
Abstract
In Gram-negative bacteria, the outer membrane proteins (OMPs) perform a crucial role in antibiotic resistance, but it is largely unknown how they behave in response to antibiotic stress. In this study, we treated Aeromonas hydrophila with two different doses of oxytetracycline (OXY) to induce antibiotic stress. Proteins were isolated from sarcosine-insoluble fractions and quantitatively examined by using tandem mass tag labeling-based mass spectrometry to identify differentially expressed proteins. As a result, we identified 125 differential proteins in the 5 μg/ml OXY treatment group, including 20 OMPs, and 150 proteins from the 10 μg/ml OXY group, including 22 OMPs. Gene ontology analysis showed that translation-related proteins, including 30S and 50S ribosome proteins, were significantly enriched in increasing abundance under OXY stress; whereas the downregulated proteins were associated with the transport process, such as maltodextrin, maltose, and oligosaccharide transport. We then validated a subset of the identified differential proteins by using Western blot and quantitative polymerase chain reaction analyses. Finally, the quantitative real-time PCR (qPCR) results showed that at the transcription level, the expression of five OMP genes, including AHA_1280 (protein name A0KHS0), AHA_1281 (A0KHS1), AHA_1447 (A0KI84, BamE), AHA_1861 (A0KJE1), and AHA_2766 (A0KLX3), and one lipoprotein gene AHA_1740 (A0KJ25) was consistent with proteomic results under 5 and 10 μg/ml OXY treatment, respectively. In addition, the Western blotting also demonstrated that two altered OMP proteins A0KHS1 and A0KHH2 were upregulated for both OXY treatment groups. This study indicates that bacteria regulate the expression levels of OMPs in response to antibiotic stress and further contribute to our understanding of the functions of OMPs in antibiotic resistance. Moreover, our results suggest that the upregulation of translation and downregulation of the transport process may affect bacterial fitness during OXY stress. These findings may provide new clues to the antibiotic resistance mechanism in A. hydrophila.
Introduction
A
In bacteria, the efflux transport system functions to pump out undesired substances, such as antibiotics and toxins. In E. coli, the OM channel protein TolC associates with AcrA and AcrB to form the AcrAB-TolC efflux pump, which plays a major role in multidrug resistance. 9 Further, some studies identified other efflux pumps as contributing to antibiotic resistance in Pseudomonas aeruginosa, such as MexAB-OprM, MexXY-OprA, MexCD-OprJ, and MexEF-OprN. These studies indicate that OMPs exert a significant influence in antibiotic resistance in Gram-negative bacteria.10,11 However, whether the A. hydrophila OM subproteome contributes to an OXY-induced stress response remains elusive.
In this study, we examined A. hydrophila OMP expression in response to different doses of OXY stress by using a quantitative tandem mass tag (TMT) labeling proteomic approach. We then analyzed the identified differentially expressed proteins using bioinformatics to determine protein functions and predicted protein subcellular locations. In addition, a subset of the identified differentially expressed OMPs was further validated by using quantitative polymerase chain reaction (qPCR) and Western blot assays. The results present herein show that the OMPs have a significant impact on drug resistance in Gram-negative bacteria.
Materials and Methods
Bacterial strains and cultivation
The A. hydrophila ATCC 7966 strain was maintained at a minimum inhibitory concentration (MIC), and the ratio of A. hydrophila ATCC 7966 to OXY was 2.5 μg/ml, as previously described. 12 Cells were cultured in Luria Bertani (LB) medium at 30°C overnight and diluted 1:100 with fresh LB medium for further culturing. Once the OD600 reached 1.0, we added OXY to the culture at a concentration of either 5 or 10 μg/ml, that is two- and four-fold of the MIC concentration, respectively. We cultured the mixtures for 1 hr and then harvested the bacterial cells and washed them twice with phosphate-buffered saline (PBS, pH 7.4).
Extraction of OMPs
We isolated OMPs as previously described by using a sarcosine-insoluble method. 5 Briefly, the obtained bacterial pellets were resuspended in PBS buffer and lysed by sonication. The samples were then centrifuged at 5,000 g for 20 min to remove unbroken cells and cellular debris. The supernatants were collected and further centrifuged at 100,000 g for 1 hr at 4°C by using an Optima LE-80 K ultracentrifuge (Beckman, Palo Alto, CA). Next, the supernatants were discarded and the pellets were dissolved in 2% (w/v) sodium lauroyl sarcosinate (Sigma-Aldrich, St. Louis, MO) for 30 min at room temperature. The samples were then ultra-centrifuged again under the same conditions. We determined protein concentrations by using a Bradford assay.
Sample preparation and TMT labeling
On the basis of the filter-aided sample preparation method, we reduced 50 μg of protein from each sample with dithiothreitol, alkylated with iodoacetamide, and digested with trypsin at a ratio of 1:20 for 12–16 hr. 13 The digested peptides from each sample were labeled by using TMT isobaric and isotopic mass-tagging kits (Thermo Fisher, San Jose, CA) according to the manufacturer's protocols. The labeling scheme included A. hydrophila without OXY treatment labeled as TMT126, 5 μg/ml OXY as TMT127, and 10 μg/ml OXY as TMT128. The labeled peptides were pooled and desalted by using C18 SPE cartridges (Waters, Inc., Milford, MA) and dried by using a CentriVap Concentrator (Labconco, Inc., Kansas City, MO).
LC MS/MS analysis
We dissolved dried peptides in 0.1% formic acid and then quantitatively analyzed the peptides with the AB/Sciex TripleTOF 5600+ system (AB SCIEX, Concord, Canada) coupled with the nanoAcquity ultraperformance liquid chromatography system (Waters, Inc.). Peptides were loaded into a reverse-phase Symmetry C18 trap column (180 μm × 20 mm; Waters, Inc.) at a flow rate of 3 μl/min for 5 min and then separated in a 1.7 μm BEH130 C18 analytical column (100 μm × 100 mm; Waters, Inc.) at a flow rate of 300 nL/min for 90 min. Next, the obtained eluent from the BEH130 C18 column was analyzed by using the AB SCIEX TripleTOF 5600+ mass spectrometry (MS) system. We determined protein identification and quantification by using Scaffold 4.3.4 (Proteome Software, Portland, OR) and by searching the A. hydrophila ATCC7966 database. Proteins were identified based on the criterion of at least two matched peptides and a false discovery rate <1%. Further, the TMT average reporter-ion ratio had to be ≥1.5 (increased) or ≤0.67 (decreased) to be considered a significant change. We performed each experiment with at least three technical replicates and normalized samples with related control peptides.
Bioinformatics analysis
We analyzed overlapping upregulated and downregulated proteins among the two OXY treatments by using an online Venn diagram tool (www.stefanjol.nl/venny). We performed gene ontology (GO) enrichment of the differentially expressed proteins by using the online software OmicsBean (www.omicsbean.com). 14 We performed protein subcellular location predictions by using Gneg-mPLoc v. 2.0 (www.csbio.sjtu.edu.cn/bioinf/Gneg-multi/#). 15
Proteomics validation via quantitative real-time PCR
We extracted A. hydrophila total RNA from the three experimental groups by using RNAiso Plus reagent (Takara Bio, Tokyo, Japan). cDNA was synthesized by using 1 μg of isolated RNA per sample and a PrimeScript TM RT reagent kit with gDNA Eraser (Takara Shuzo, Otsu, Japan) according to the manufacturer's protocols. We examined the expression levels of 12 proteins (Supplementary Table S1; Supplementary Data are available online at www.liebertpub.com/mdr), with glyceraldehyde-3-phosphate dehydrogenase (GAP-1) expression used as an internal control (forward: 5′-AGAGCCTCAATGCCTATCTGC-3′ and reverse: 5′-ACCCGAACTCGTTGTCAT ACC-3′). 16 We used SYBR® Premix Ex Taq™ II (Tli RNase H Plus; Takara Shuzo) for all of the quantitative real-time PCRs (qPCRs) under the following conditions: 5 sec at 95°C, 34 sec at 60°C, and finally 40 cycles of 30 sec at 95°C, with a melting curve added. We performed all experiments with at least three independent replicates.
Western blotting
We performed Western blotting as previously described, with proteins separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and subsequently transferred to a PVDF membrane. 17 The transfer was performed for 15 min at 25 V in 10 × Transfer buffer (Bio-Rad, Hercules, CA) by using the Trans-Blot Turbo Transfer System (Bio-Rad). The membrane was blocked with 5% milk in PBS containing 0.05% Tween-20 (PBST) for 1 hr at room temperature, and it was then incubated for 1 hr at room temperature with a primary antibody (1:4,000) targeting one of two proteins (A0KHH2 and A0KHS1). The membrane was then washed three times with PBST, incubated with horseradish peroxidase-conjugated goat anti-mouse secondary antibody (1:4,000) for 1 hr at room temperature, and finally subsequently rinsed three times. The positive bands were detected by using Clarity Western ECL substrate and imaged by using the ChemiDoc XRS+ System (Bio-Rad). We used a PVDF membrane stained with Coomassie Brilliant Blue R350 as a loading control. 18
Results
Quantitative TMT labeling analysis
In agreement with a previous study, A. hydrophila exposed to 5 or 10 μg/ml OXY showed significantly inhibited growth. 12 To investigate the roles of OMPs during antibiotic stress, proteins were extracted by using the sarcosine-insoluble method. Protein samples from each group were then separated by SDS-PAGE, with the visible bands showing apparent differences between the two treatments and the control (Fig. 1A). The samples were then prepared for high-resolution liquid chromatography-mass spectrometry (LC MS/MS), with the samples trypsin digested, TMT labeled, and pooled. We performed two independent biological replicates per experimental group.

Comparative analysis of OM fractions from Aeromonas hydrophila ATCC 7966 under OXY stress.
In this study, a total of 259 proteins derived from 1,960 peptides were identified by TMT labeling-based LC MS/MS. Relative to the control, 125 (33 down- and 92 upregulated) proteins in the 5 μg/ml OXY treatment group and 150 (28 down- and 122 upregulated) in the 10 μg/ml OXY treatment group were significantly differentially expressed, with a TMT average reporter-ion ratio having to be ≥1.5 (increased) or ≤0.67 (decreased) (Table 1). Of the identified proteins, 16% (5 μg/ml group) and 14% (10 μg/ml group) were OMPs according to the predicted subcellular locations. Among the identified differentially expressed proteins, an overlap between the 5 and 10 μg/ml OXY treatment groups was found between 23 downregulated proteins and 89 upregulated (Fig. 1B). In this study, we analyzed the correlation of protein ratios (log2 of TMT ratios) between the two independent biological replicates for a given treatment group. The 5 μg/ml OXY treatment group had a regression coefficient (R2) of 0.900 (Supplementary Fig. S1A), and the 10 μg/ml OXY group had an R2 = 0.988 (Supplementary Fig. S1B), which shows high reproducibility. In addition, we also compared the correlation of protein ratios (log2 of average ratios) between the 5 and 10 μg/ml OXY treatments (R2 = 0.846) and indicated that similar biological behaviors were present in response to the different OXY dosages (Fig. 1C).
CK means the control that A. hydrophila treated without OXY; 5 and 10 mean the 5 and 10 μg/ml OXY treatment groups, respectively. OXY, oxytetracycline.
Functional analysis of OXY stress-induced expression
We performed functional analysis of the identified differentially expressed proteins by using GO analysis. In the group treated with 5 μg/ml OXY (Fig. 2), the upregulated proteins were mainly annotated to several biological processes, including biosynthetic processes, protein metabolic process, and translation. Many of the downregulated proteins were involved in transport processes, such as maltodextrin transport (3%), maltose transport (3%), oligosaccharide transport (3%), energy-coupled transmembrane transport (3%), ATP hydrolysis-coupled proton transport (3%), and polysaccharide localization (3%). In the molecular function category, some of the upregulated functions corresponded to the translational process, such as structural constituent of ribosome (15%), rRNA binding (15%), RNA binding (11%), nucleic acid binding (11%), and tRNA binding (2%). Further, the downregulated expression related to transporters, to include decreases in maltodextrin transmembrane transporter activity, maltose transporting porin activity, maltose transmembrane transporter activity, and multiple saccharide transmembrane transporter activities. These results indicate that there are prominent differences in the associated biological process and molecular function between the upregulated and downregulated proteins during OXY stress. Interestingly, the finding that translation-related proteins were upregulated and transporter-related proteins were downregulated in response to tetracycline antibiotic stress was similar to our previous studies.12,19 The increased translation and translational activities (such as rRNA binding, RNA binding, and tRNA binding) noted after OXY stress would, subsequently, enhance protein synthesis and may serve to repair proteins damaged by OXY exposure. 20 In addition, we found that several transport activities and the OMP LamB were downregulated, thus suggesting that a reduction in these activities may enable survival during antibiotic stress. 21 We also performed GO analysis for the 10 μg/ml OXY treatment group, with similar results noted (Supplementary Fig. S2).
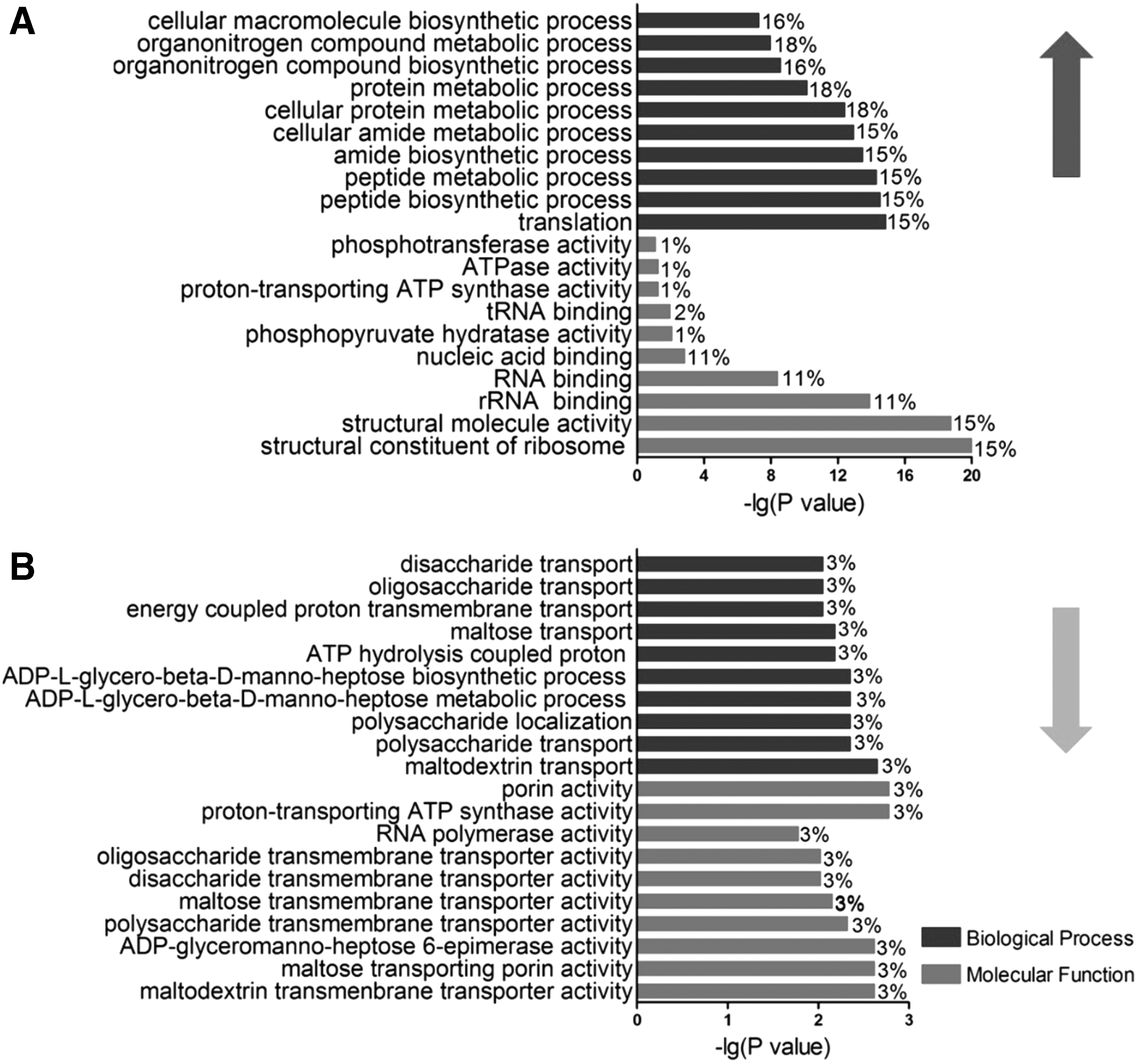
GO analysis of differentially expressed proteins in Aeromonas hydrophila in response to 5 μg/ml OXY treatment. GO analysis of differentially upregulated
KEGG pathway analysis of differentially expressed proteins and cluster analysis of differential OMPs
To further characterize the response of A. hydrophila to OXY stress, pathways were constructed for the identified differential proteins in KEGG (Fig. 3). For the 5 μg/ml OXY treatment group, pathways related to β-lactam resistance and ribosomes were the most enriched for the upregulated proteins, whereas RNA polymerase and purine metabolism-related pathways were enriched for the downregulated proteins. Interestingly, oxidative phosphorylation was associated with both the up- and downregulated proteins, with 70% of the associated proteins upregulated. Moreover, the two different OXY dosages had some pathways in common. In the 10 μg/ml OXY treatment group, however, arginine and proline metabolism were downregulated and RNA degradation was upregulated (Supplementary Fig. S1B). In addition, we performed hierarchical clustering of the 27 altered OMPs to generate a dendrogram and heat map, with most of the OMPs clustered based on expression level (Fig. 4).

Distribution percentages of differential proteins in the 5 μg/ml OXY treatment groups using KEGG pathway enrichment. Color bars indicate downregulated (gray) and upregulated (black) proteins. The number in the bar represents the number of enriched proteins.

Hierarchical clustering of differentially expressed OMPs responds to OXY stress. A heat map was constructed to visualize the differential proteins in the 5 and 10 μg/ml OXY treatment groups. The columns correspond to two biological replicates for each dosage. (columns 1 and 2: the log2 ratio of bacteria under 10 μg/ml OXY stress/control and columns 3 and 4: the log2 ratio of bacteria under 5 μg/ml OXY stress/control). CK means the control that Aeromonas hydrophila treated without OXY; R1 and R2 mean biological replicate 1 and 2, respectively. OMPs, outer membrane proteins.
Validation of several altered proteins via qPCR
We selected a subset of the identified differentially expressed proteins for further validation via qPCR. The selected targets were predicted to be located in the OM and included nine OMP genes: AHA_1280 (protein name: A0KHS0), AHA_1281 (A0KHS1), AHA_1447 (A0KI84, BamE), AHA_1861 (A0KJE1), AHA_2766 (A0KLX3), AHA_3953 (A0KQ36), AHA_0854 (A0KGK5), AHA_1182 (A0KHH2), and AHA_3433 (A0KNS1); one lipoprotein: AHA_1740 (A0KJ25); and one maltoporin transporter gene: AHA_1166 (LamB; Fig. 5). The qPCR results for AHA_1280 (protein name A0KHS0), AHA_1281 (A0KHS1), AHA_1447 (BamE), AHA_1740 (A0KJ25), AHA_1861 (A0KJE1), and AHA_2766 (A0KLX3) were consistent with the proteomic results for both OXY treatment groups. However, AHA_3953 (A0KQ36) levels for the 5 μg/ml OXY treatment were higher than the protein levels, although no significant difference was noted for the 10 μg/ml OXY treatment. For AHA_1166 (A0KHF6, LamB), levels were slightly increased in the 5 μg/ml OXY group and decreased in the 10 μg/ml OXY group; whereas at the protein level, both dosages showed a significant decrease. Interestingly, the mRNA expression levels of AHA_0854 (A0KGK5), AHA_1182 (A0KHH2), and AHA_3433 (A0KNS1) were completely contrary to the noted protein levels for both dosages.
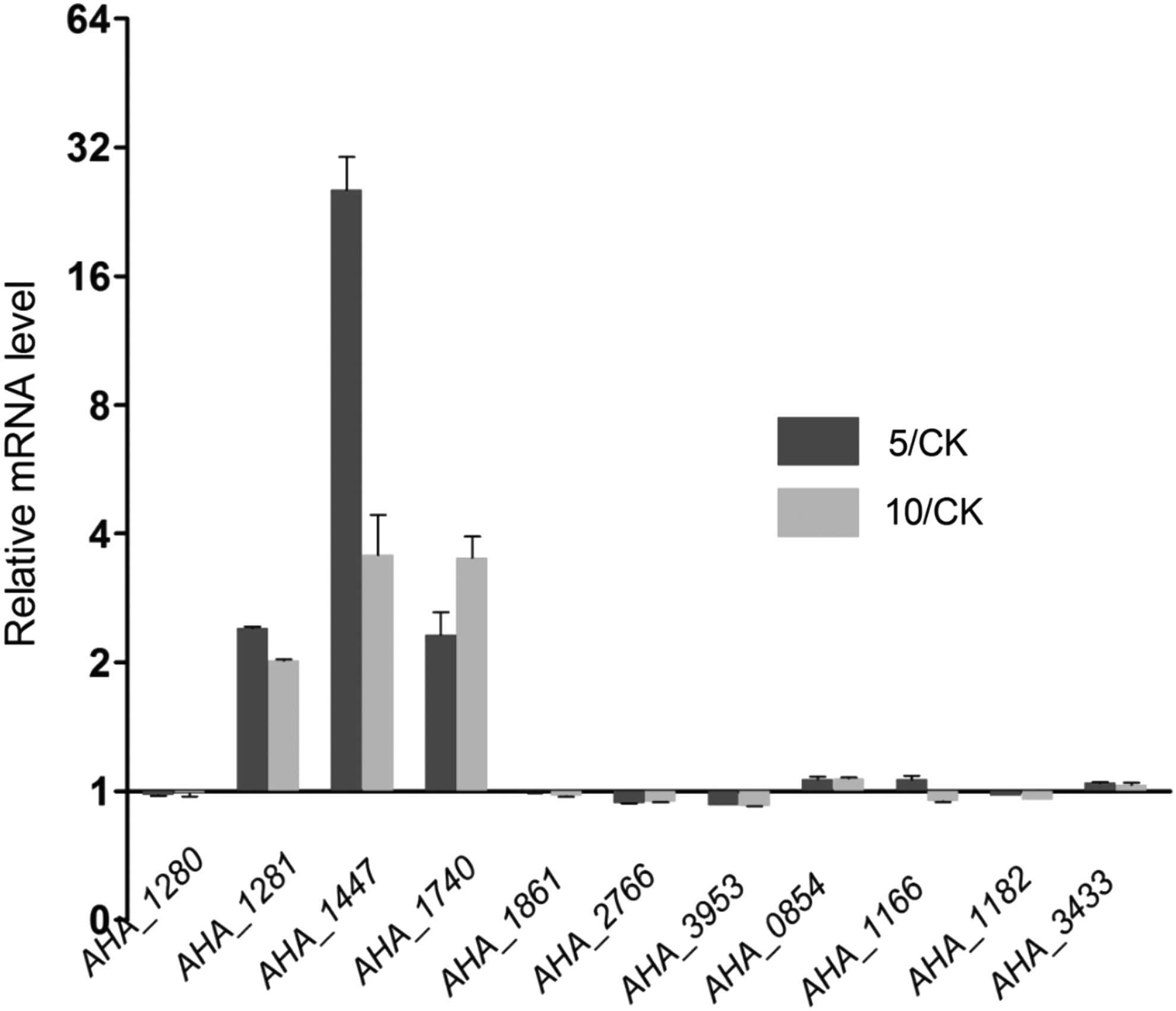
qPCR analysis of a subset of differential OMPs in Aeromonas hydrophila under OXY stress. The mRNA levels of 11 genes were analyzed by qPCR for both OXY dosages, including nine OMP genes: AHA_1280 (protein name: A0KHS0), AHA_1281 (A0KHS1), AHA_1447 (A0KI84, BamE), AHA_1861 (A0KJE1), AHA_2766 (A0KLX3), AHA_3953 (A0KQ36), AHA_0854 (A0KGK5), AHA_1182 (A0KHH2), and AHA_3433 (A0KNS1); one lipoprotein: AHA_1740 (A0KJ25); and one maltoporin transporter gene: AHA_1166 (LamB). CK means the control that A. hydrophila treated without OXY; 5 and 10 mean the 5 and 10 μg/ml OXY treatment groups, respectively. qPCR, quantitative real-time PCR.
Validation of proteomic analysis by Western blotting
To further validate the differentially expressed proteins in the 5 and 10 μg/ml OXY treatment groups, two altered OMP proteins (A0KHH2 and A0KHS1) were selected and examined by Western blotting. As shown in Fig. 6, both proteins were increased as the previous quantitative proteomics analysis under both OXY doses treatment.
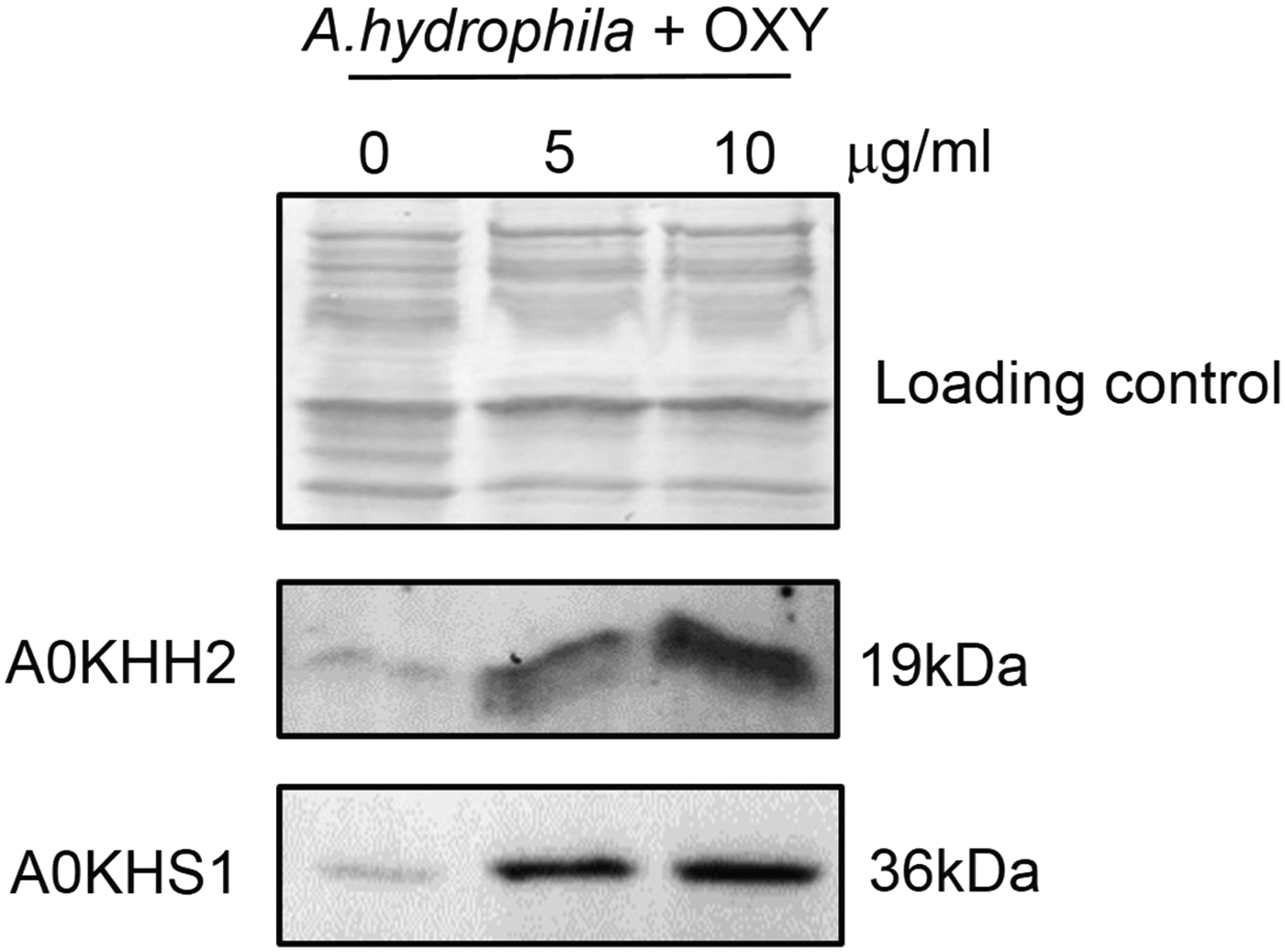
Western blotting validates selected differentially expressed OMPs in Aeromonas hydrophila under OXY stress. Coomassie staining was used as the loading control (upper).
Discussion
For decades, A. hydrophila has posed a great threat to the aquaculture industry, with it causing a fulminant epidemic, bacterial septicemia, and hemolytic ascetic disease. 22 A. hydrophila infections are of particular concern in large-scale fish farming, where antibotics were frequently used and abused for a long time to prevent and treat the bacterial infections so that the development of resistance is a large concern; with OXY and chlortetracycline resistance reported in our previous studies.21,23 Herein, we performed an OXY-susceptibility stress assay with samples exposed to either 5 or 10 μg/ml OXY. To further characterize the bacterial response to antibiotic stress, we identified differentially expressed sarcosine-insoluble proteins by using TMT labeling-based MS. Proteomic analysis displayed that many sarcosine-insoluble proteins were differentially expressed for both OXY treatment groups. The following GO categories of altered proteins demonstrated that translation processes, such as structural constituent of ribosome, RNA binding, and rRNA binding, were significantly enriched in increasing abundance. This result was consistent with our previous studies about fitness mechanisms of A. hydrophila based on proteomics profiles using iTRAQ labeling and SWATH-MS methods under antibiotics stresses. 12 In addition, we also founded that translation-related proteins displayed increasing abundance when A. hydrophila exhibited fitness and acquired resistance to chlortetracycline in biofilm. 21 Therefore, the enrichment of the translation process may be a global resistant strategy against antibiotics. However, the expression of transport-related proteins and some OMP proteins, such as A0KLX3, A0KHS0, A0KNS1 and LamB, were downregulated under antibiotic conditions, which should be correlated to the antibiotic uptake and accumulation in bacteria. 8
Many of the OMPs had altered expression levels in response to OXY stress, with many of them downregulated at both the OXY concentrations. One of these proteins is OmpAII (A0KHS0). OmpA is a major β-barrel porin that is highly conserved among Gram-negative bacteria and contains two homologous proteins OmpAI and OmpAII in A. hydrophila. It is commonly examined when studying OMP structures and folding dynamics, and serves multiple functions in adhesion and invasion, serving an immune target and functioning as a receptor for colicin and bacteriophages.24,25 Meanwhile, several studies have demonstrated that OmpA associates with antibiotic resistance. In Acinetobacter baumannii, the deletion of OmpA can change the susceptibilities to some clinical antimicrobial agents and suggest that OmpA is involved in the intrinsic resistance of this bacteria. 26 Nevertheless, this study found that OmpAII decreased enrichment when bacteria were exposed to a high dose of OXY, which may contribute to increasing the resistance to OXY in A. hydrophila. Xiong et al. reported that OmpA family VPA1186 was downregulated at the level of protein expression in Vibrio alginolyticus when the antibiotic-susceptible strain was exposed to antibiotics, such as tetracycline, kanamycin, and nalidixic acid, which was similar to the earlier mentioned result of this study. 27 In addition, the study, that is adapted tolerance to benzalkonium chloride in E. coli K-12, found that OmpA was downregulated in adapted strains as well. 28 Thus, based on several previous studies, we speculate that the downregulated expression of OmpA may be a particular stress feedback in the adaptive resistance to antibiotics.29,30 But further research will be performed to elucidate the role of OmpA on antibiotic resistance in A. hydrophila.
Besides that, interestingly, the maltoprotein (LamB) was downregulated under OXY stress. This OMP that is involved in the transport of maltose sugar, disaccharide/oligosaccharide, and transmembrane transporter activities decreases due to its downregulation under OXY stress. Further, according to previous studies, LamB as a porin channel of OM is a negative regulator to various antibiotics through decreasing its abundance in several bacterial species, which results in decreasing the uptake of antibiotics.7,31 Thus, knockout of this gene in E. coli has a critical impact on resistance to various classes of antibiotics, including OXY, chlortetracycline, and tertracycline. 32 Our results presented herein show that OXY-induced LamB downregulation warrants further examination and may provide a novel strategy to combat A. hydrophila.
In this study, BamB (A0KJ47), BamC (A0KGP4), and BamE (A0KI84), which are OMP assembly factors and belong to the BAM complex, were significantly increased under OXY stress. This complex exerts significant impact on the protein folding and inserting of nascent OMPs into the OM in bacteria with two other proteins BamA and BamD. 33 The previous reports displayed that the depletion of these three genes (bamB, bamC, and bamE) exhibited the defects of OMPs folding and a decreasing barrier function of OM permeability, resulting in increasing sensitivity to antibiotics.34,35 In this study, the upregulated expression of these proteins contributed to elevating the OM permeability barrier function so that A. hydrophila built up high tolerance to OXY stress.
Conclusions
In summary, this study examined the effect of OXY stress on A. hydrophila using quantitative proteomics, with proteins extracted by using a sarcosine-insoluble method. The results demonstrated that biological processes such as translation and transporter process play a key role in antibiotic stress. Further, many of the downregulated proteins were predicted to be OMPs and were further validated by qPCR and Western blot analyses. Thus, this study has further extended our understanding of the functions of OMPs in response to antibiotic stress.
Footnotes
Acknowledgments
This work was sponsored by grants from the NSFC (Nos. 31200105, 31670129, and 31470238), the Fujian Agricultural and Forestry University Foundation for Distinguished Young Scholars (No. XJQ201201), and the Education Department of Fujian Province for Youths (No. JAT160171).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.