains showed an MDR phenotype, being resistant to more than three antibiotic classes (gentamicin, 84%; ciprofloxacin, 82%; erythromycin, 76%; clindamycin, 74%; tetracycline, 50%).
The susceptibility of the main anti-Gram-positive drugs (linezolid, daptomycin, tigecycline, vancomycin, teicoplanin, dalbavancin, and ceftobiprole), related to the MRSA clones, was reported as MIC range, MIC50/90 values, and percentage of resistance (Table 1).
Tigecycline was found to be extremely active against almost all clones, with only few exceptions (n = 2, MICs >0.5 mg/L). Five strains belonging to the main HA-MRSA clones: ST228 (n = 2); ST5 (n = 2); ST8 (n = 1) were linezolid resistant, this was due to the G2576T mutation in 23S rDNA domain V.
Higher MIC90 and reduced susceptibility values—ranging from 50% (ST22) to 12.5% (ST239/241)—were found for daptomycin among all clones.
Dalbavancin showed MIC values one to two dilutions higher with respect to the breakpoint in nine hVISA strains, mostly belonging to the major ST228/SCCmec I clone. Dalbavancin MBC/MIC ratios for these strains were ≥16 mg/L.
Also for ceftobiprole, 10 strains showed decreased susceptibility, with MICs of 4 mg/L; 9/10 were ST228-SCCmec I, 7 of which were hVISA; the remaining strain was an hVISA belonging to ST239-SCCmec III.
Mutations in rpoB, RIF-R, and correlation with clones
rpoB sequence analysis revealed 13 amino acid substitutions in the RRDR, 10/13 in Cluster I, 2/13 in Cluster II, and only one in a region between Cluster I and II (Fig. 1).
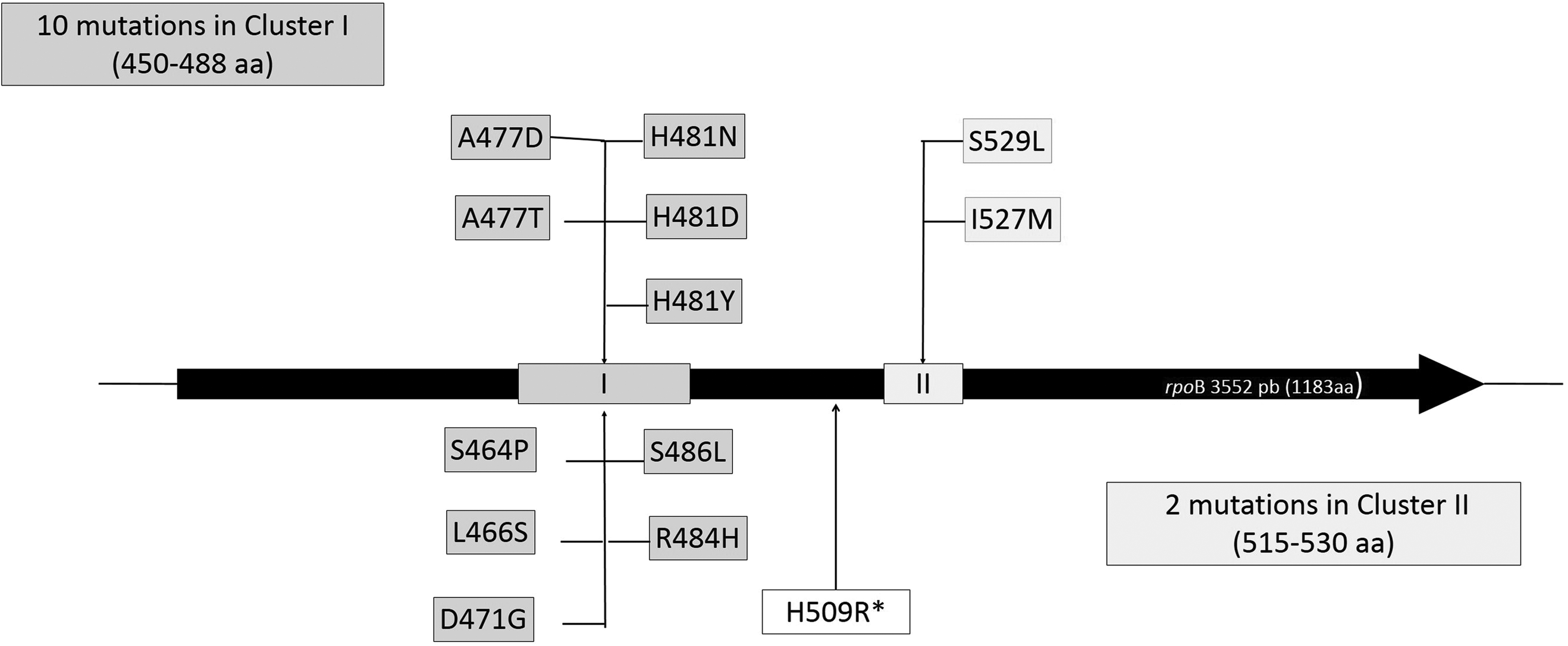
Position of the different rpoB mutations.
Table 2 shows the distribution of all RIF-R MRSA genotypes, rpoB mutations found, and the corresponding MIC values.
ACME, arginine catabolic mobile element; PVL, Panton-Valentine leukocidin.
All isolates harboring H481N as a single amino acid substitution showed low levels of resistance to rifampicin (MIC values ranging from 1 to 8 mg/L), and this was the most prevalent mutation (50%), found in 25/50 strains. In 11/50 strains (22%), H481N was found in combination with additional mutations in other sites (L466S; A477T; I527M; S529L), reaching an increased level of resistance (MICs ≥64 mg/L).
Substitutions of amino acids with a different chemical structure at the equivalent locus—H481D and H481Y (found in a DNS/hVISA strain)—or different loci (A477D; S486L), largely altering the antibiotic binding to the target site, conferred high-level resistance (MICs ≥64 mg/L).
Other less frequent rpoB variants were identified alone (H509R, MIC 16 mg/L; S464P, MIC 8 mg/L) and associated (R484H/D471G, MICs ≥64 mg/L) (Table 2).
We observed only few close correlations between rpoB mutations and clones:
• ST228-SCCmec I, ST239/ST241-SCCmec III, and ST8-SCCmec IV clones carried the H481N substitution, alone and in combination. • A477D was mainly associated with ST5-SCCmec II strains (belonging to different spa-types), and in one strain belonging to ST8-SCCmec IV-t008. All these strains were vancomycin susceptible S. aureus. • R484H/D471G linked mutations were always associated with ST8-SCCmec IVc-t008 clone. In addition, these isolates showed reduced susceptibility to daptomycin and the hVISA phenotype. • H481D was mainly associated with ST228-SCCmec I-t041 strains, and in one strain belonging to ST5-SCCmec II-t3217. • Only one isolate belonging to ST8-SCCmec IVc-t121 (hVISA) carried the H509R mutation. • Only one isolate belonging to ST22-SCCmec IVh-t902 (hVISA and DNS) carried the H481Y mutation. • The remaining S464P and S486L rare mutations were carried by ST228-SCCmec I-t001 and ST63-SCCmec IVh-t209, respectively.
Molecular modeling of RpoB and amino acidic substitutions
The 3D protein structure and the alteration of the molecule binding to the target site were reconstructed by PyMol analysis.
All mutations in rpoB leading to amino acid replacements were located in Cluster I (481, 477, 466, and 464 positions, and the rare R484H/D471G association) and in Cluster II (527 and 529 positions). A novel H509R mutation, located in the space between the two Clusters, was due to a replacement of a histidine with an arginine.
All substituted amino acids fell within the hydrophobic pocket of the rifampicin target site, and prevented antibiotic binding, with the exception of the H509R mutation, situated behind the hydrophobic pocket (Fig. 2).
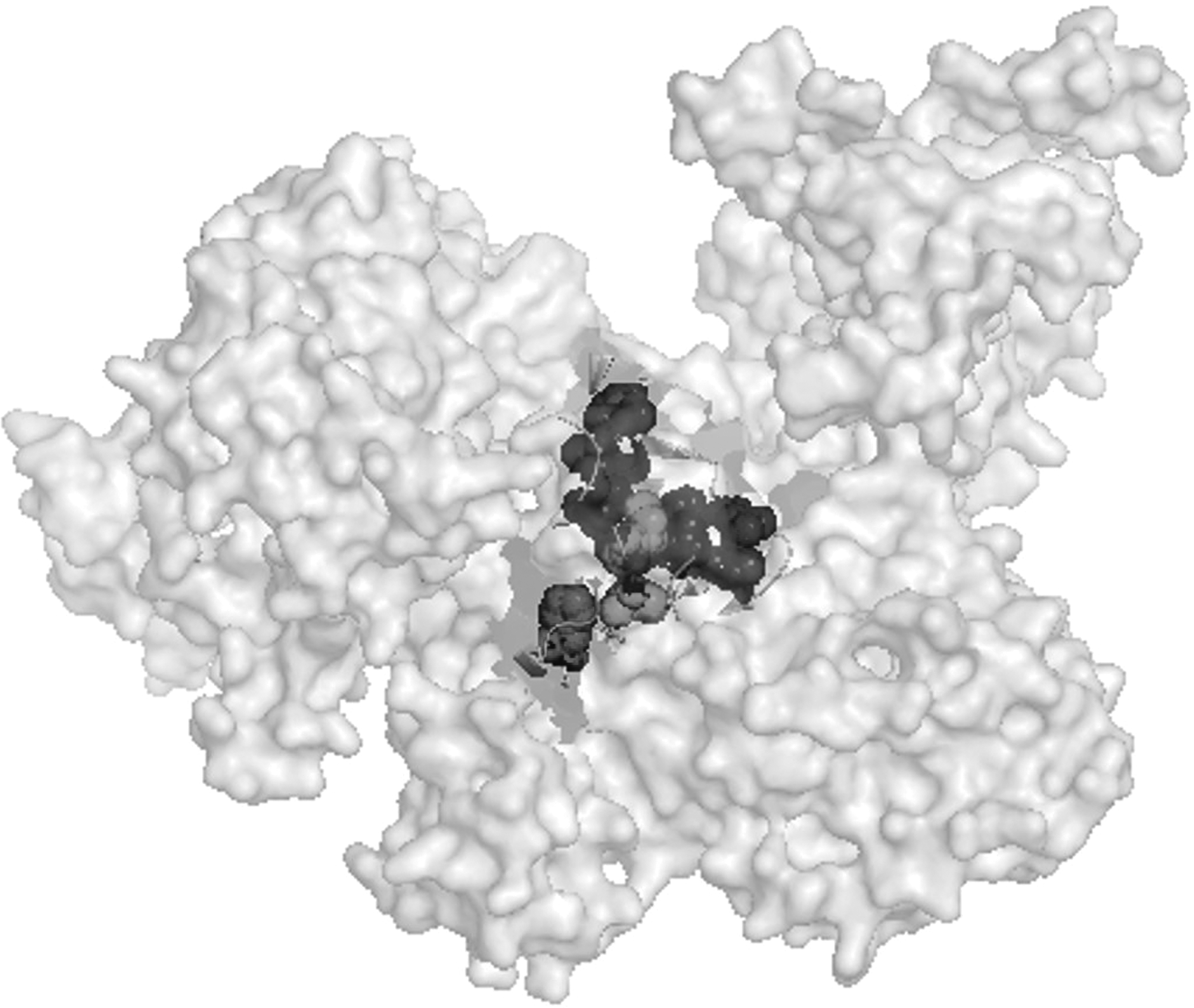
Modeling of the Staphylococcus aureus beta subunit, and the amino acidic residues analyzed in the study.
Discussion
In Italy, RIF-R has become an increasing trait of MDR-MRSA isolates (16.4%), with respect to the European trend (5.7%) as demonstrated by a recent surveillance study. 5 This trend is of great concern, especially after the reevaluation of this molecule for the treatment of severe and chronic infections caused by MRSA, often in combination with vancomycin and daptomycin. 23
All 50 RIF-R isolates belonged to the major MDR-MRSA clones circulating in Italy, with a higher prevalence of ST228-SCCmec I (n = 22, 44%), as already reported by HA-MRSA epidemiology, in Italy. 5 These isolates were MDR, with a high percentage of hVISA and DNS phenotypes, respectively 60% and 22%.
There was a subtle decrease in the activity of dalbavancin. These strains were dalbavancin tolerant. Although fewer data in the literature are available on tolerance to dalbavancin, 24 these isolates exhibited the hVISA phenotype and belonged to the ST228-SCCmec I clone, reinforcing the hypothesis that particular clones show a greater propensity to spread and persist in hospital settings, and to acquire resistance or tolerance to diverse antibiotic classes, making them difficult to eradicate them.
Vancomycin and daptomycin decreased susceptibility was already indirectly correlated with rpoB substitutions and resistance to rifampicin.3,10,25 This alteration was plausibly due to the decrease in transcriptional complex stability, global rearrangement of the transcriptional products, and consequent destabilization of all the cellular structures, such as the cell wall and the cytoplasmic membrane. 26
Although Bæk et al. 10 reported that H477D substitution was implicated in decreased susceptibility to daptomycin (but also to vancomycin and beta-lactams), our results showed that, in our strains, this locus is only responsible for high-level resistance to rifampicin, and is largely clonal (ST5-SCCmec II). H477D was also implicated in reduced virulence traits, slow-growth, and modulation of Spx levels, an RNA polymerase-interacting stress regulator responsive to oxidation and cell wall antibiotics. 10 A correlation between further compensatory rpoB mutations (P519L and R484H) and Spx was recently discovered, 27 conferring new properties to RNA polymerase. An R484H mutation was found (associated with D471G, which did not affect RIF-R), 28 in two hVISA strains with daptomycin-decreased susceptibility, one of which was also linezolid resistant.
The H481Y rpoB mutation was also associated with strong modifications in the transcriptome profile that reduce virulence and increase persistence in an MRSA strain, compared to its isogenic parent strain. 29 Here, an H481Y substitution was found only in one strain isolated from a patient with bacteremia, belonging to ST22-SCCmec IVh/spa-type t902 with reduced susceptibility to daptomycin and with the hVISA phenotype.
In this study, combined antibiotic susceptibility and molecular analyses gave the relationship between RpoB amino acid substitutions, clones, and the susceptibility to the main anti-Gram-positive drugs among the 50 RIF-R MRSA strains isolated from hospitalized patients.
The occurrence of common rpoB mutations might be explained by the epidemic nature of many MRSA strains and, consequently, to the frequent selective exposure of these strains to rifampicin. While not clonally related, rare mutations arise at a low frequency among less diffused MRSA epidemic clones.
As already described in the literature, position 481 represents a hot spot for amino acid replacement, with a different impact on RIF-R and resistance to other antimicrobial drugs.28,30
In fact, H481N was the most widespread mutation found in our sample, responsible for low-level RIF-R. The higher resistance values (MIC ≥64 mg/L) were also attributed to H481D and H481Y substitutions, influenced by the chemical features of the substituted amino acid. In particular, the replacement of a histidine (H) with a tyrosine (Y)—aromatic amino acid that is larger than asparagine (N)—was responsible for a considerable steric hindrance in the hVISA ST22-SCCmec IVh/spa-type t902 strain with reduced daptomycin susceptibility.
The substitution of a histidine with an arginine (R) at position 509, conferring an MIC of 16 mg/L, was found in a hVISA strain belonging to ST8-SCCmec IVc-spa-type t121 (CA-MRSA-USA300-like). Even if this substitution did not face the rifampicin target region, it could induce a conformational change that indirectly prevents antibiotic binding to the target site, thus determining RIF-R. The presence of out-of-cluster mutations conferring RIF-R was rare, but already described by other authors in Streptococcus pyogenes 31 and Mycobacterium tuberculosis. 32 The occurrence of this novel mutation could be related to its source of isolation (orthopedic ward) (data not shown) where rifampicin, often in association with other antibiotics, in particular vancomycin, for therapeutic purposes is commonly used. 11
High-level resistance to rifampicin was ascribed to multiple mutations, including H481N, indicating a step-by-step mechanism in resistance development, probably due to exposure to rifampicin therapy.
In conclusion, the results of this study show the nature and location of mutations in the rpoB gene among the Italian MRSA hospital isolates. All RIF-R strains belonged to the major genetic backgrounds with a strong prevalence of ST228, one of the major MDR clones with a strong association with reduced susceptibility to glycopeptides (hVISA). The spread of RIF-R strains in clinical settings represents a serious threat, due to their complex resistance nature, even to new anti-Gram-positive drugs, making these infections particularly difficult to treat.
Footnotes
Acknowledgments
Some of the results of this study here have been presented at the 26th ECCMID congress (P1344) and at the 44th Italian Society of Microbiology (SIM) congress (P127). We wish to thank the Scientific Bureau of the University of Catania for language support. This work was partially supported by the National Operational Programme for Research and Competitiveness 2007–2013 (project number PON01_02589) and by Prof. Stefania Stefani research grants.
Disclosure Statement
No competing financial interests exist.