
Abstract
Alterations in PBP2a have been recognized in cefotaxime-resistant laboratory mutants and β-lactam–resistant clinical isolates of Streptococcus pneumoniae. DNA sequencing revealed fundamental differences between these two settings. Internal stop codons in pbp2a occurred in all three laboratory mutants analyzed, caused by a mutation in pbp2a of mutant C604, and tandem duplications within pbp2a resulting in premature stop codons in another two mutants C403 and C406. In contrast, mosaic PBP2a genes were observed in several penicillin-resistant clinical isolates from South Africa, the Czech Republic, Hungary, and in the clone Poland23F-16, with sequence blocks diverging from sensitive strains by over 4%. Most of these pbp2a variants except pbp2a from the South African strain contained sequences related to pbp2a of Streptococcus mitis B6, confirming that this species serves as reservoir for penicillin-resistance determinants.
Introduction
S
The first report of two S. pneumoniae isolates with decreased susceptibility to penicillin in 1960, 1 which remained almost unrecognized, was followed in the late 1970s by the description of high level penicillin–resistant S. pneumoniae (PRSP) also expressing resistance to multiple non-β-lactam antibiotics in South Africa. 2 Since then, PRSP has spread rapidly worldwide, and high rates have been reported in many countries, as documented in numerous publications (for reviews, see McGee et al. 3 and Henriques-Normark 4 ). PRSP especially poses severe therapeutic problems for the treatment of meningitis.5–7
The introduction of a 7-valent pneumococcal conjugated vaccine in 2000 followed by a 10-valent and a 13-valent vaccine in 2009 directed against the most frequent serotypes of PRSP and highly virulent serotypes resulted in a significant decrease of infections caused by PRSP. However, new antibiotic-resistant clones expressing nonvaccine serotypes have appeared due to serotype switching within clones and dissemination of antibiotic resistance determinants into new clones, a result of the fabulous property of genetic transformation of the pneumococcus.8–10 To understand the mechanisms of antibiotic resistance and the route of gene acquisition is therefore of great importance.
Resistance to β-lactams is a multifactorial process, which involves alterations in penicillin-binding proteins (PBPs), the target enzymes for this class of antibiotics. PBPs are inhibited by β-lactams through formation of a covalent complex to the active site serine residue that is located in the conserved motif SXXK of the penicillin-binding/transpeptidase (TP) domain common to all PBPs. This motif together with another two conserved triads SXN and KS/TG form the active site cavity. PBPs are multidomain proteins and are grouped into different classes according to their domain structure: the high-molecular mass (hmm) PBPs of class A, bifunctional enzymes with an N-terminal glycosyl-transferase (GT) domain, hmm PBPs of class B with an N-terminal extension of unknown function, and the low-molecular mass (lmm) PBPs that act as D,D-carboxypeptidases.
S. pneumoniae contains six PBPs as follows: the hmm class A PBPs 1a, 1b, and 2a, two hmm class B PBPs PBP2x and 2b, and the lmm PBP3. Both class B PBPs are essential. 11 Individual class A PBPs are dispensable, but double mutants of PBP2a and PBP1a could not be obtained.12,13 PBP2a mutants showed a higher susceptibility to the antibiotic moenomycin,12,13 an inhibitor of GT activity. In fact, PBP2a derivatives show in vitro GT activity with lipid II, the precursor of peptidoglycan biosynthesis,14–16 and cross-linking activity as well. 14 The five hmm PBPs are localized in the division zone of the ovoid cells and play distinct roles in the septal and peripheral peptidoglycan synthesis machinery.17,18
In contrast, PBP3 localizes over the entire surface of the cells,19,20 with a concentration of PBP3 at the septa of some constricting cells. 20 PBP3 mutants are morphologically highly altered and contain misplaced division septa.20,21 This indicates that PBP3 somehow controls cell division indirectly, either by interfering with the assembly of the divisome or by affecting the enzymes required for peptidoglycan synthesis due to the presence of an abnormal pentapeptide-rich peptidoglycan.
The three PBPs 2x, 2b, and 1a are the main players in the development of β-lactam resistance of clinical isolates. In penicillin-resistant strains, these PBPs display a reduced affinity to β-lactams, and thus, higher concentrations of the drug are required for inhibiting the enzymes and consequently cellular growth in vivo. While PBP genes are highly conserved among penicillin-sensitive strains, PRSP contains highly altered PBP2x, PBP2b, and PBP1a genes due to mosaic blocks with sequences diverging over 20% at the DNA level or 10% at the amino acid level.22–24 Evidence has been provided that especially the commensal Streptococcus mitis serves as reservoir for PBP genes involved in the resistance phenotype through gene transfer followed by recombinational events.25,26 This and subsequent intraspecies recombination and acquisition of mutations result in a highly diverse population of these PBP alleles.27,28
Mutations in PBP2x and PBP2b confer low-level resistance without alterations in other proteins, 29 but since PBP2b does not interact with cefotaxime and other antibiotics with a related side chain it is not involved in cefotaxime resistance. 30 High penicillin resistance levels require alterations in PBP1a as well (for reviews, see Hakenbeck et al. 31 and Zapun et al. 32 ). Rarely, alterations in PPB1b, PBP2a, and PBP3 contribute to resistance, and their sequences are well conserved among PRSP.33–35 An authentic stop codon within the TP domain of pbp1b occurs in a high level penicillin–resistant Streptococcus mitis B6 and confers a marginal resistance increase in transformation experiments, 36 and a single cefotaxime-resistant laboratory mutant contained a mutated PBP3. 35
Occasionally, PRSP contains an altered PBP2a, which contributes especially to cefotaxime resistance,37–39 probably because PBP2a has particular low affinities to penicillins relative to cefotaxime affinity. 40 Efficient transfer of PBP2a from high-level resistant S. mitis strains to S. pneumoniae has been documented under laboratory conditions.36,41 Moreover, an altered PBP2a is present in several cefotaxime-resistant laboratory mutants where it cannot be detected by the common labeling procedure with radioactive or fluorescent β-lactam derivatives. 42
In the present study, pbp2a of the cefotaxime-resistant laboratory mutants and a collection of high-level clinical PRSP isolates from different locations were analyzed to identify mutations mediating reduced penicillin affinity and thus being potentially involved in the resistance phenotype.
Materials and Methods
Bacterial strains, plasmids, and growth conditions
Streptococcus pneumoniae R6 is a nonencapsulated derivative of the Rockefeller University strain R36A. 43 The cefotaxime-resistant mutants, all derived from the R6 strain, are independent spontaneous mutants described previously 42 ; R6 derivatives used in the present study containing various combinations of PBP genes are listed in Table 1. The streptomycin-resistant R6 derivative containing the rpsL41 allele was needed to replace wild-type pbp2a by other pbp2a alleles using the Janus gene replacement procedure. 44 R6ciaR::aad9 has been described. 45
Number in brackets indicates the regions of duplicated nucleotides. C403: C84TTACGTC91; C406: A1569AATGAAGGTTTAATGCCTGAAGCTCATTTTATTAGTAGAATTGAAAATGCTAGTGGACAAGTTATTGCGAGTCATAAAAATTCACAAAAACGGGTGATTGATAAGTCTGTAGCTGACA1687.
MICs were obtained by at least three independent experiments with 0.05 μg/ml dilution steps of CTX. One number indicates identical MICs in the experiments.
MIC value taken from Krauß and Hakenbeck. 35
CTX, cefotaxime; Ery, erythromycin; Kan, kanamycin; MIC, minimum inhibitory concentration; Spc, spectinomycin; Str, streptomycin.
Clinical isolates of S. pneumoniae have been described: strains 8249 and 2904422; strains from Poland 46 ; and Romanian isolates were obtained from Marina Pana, Bucharest. 47 Pneumococci were cultivated without aeration in C-medium 48 supplemented with 0.2% yeast extract (Difco) at 30°C or on D-agar plates 49 supplemented with 3% defibrinated sheep blood. Growth in liquid culture was monitored by nephelometry and is given in nephelometry units (N). Escherichia coli DH5a [Φ80dlacZΔM15 Δ(lacZYA-argF) recA1 endA1 hsdR17 supE44 thi-1 gyrA96 phoA relA1] was used for propagation of plasmids. Escherichia coli BL21 [B: F− dcm ompT hsdS (rB− mB−) gal] was used for propagation of the pGEX-6P-1 derivative expressing GST-PBP2a*. E. coli strains were grown in LB medium or on LB agar. 50 Growth of E. coli was followed by measuring of the optical density at 600 nm using a spectrophotometer.
Transformation experiments
Transformation of S. pneumoniae was performed according to published procedures 51 ; for transformation of C303 and C306 as recipient, cells were grown in the presence of 20 ng/ml of competence stimulating peptide to overcome the competence deficiency due to ciaH mutations in these strains. Cells were incubated for 30 minutes with DNA at 30°C, followed by a 2-hour phenotypic expression period at 30°C. Antibiotic resistance genes used for chromosomal integrations in S. pneumoniae were selected with 200 μg/ml kanamycin (Kan), 200 μg/ml streptomycin (Str), or 20 μg/ml spectinomycin (Spc). In case of altered pbp2a, at least five transformants from each experiment were analyzed for PBP profiles and cefotaxime MICs, and at least two transformants were selected for DNA sequencing of pbp2a; correct DNA sequences of pbp2x and ciaH were also confirmed by sequencing of PCR products using oligonucleotides listed in Table 2. E. coli was transformed using chemically competent cells, 50 and transformants were selected in the presence of 100 μg/ml ampicillin, 15 μg/ml tetracycline for pGEX0-, or 500 μg/ml erythromycin for pJDC9-derivatives.
Lowercase indicates sequences not matching the PBP gene.
PBP, penicillin-binding protein.
DNA manipulations and oligonucleotides
All DNA manipulations were performed using standard methods. 50 Chromosomal DNA was isolated from S. pneumoniae as described 52 ; in case for S. mitis strains, lysozyme (25 mg/ml) plus cellosyl (0.5 mg/ml) was added during the lysis procedure as described. 53 Plasmid purification from E. coli was done according to the manufacturer's protocol using the QIAprep Spin Miniprep Kit (Qiagen).
Restriction enzymes and T4 DNA ligase were purchased from New England Biolabs and Roche Diagnostics and used as described by the manufacturers. PCR products and DNA recovered after restriction endonuclease digestions were purified using a JetQuick® PCR Purification Spin Kit (Genomed). DNA fragments from agarose gels were purified using NucleoSpin® Extract II (Macherey-Nagel). PCRs from DNA were performed using GoldStar Red Taq polymerase (Eurogentec) or high fidelity iProof™ High Fidelity DNA polymerase (Bio-Rad), and in case of S. pneumoniae cells with DreamTaq DNA polymerase (Thermo Fisher Scientific) according to the manufacturer's instructions. The oligonucleotides used in this study are listed in Table 2 and were obtained from MWG-Biotech and Eurofins MWG Operon. The inserts of recombinant plasmids were sequenced to confirm the insertion and subsequently used to transform pneumococcal strains. Plasmid integration into pneumococcal chromosome was verified by PCR amplification and sequencing.
Amplification and sequencing of PBP2a genes
PBP2a genes from different S. pneumoniae strains and mutants were amplified using the oligonucleotide pairs 2a1U plus 2adown and pbp2afarup plus pbp2afardown2. PBP2a genes from clinical strains were amplified with 2a2U or 2a1U plus 2a2268R. Streptococcus mitis B6 pbp2a was amplified with 2a1U_B6 and B6_2aR. Amplification of chromosomal DNA with PCR was carried out in a biomed Thermocycler for 30 cycles of denaturation at 96°C for 30 seconds, annealing at 52°C for a 1 minute and extension at 72°C for 1 minute, followed by a 3-minute delay period at 72°C after the last cycle. The reaction mixture (100 μl) contained 10 pmol of each oligonucleotide primer and 2.5 U of Taq polymerase (PerkinElmer, Norwalk, CT). PCR amplified DNA was directly sequenced using the ABI PRISM Dye Terminator Cycle Sequencing Ready Reaction Kit (PerkinElmer). Automatic cycle sequencing was conducted with an ABI 377 sequencer.
Construction of mutants and plasmids
Insertion-duplication mutagenesis of pbp2a was performed using an internal 616-bp fragment of pbp2a (nucleotide 984–1,600) cloned into the vector pJDC9. 54 First, a pbp2a fragment was PCR amplified from Streptococcus pneumoniae R6 with the primer pair FU4 + FL4 and cloned directly into the pGEM®-T Easy Vector (Promega) suitable for cloning of PCR products. After restriction with EcoR1, the insert was cloned into pJDC9 resulting in the plasmid pIM2a. Plasmids were transformed into S. pneumoniae and transformants were selected with 1 μg/ml erythromycin. Insertion and orientation of the pJDC9 constructs were examined using the primers 2a_5U located within the pbp2a fragment, in combination with primers 1201S and 1211S upstream and downstream of the integrated region. The successful disruption of pbp2a in the S. pneumoniae transformants was verified by PBP profile analysis and DNA sequencing.
Different pbp2a variants were introduced into Streptococcus pneumoniae R6 and derivatives by the Janus gene replacement procedure. 44 Three PCR fragments were amplified containing the kan-rpsL region (primers janFOR and janREV), 44 the 5′-region of pbp2a (primers 2a1U and 2aJANr_link), and the 3′-pbp2a region (primers 2a2268r and 2aJANf_link). Fragments were purified and combined by overlapping PCR. 55 PCR products were transformed into R6rpsL41 and transformants selected with 200 μg/ml kanamycin. The resulting strains expressing a StrS and KanR phenotype (rpsL41 pbp2a:: kanR rpsL) were used as recipients for the various pbp2a alleles by selecting streptomycin-resistant transformants obtained with PCR amplified chromosomal DNA. Presence of the correct pbp2a allele was verified by DNA sequencing. The presence of rpsL41 does not influence CiaRH-mediated regulation. 56
To construct a strain producing PBP2a with specific mutations, site-directed mutagenesis was performed by an overlapping PCR approach. A PCR fragment carrying the mutation A1231>G was obtained with the primers 2a1U, 2aT>Ar, 2aT>Af, and 2a2268r to obtain R6pbp2aA411. DNA of this strain was then used to obtain a PCR fragment with primers 2a1U, 2aQ>Lr, 2aQ>Lf, and 2a2268r to obtain R6 pbp2aA411pbp2aLeu431. PCR fragments were cloned into the pGEM-T Easy Vector before transfer into Streptococcus pneumoniae R6 through the Janus replacement procedure as previously described. R6 containing the PBP2x mutation Ala550 was obtained by transformation with pbp2xC403 and selection with cefotaxime. The correct mutations were verified by DNA sequencing.
Replacement of pbp2a with the kanamycin cassette aphIII
PCR fragments were amplified and primer pairs shown in brackets representing sequences upstream (fragment 1; pbp2a up f CS and PM 364) and downstream (fragment 2; pbp2a down r1 and PM 363) of pbp2a. The kanamycin resistance cassette aphIII was obtained by cleavage of plasmid pUC19 aphIII with EcoRI and XbaI. The three PCR fragments were ligated after restriction of fragment 1 with EcoRI and fragment 2 with XabI. Transformation into the recipient S. pneumoniae strains and selection with kanamycin resulted in replacement of pbp2a with aphIII. Correct integration of the aphIII was verified by PCR of S. pneumoniae cells and the primer pair pbp2a up ext CS/pbp2a down r CS.
Detection of PBPs
Preparation of samples, PBP labeling with BocillinFL, and separation of proteins by sodium dodecyl sulfate (SDS)–polyacrylamide gel electrophoresis (PAGE) were carried out as described previously. 57 In case of S. mitis, cells were lysed by adding 0.5 mg/ml cellosyl and 0.8 mg/ml lysozyme to the lysis buffer. 53 BocillinFL-PBP complexes were visualized by fluorography detection with a FluorImager 595 fluorescence scanner (Molecular Dynamics) at 488 nm.
After SDS-PAGE, the proteins were transferred to a PVDF membrane (Roche Diagnostics) and probed with affinity purified polyclonal anti-PBP2a antibody followed by incubation with alkaline phosphatase-conjugated goat anti-rabbit immunoglobulin G (Sigma-Aldrich) and staining with 4-nitrobluetetrazolium chloride and 5-bromo-4-chloro-3-indolylphosphate (Roche Diagnostics). The presence of equal amounts of protein in the samples was verified on separate SDS-polyacrylamide gels by Coomassie blue staining.
Anti-PBP2a antibodies
A soluble PBP2a GST-tagged (glutathione S-transferase) derivative was constructed and coupled to GSH-Sepharose. In brief, a PCR product of pbp2a between T234–T2195 (residues 78–731) was cloned into the vector pGEX-6P-1-tet 58 after restriction with BamHI and XhoI and ligation. The resulting pbp2a* was overproduced in Escherichia coli BL21 and purified as described by chromatography on GSH-Sepharose. 58 Rabbit anti-PBP2a antisera were obtained from Seqlab GmbH and affinity-purified using PBP2a* coupled to 1 ml HiTrap NHS-activated Sepharose (GE Healthcare). After washing with 20 ml sodium phosphate buffer (10 mM, pH 7.2), PBP2a-specific antibodies were eluted with glycine (100 mM, pH 2.2). Fractions (3 ml) were collected directly in 250 μl Tris buffer (1.5 M, pH 8.8) and used at a dilution of 1:2,000.
Results
PBP2a in high level cefotaxime–resistant S. pneumoniae
Cefotaxime-resistant mutants isolated from the laboratory strain Streptococcus pneumoniae R6 have been described in which PBP2a was not detectable after labeling with β-lactams and protein separation on SDS-polyacrylamide gels, neither with radioactive β-lactam nor with anti-β-lactam antibiotics where high penicillin concentrations for labeling of PBPs can be applied to detect PBP-β-lactam complexes on Western blots.42,59 This phenotype was observed in three independently isolated mutants with increasing concentrations of cefotaxime: C403and C406 obtained after four selection steps and C604 obtained after six selection steps. All three mutants contained mutations in PBP2x and CiaH and C604 one mutation in PBP3 as well (Fig. 1).
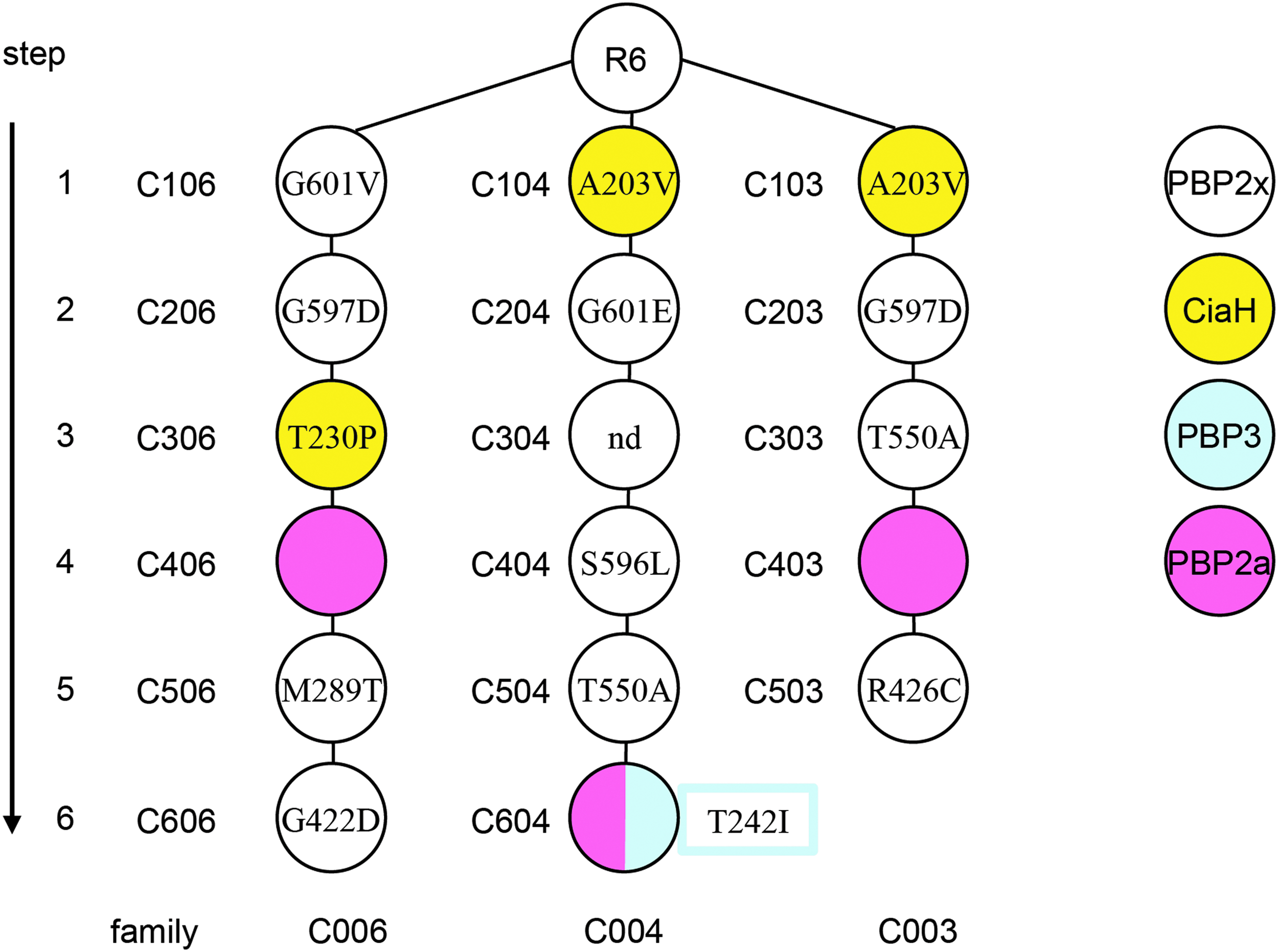
Cefotaxime-resistant lineages of laboratory mutants. The cefotaxime-resistant mutant lineages and the occurrence of PBP2a mutations during the selection procedure are shown. Mutations identified in PBP2x, 52 PBP3, 35 and CiaH 73 are shown. The first digit of the mutant indicates the selection step, and the last digit specifies the mutant lineage.
To identify PBP2a mutations, pbp2a was amplified by PCR from chromosomal DNA of the three mutants for DNA sequencing. Surprisingly, the PCR fragment obtained with C406 was larger than that compared to the PCR products of the other two mutants and the parental strain R6, indicating some type of insertion in pbp2aC406 (Fig. 2A).

PBP2a in cefotaxime-resistant laboratory mutants.
DNA sequencing revealed that it contained a 119 nucleotide tandem duplication within pbp2a (nucleotide 1,569–1,687), resulting in a change of amino acid composition after residue 564 and a truncated predicted peptide of 566 amino acid compared to the 731 amino acid PBP2a of strain R6 (Fig. 2B). Similarly, pbp2a of C403 contained a tandem duplication of eight nucleotides (84–91) resulting in an altered peptide sequence after amino acid 30 and a stop codon after 42 amino acid, whereas C604 contained a single mutation in codon 215 TGG encoding Trp215 which was changed to TAG, resulting in a stop codon after amino acid residue 214 (Fig. 2B).
Thus, none of the three mutants apparently produced a functional PBP2a. Tandem duplications have been observed in a macrolide-resistant S. pneumoniae, which contained an 18-bp tandem repeat in the gene encoding the ribosomal protein L22. 60 Interestingly, such duplications have been recognized in phase variants of serotype 3, 8, and 37 S. pneumoniae, and the frequent isolation of capsular revertants indicated a precise excision mechanism. 61 No instability of the PBP2a mutants was observed.
Cefotaxime resistance mediated by the lack of PBP2a
Initial transformation experiments with chromosomal DNA of the mutants C403, C604, and C406, using C303 as recipient, resulted in a large number of transformants after cefotaxime selection with MICs similar to C403. Selected transformants all showed a lack of the PBP2a band after labeling with BocillinFL and SDS-PAGE on fluorograms (not shown). However, in this and subsequent experiments additional mutations in ciaH were detected in several transformants, potentially contributing to cefotaxime resistance, clearly indicating that cefotaxime selection is not the method of choice to determine the impact of pbp2a on the resistance phenotype. In particular strongly activating ciaH alleles, including ciaHC103, are genetically unstable, 56 and therefore, ciaH was sequenced in all constructs to verify the presence of the ciaH wild-type allele.
To avoid cefotaxime selection for testing whether a disrupted pbp2a indeed confers cefotaxime resistance, first pbp2aC403 was introduced into strain C303 through the Janus cassette, a bicistronic cassette conferring a KanR StrS phenotype, which enables the replacement of genes through negative selection in a two-step procedure. 44 Insertion of the Janus cassette in the first step results in the disruption of the gene, and thus, this technique can only be applied for nonessential genes, such as pbp2a,12,13 or nonessential regions adjacent to essential genes. Streptomycin-resistant recipients are required for this procedure mediated by the rpsL41 allele, which does not influence cefotaxime resistance. 56 We chose pbp2aC403 because of the clear difference in MIC values for cefotaxime between C303, which contains only mutations in PBP2x and CiaH, and C403, which shows affinity changes in PBP2a as well (0.3 μg/ml versus 1.1–1.2 μg/ml; Table 1).
Transformants were readily obtained, and sequencing proved the existence of the truncated pbp2aC403; consequently a PBP2a band could not be detected in their PBP profiles on fluorograms or by Western blotting using anti-PBP2a antibodies (Fig. 3, left side). Surprisingly, the MIC values of transformants obtained did not reach the MIC of the donor strain C403 by far (Table 1), similar to transformants obtained by insertion-duplication using the vector pJDC9 and which did not contain additional mutations in ciaH (not shown). The effect of a pbp2anull genotype was investigated in C303, as well as in C306, using allelic replacement of pbp2a with the gene aphIII conferring kanamycin resistance. Again, these transformants did not contain PBP2a (e.g., see Fig. 3, right side) and showed only a slight increase in resistance which did not reach the level of the next step mutant in which this mutation had been originally identified (Table 1).

PBP2a in cefotaxime-resistant laboratory mutants and transformants of the R6 strain. Cell lysates were labeled with BocillinFL (top), and PBPs were detected after SDS-PAGE and fluorography with the FluorImager. The PBP profile of the parental strain Streptococcus pneumoniae R6 is included for comparison. The positions of the PBPs are marked on the left side. Bottom: Western blot developed with anti-PBP2a antibodies. The arrow shows the position of PBP2a present in strains R6 and C306. PBP, penicillin-binding protein; SDS-PAGE, sodium dodecyl sulfate–polyacrylamide gel electrophoresis.
No attempts were made to use C504pbp3C604 as recipient in such experiments. It should be noted that relative differences in cefotaxime MICs between the various constructs (although as low as 0.05 μg/ml) were consistent in all experiments, including one final test that included all mutant constructs described in this study. However, differences below 0.05 μg/ml cefotaxime cannot be excluded.
The results strongly suggest that in addition to pbp2a mutations, other still unknown genes are affected in C403 and C406. Nevertheless, a slight effect on cefotaxime resistance in a genetic background with a mutated pbp2x was mediated by the absence of an intact PBP2a gene.
PBP2a in clinical isolates
PBP2a genes were examined in 19 PRSP of the Kaiserslautern clinical isolate collection. Isolates with very high β-lactam resistance levels were chosen, including isolates from Poland and Romania, countries where high rates of PRSP have been recorded and where affinity changes of PBP2a have been observed as described in strains 8249 (South Africa) and 29044 (Czech Republic). 22 In addition, another two pbp2a described in previous publications were used. These included pbp2a from a high level cefotaxime–resistant clinical strain from Japan SP00084. 38 Smith et al. 39 described three clonally related isolates with identical mosaic pbp2x, pbp2b, and pbp1a, whereas two (isolate 1 and isolate 2) contained the same altered pbp2a and expressed higher MIC values compared to isolate 3. Isolate 3 could be transformed to higher β-lactam resistance levels with pbp2a of isolate 1.
Moreover, pbp2a of the high-level β-lactam–resistant Streptococcus mitis strains B6 62 and DD28 63 was used for comparison, as were pbp2a of the penicillin-sensitive Streptococcus pneumoniae R6 and Streptococcus mitis strains NCTC12261, NCTC10712, and M3 as references.
The full-length pbp2a sequence encoding the 731 amino acid PBP2a was retrieved from genome data if available and was obtained for pbp2a of strain Streptococcus pneumoniae 29044. Sequences covering at least codons 258–651 were obtained in all other cases, which include most of the TP domain located between codons 301–678 as defined by Di Guilmi et al. 64 It should be noted that Streptococcus mitis B6 has an extension of two amino acids at the C-terminus due to the integration of an ISSmi1 element at the very end of pbp2a. 65
Properties of the strains and accession numbers of the genomes, respectively, PBP2a genes are listed in Table 3.
Serotype of Streptococcus pneumoniae strain R6 is that of the progenitor D39; strain 29044 is a member of clone CSR 14 -10; serotype 6B isolates from Poland belong to Poland6B-20, and all 23F isolates are members of the clone Poland23F-16 or single locus variants thereof. 46
CZ, Czech Republic; DE, Germany; GB, Great Britain; HU, Hungary; JP, Japan; na, not available; NO, Norway; PEN, benzylpenicillin; PL, Poland; RO, Romania; SA, South Africa; SP, Spain; SW, Sweden; USA, United States.
PBP2a gene sequences covering nucleotide positions 903–1,973 were used for a phylogenetic analysis with the MEGA6 program. Since mosaic genes were included in this analysis, the tree does not reveal the phylogenetic relationship, but rather was used to provide a visual aid to assess the sequence similarities. As shown in Figure 4, 11 sequences clustered together with pbp2a of the penicillin-sensitive reference strain Streptococcus pneumoniae R6 (R6-cluster), whereas all others were more dispersed. All pbp2a outside the R6 cluster diverged from pbp2aR6 by between 3.3% (SP00084) and 6.4% (Hu11) over the entire sequence.
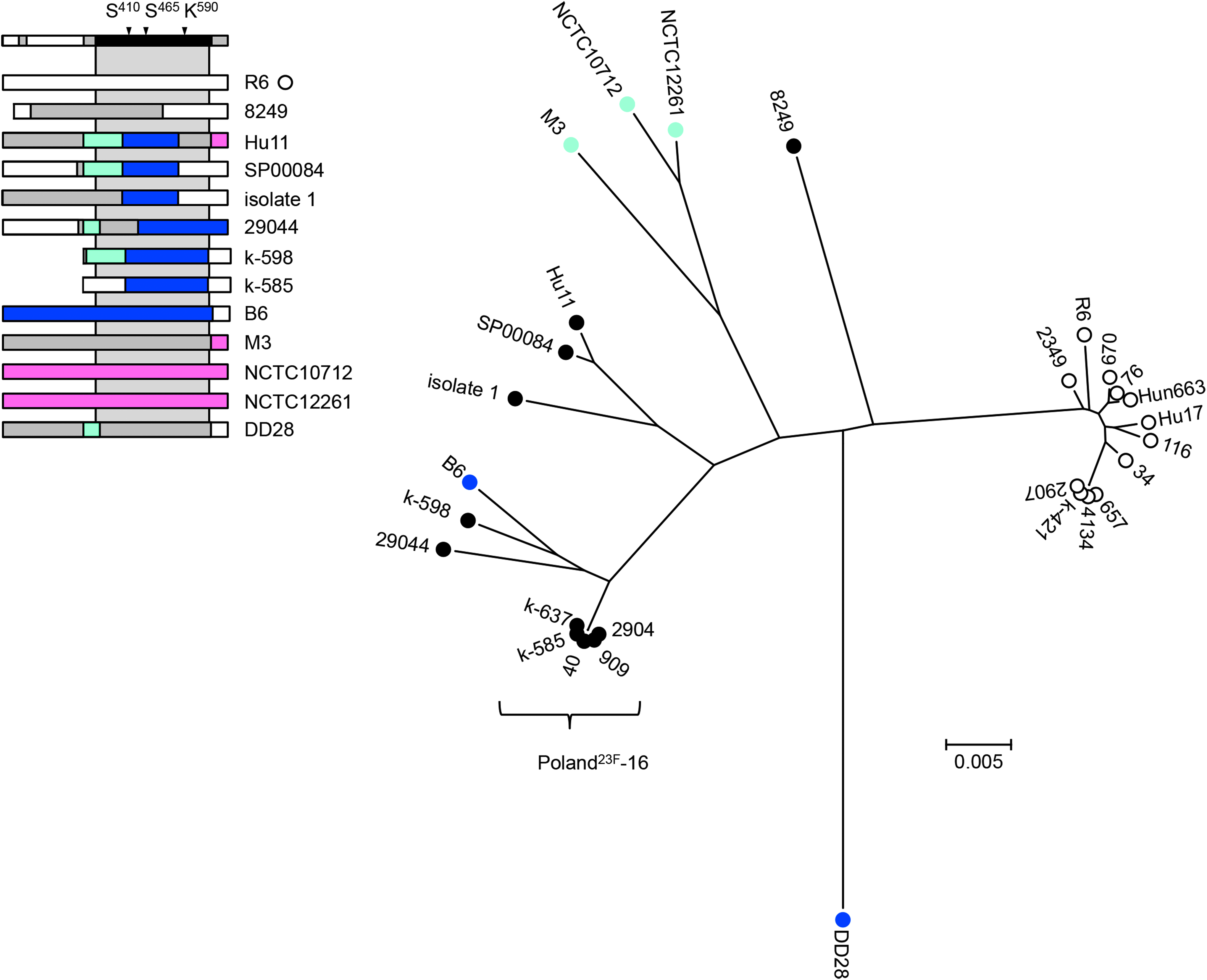
Phylogenetic tree of PBP2a genes. The tree was generated from pbp2a between codons 301 and 658 covering the TP domain. Bootstrap values (percentages) are based on 1,000 replications; the bar refers to genetic divergence as calculated by the MEGA6 software. 74 White circles: Streptococcus pneumoniae R6 related pbp2a (<1% nucleotide divergence); black circles: sequences differing from pbp2a of Streptococcus pneumoniae R6 by > 4.5% nucleotide; penicillin-susceptible S. mitis: light blue; penicillin-resistant S. mitis: dark blue. The mosaic structure is indicated on the left with the domain structure of PBP2a shown on top. The gray area represents the TP domain used for the phylogenetic tree. The colors mark pbp2a sequences diverging by <1%; gray: unrelated sequences differing by > 4.5%.
A closer look revealed sequence blocks almost identical to Streptococcus mitis B6 sequences in 11 of these sequences corresponding to a mosaic structure of pbp2a, whereas pbp2a of strain 8249 was distinct (Fig. 4). Five of the serotype 23F strains from Poland, all members of the clone Poland23F-16 or single locus variants thereof, 46 carried the same mosaic variant; pbp2a of strain 29044 was highly similar to that of the PRSP clone CSR14–10. This strongly indicates that occasionally pbp2a in PRSP has acquired sequence blocks from S. mitis by genetic transformation and, thus, represents a mosaic gene in particular clones. All other S. pneumoniae pbp2a were highly similar (<1% divergence) and differed from S. mitis pbp2a between 5.5% and 6.6%.
Distinct alleles occurred in S. mitis even in penicillin susceptible strains, similar to as described for other PBP genes.27,28 The PBP2a gene of Streptococcus mitis NCTC10712 differed from pbp2a of Streptococcus mitis NCTC12261 by only 0.9%, but from pbp2a of the other S. mitis by 5.7–5.9%. BLAST searches of unique pbp2a sequences revealed no match above 96% in case of Streptococcus pneumoniae 8249 and Streptococcus mitis DD28; Streptococcus mitis M3 pbp2a was highly similar to that of Streptococcus mitis SK578 (JPFY01000008). No sequence similarities with Streptococcus oralis Uo5 pbp2a, which differs from pbp2a of Streptococcus pneumoniae R6 by almost 20% and does not play a role in this high level β-lactam–resistant strain, 66 were detected in any of the pbp2a genes analyzed in this study.
Mutations in PBP2a of PRSP
So far, mutations relevant for penicillin resistance have only been identified within the TP domains of PBPs. Structural information for the 731 amino acid PBP2a-protein is not available. A PBP2a fragment spanning amino acid 301–678, obtained by limited proteolysis with trypsin of a purified PBP2a derivative, was still able to bind penicillin. 64 This fragment contains the three active site motifs and, thus, includes the TP domain. Therefore, we concentrated our search for mutations potentially related to β-lactam resistance on this region in PBP2a of clinical isolates.
The deduced amino acid sequences revealed a change of Thr411Ala, next to the active site Ser410, in all deduced PBP2a with B6-related sequence blocks, as well as in PBP2a of Streptococcus pneumoniae 8249 and Streptococcus mitis DD28 (Fig. 5). This mutation has been highlighted in strain SP00084 with high-level cefotaxime resistance. 38 Ala411 is also present in PBP2a of isolate 1, 39 and occurs in PBP2a of two PRSP strains 1983 and 55938 (as deduced from the available sequence reads), and contributes to β-lactam resistance as shown by genomic sequencing of Streptococcus pneumoniae R6 transformants obtained by serial transformations. 37
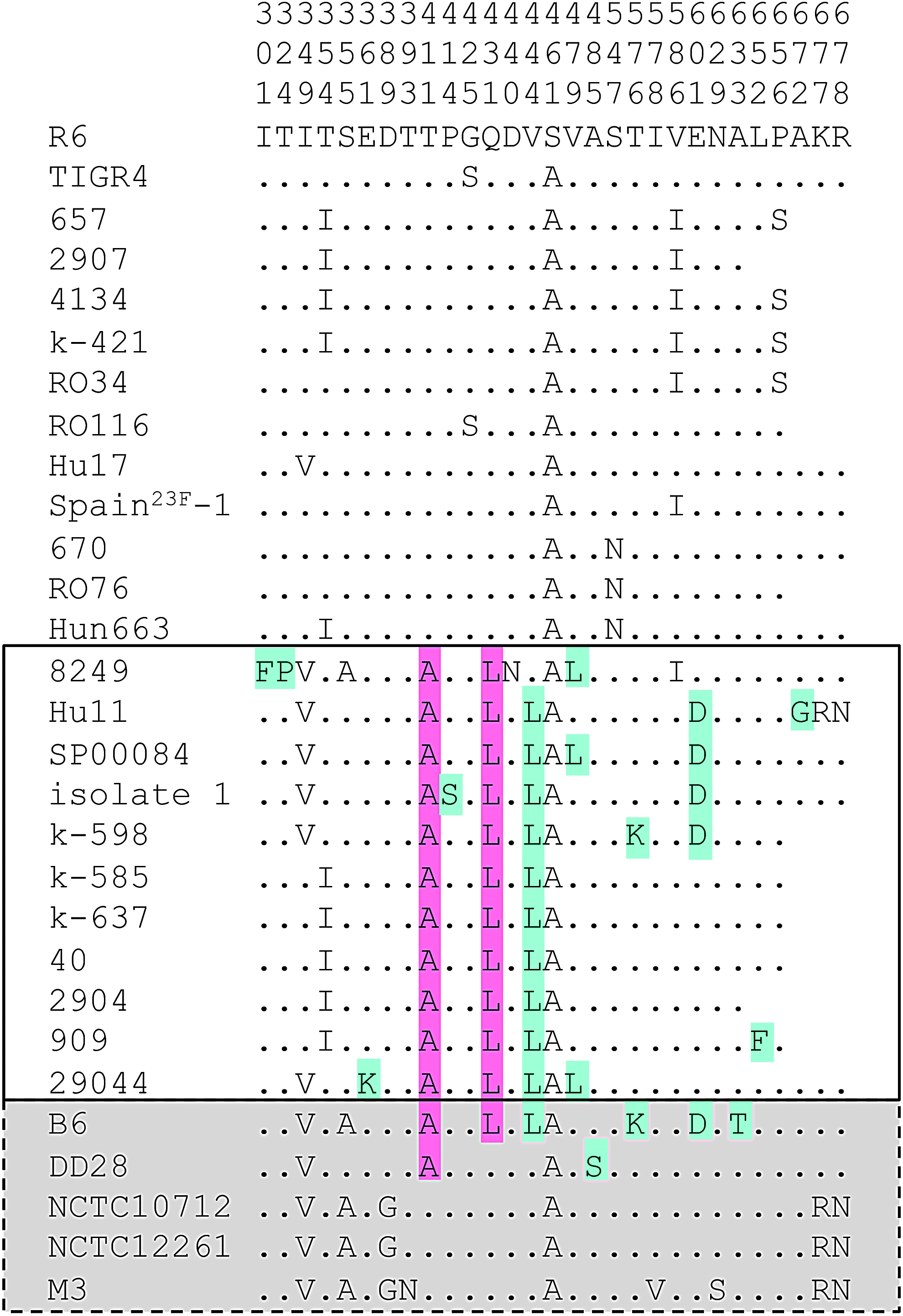
Deduced amino acid sequences of PBP2a in penicillin-susceptible and -resistant Streptococcus pneumoniae and Streptococcus mitis. PBP2a of Streptococcus pneumoniae R6 was used as a reference. The positions of the amino acids are indicated vertically in the top three rows. Only positions with altered residues are shown; residues identical to those in PBP2a of strain R6 are indicated by dots. The mutations A411 and L431 common to mosaic Streptococcus pneumoniae PBP2a (boxed) are highlighted in pink; light blue indicates mutations that occur only in a subset of sequences containing mosaic blocks (see text for details). S. mitis sequences are shaded gray.
Most PBP2a with Ala411 also contained the changes Gln431Leu and Val444Ala, except for PBP2a of strain 8249 which has Val444. Both mutations are absent in Streptococcus mitis DD28 PBP2a. Four PBP2a (strains Hu11, isolate 1, SP00084, and k-598) contained the mutation Glu601Asp. All these changes were present in Streptococcus mitis B6 PBP2a and absent in other PBP2a variants of S. pneumoniae with different levels of β-lactam resistance. Ile586, Ser425, Ile354, and Ser656, present in the nonmosaic PBP2a of our study, were also mentioned in a study by Chesnel et al. 34 It is interesting that the authors observed sequence differences in one pbp2a variant consistent with a recombination event, but this sequence type was found in a susceptible strain.
The presence of PBP2a mutations did not appear to be related to the resistance level of the strains. For example, the three Romanian strains with benzylpenicillin MICs of 4–8 μg/ml contained only amino acid changes that were present in susceptible S. pneumoniae such as strains R6 (Gly425Ser) and 657 (Val586Ile Pro656Ser); Ser547Asn is also present in strain Hun663 (clone Hungary19A-6) where PBP2a does not seem to play a role in penicillin resistance, 67 and in strain 670 (Spain6B-2). This is not surprising given the fact that other PBPs and non-PBP components as described above feed into the phenotypic expression of penicillin resistance. It is likely that murM contributes to high-level resistance as shown for PRSP isolates from Romania 68 and for members of the Hungary19A-6 clone67,69 where the ciaH232 allele also plays a role.56,70
Ala411 mediates low affinity to BocillinFL
Since the mutations Ala411 and Leu431 were prevalent among mosaic PBP2a from PRSP, mutated pbp2a was constructed by site-directed mutagenesis that contained the single Ala411 mutation or both, Ala411 plus Leu431. They were introduced into strain R6pbp2x2349 by the Janus procedure to investigate the role of these mutations. S. pneumoniae 2349 is a member of the PRSP clone Spain23F-1.36,41 The mutation Ala411 resulted in slow binding and low labeling intensity of BocillinFL, and binding reached a plateau of ∼30% compared to the value obtained for PBP2aR6 after 16 minutes (Fig. 6). The introduction of Leu431 led to an altered electrophoretic mobility so that PBP2x and PBP2aA411L431 were difficult to resolve on SDS-gels, and therefore, the affinity change could not be analyzed quantitatively in this case.
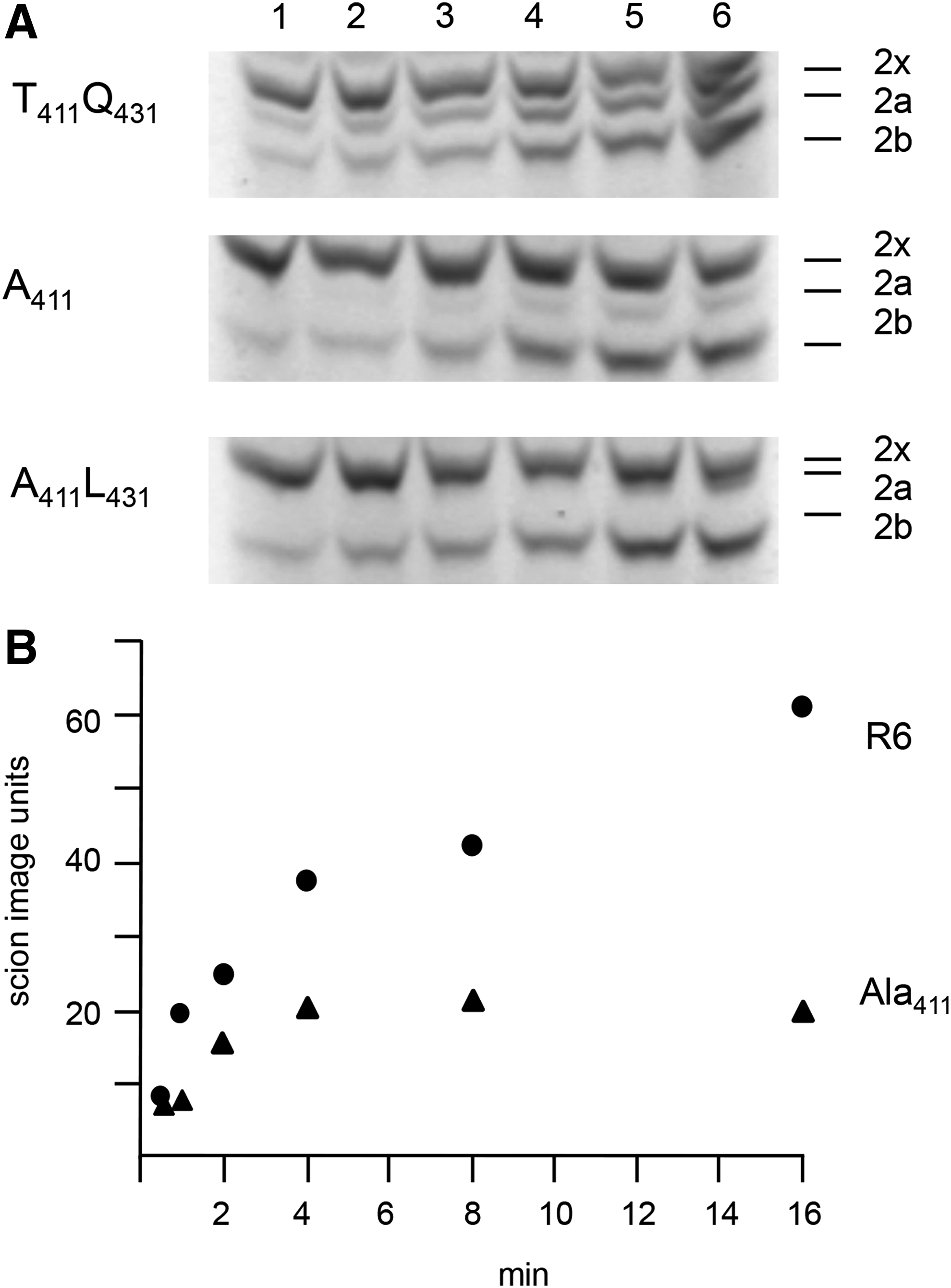
Binding of PBP2a with mutations Ala411 and Leu431 to BocillinFL.
When introduced into R6pbp2x2349, only minor changes in cefotaxime susceptibility were conferred by Ala411, or by both mutations Ala411 and Leu431, increasing the MIC from 0.3 (R6pbp2x2349) to 0.35 μg/ml only (Table 1). Since the affinity of PBP2a for penicillins is lower than for cephalosporins 40 it would be interesting to analyze the affinity for cefotaxime in the PBP2a mutant constructs with purified protein derivatives.
The full-length pbp2aB6, which contains the mutations Ala411 and Leu431, is known to confer cefotaxime resistance after transformation in Streptococcus pneumoniae R6 containing the PBP2x gene of Streptococcus mitis B6. 36 When introduced into the R6pbp2x2349 background, pbp2aB6 also mediated an over twofold increase of cefotaxime MIC from 0.3 to 0.65 μg/ml (Table 1), strongly suggesting that other mutations in PBP2aB6 are important for resistance. No difference in cefotaxime MIC was apparent between R6 and R6pbp2aB6, that is, in the absence of a mosaic pbp2x, at least within the range of 0.05 μg/ml used for MIC determination (Table 1). It should be noted that R6pbp2aB6 grew similar to the parental R6 strain and did not lyse rapidly as the strain R6pbp2aC403 (not shown).
Discussion
Clinical isolates versus laboratory mutants: a conclusion
Once more, distinct modes of penicillin resistance development became apparent in cefotaxime-resistant laboratory mutants versus clinical isolates in terms of PBP2a mutations. Clinical isolates have the opportunity to capture genes from other species resulting in the well-known mosaic gene structures of pbp2x, pbp2b, and pbp1a and, as described here, also of pbp2a in particular PRSP clones. Moreover, clinical isolates had sufficient time to evolve compensatory mutations that help to alleviate potential fitness loss associated with some mutations that increase the β-lactam resistance level.
The finding that mutants selected in the laboratory with cefotaxime contained truncated versions of pbp2a rather than mutations that lead to amino acid changes in the protein as seen in PRSP is in agreement with fundamental differences between these two settings. This has been pointed out before in context with other β-lactam resistance determinants: mutations selected in PBP2x and PBP2b are distinct from those identified in clinical isolates in most cases 31 ; ciaH mutations are frequent in laboratory mutants but rare in PRSP and less activating56,70; and finally, alterations in pbp1a which are a prerequisite for high resistance levels in PRSP have not been observed in laboratory mutants. PBP1a mutations in PRSP are likely to have altered enzymatic activity as suggested previously. 70
Since double mutants in PBP2a/PBP1a are not viable, the absence of an enzymatically functional PBP2a in the three cefotaxime-resistant mutants might prevent mutations in PBP1a. In this context it should be pointed out that R6 derivatives may express phenotypes distinct from its progenitor D39 due to the presence of ≥ 80 mutations in R6 compared to D39, 71 at least with respect to several genes implicated in cell wall biosynthesis as reported recently. 72 Therefore, Streptococcus pneumoniae R6 might not represent the ideal genetic background to evaluate the impact on β-lactam resistance of pbp2a mutations, and this should be considered in future experiments.
The exact function of PBP2a remains largely unknown. 17 Interestingly, it has been shown that PBP2a resides in complexes with the cytoplasmic protein GpsB, a major regulator of peptidoglycan biosynthesis and other cell division proteins, and that GpsB is most likely required for PBP2a activation. 72 The authors also mention a recognition sequence for GpsB present in the 51aa positively charged, cytoplasmic amino domain of PBP2a. We do not know whether PBP2a fragments are expressed in the cefotaxime-resistant mutants with premature stop codons in pbp2a, but it is suggestive that they all have the potential to encode the cytoplasmic tail at least up to Gln 41 , which includes the highly positively charged region R 30 RSRSDRKK. Thus it is possible that the mutations in the laboratory mutants maintain the ability for GpsB-PBP2a interaction to ensure proper PG synthesis.
Further studies will be required to identify the still unknown mutations selected in the laboratory by genome sequencing, to unravel the impact of individual PBP2a mutations on resistance in context with mutations in other genes, and to reveal the function of PBP2a in peptidoglycan biosynthesis and cell division.
Footnotes
Acknowledgments
This work was supported by the Bundesministerium für Bildung und Forschung (BMBF; Grant No. 01KI9402) and by the Deutsche Forschungsgemeinschaft (DFG; Grant Nos. Ha1011/11-2 and Ha1011/11-3) to R.H. The authors thank Richard Reinhard from the Max-Planck Institute for Molecular Genetics in Berlin for his help in automated DNA sequencing, Shwan Rachid for providing pGEX-6P-1-tet 2x-sol, and Patrick Marx for pUC19 aphIII. The help in constructing aphIII mutants by Anne Sexauer and Jasmine Theis is greatly acknowledged.
Disclosure Statement
No competing financial interests exist.