
Abstract
Aim:
This work studied the impact of the quorum-sensing molecule, farnesol (FAR), on fluconazole (FLC)-resistant Candida albicans isolate CY 1123 compared with the susceptible standard strain C. albicans SC5314. The genes encoding efflux pumps belonging to the ATP-binding cassette (ABC) and major facilitator superfamilies, together with overexpression or point mutation of the ERG11 gene, are the main resistance mechanisms to azole antifungal drugs.
Results:
The upregulation of genes coding for CDR1, CDR2, and MDR1 were confirmed by qPCR with respect to the housekeeping gene ACT1 in the resistant strain. The contribution of the ERG11 gene was also observed. Markedly, increased pump activity (Cdr1 and/or Cdr2) in the CY 1123 strain was confirmed using diS-C3(3) assay. However, the addition of FAR to the yeasts diminished the difference in staining levels between the SC5314 and CY 1123 strains, demonstrating the concentration-dependent character that could be caused by an effective modulation of Cdr pumps. FAR (60 and 100 μM) was also able to decrease the minimal inhibitory concentrations (MIC50), denoting the inhibition of planktonic cells by 50%, from 8 to 4 μg/mL of FLC when the resistant strain CY 1123 was not cultivated with FLC. However, when it was exposed to 64 μg/mL of FLC, the MIC50 shifted from 64 to 8 μg/mL.
Conclusion:
Besides the many other effects of FAR on eukaryotic and prokaryotic cells, it also affects ABC efflux transporters, resulting in changes in resistance to azoles in C. albicans isolates. However, this effect is dependent on FAR concentrations.
Introduction
C
The latter mechanism involves the enhanced activity of ATP-binding cassette (ABC) and major facilitator superfamily (MFS) proteins.3,4,7,8 In C. albicans, the overexpression of Candida drug resistance protein 1 (Cdr1p), which belongs to the ABC superfamily, is one of the most prominent contributors to multidrug resistance.9–11 Of the MFS transporters, only CaMDR1p is known to extrude drugs, while its overexpression is predominantly associated with azole resistance. 3
Several approaches have been proposed to tackle efflux-mediated antifungal drug resistance.8,12 The inhibition of efflux pumps by specific compounds is considered to be a feasible strategy to overcome clinical antifungal resistance.7,13 Many traditional antioxidant, anti-inflammatory, and antibacterial agents have been considered to have favorable antifungal potential alone and/or in combination with FLC against FLC-resistant C. albicans.12,14 Li et al. 14 published a study of the synergistic effect of chloroquine combined with FLC not only on FLC-resistant C. albicans and Candida tropicalis, but also on Candida krusei isolates that are known to possess a natural resistance to FLC.
Since farnesol (FAR) is a sesquiterpene alcohol with a natural origin, is part of the essential oil of some plants, for example Zea mays, 15 but is also produced by yeasts. 16 This quorum-sensing molecule is also produced by yeasts of the genus Candida.17,18 It blocks the yeast-to-hyphae transition and has an impact on biofilm formation.19,20 Ramage et al. 19 described an inhibitory effect of 300 μM FAR when added to a preformed 24-h biofilm. Nowadays, FAR has also been examined as a potential antimicrobial agent. It is able to inhibit the growth of some microorganisms, such as Paracoccidioides brasiliensis 21 or dermatophytes, 22 but its mechanism of action has not been fully elucidated yet. In the research of Katragkou et al., 23 in vitro interactions between FAR and the drugs FLC, amphotericin B (AMB), or micafungin against C. albicans biofilms were observed. 23 It was also confirmed that the addition of FAR reduced the minimal inhibitory concentrations [MIC] of azoles (FLC and voriconazole) and AMB on free-living planktonic cells.24,25 Testing FAR in combination with FLC on FLC-resistant strains resulted in a reversal of FLC resistance, indicating a potential application of FAR as an adjuvant therapeutic agent.24,26 Both of the above reports address the topic, but in particular Sharma and Prasad 24 was a pivotal study, proving not only the effect of FAR in combination with FLC on FLC-resistant C. albicans, but also considering FAR to be a modulator of the efflux mediated by ABC transporters. However, it is worth noting that research on the interaction between FAR and multidrug transporters has still raised several questions, for example, whether FAR inhibits efflux pumps directly or modulate Cdr1p or Cdr2p expression, and whether this process is dependent on FAR concentration.
The above study of Sharma and Prasad 24 presented experiments with a C. albicans clinical isolate with resistance to FLC determined only by MIC, and efflux was measured by rhodamine assay. Cordeiro et al. 25 also reported an interaction between antifungals and FAR evaluated by MIC and fractional inhibitory concentration index (FICI), but the mechanisms of resistance to FLC in the tested strains were unknown.
The aim of our research was to study the influence of FAR in combination with FLC on the FLC-susceptible strain C. albicans, compared with the clinical isolate of C. albicans with known FLC resistance mechanisms, as well as to evaluate the interaction between FAR and FLC in terms of azole resistance.
Materials and Methods
Strain selection and susceptibility testing
For these experiments, the standard strain C. albicans SC5314, 27 susceptible to FLC, and a clinical isolate of C. albicans CY 1123, resistant to FLC, (Collection of the Research Center for Pathogenic Fungi and Microbial Toxicoses, Chiba University) were selected. The clinical isolate was not selected randomly, but because of its high resistance to FLC already reported by Tamura et al. 28 This strain was also involved in our previous study dealing with the photodynamic inactivation of the biofilm formed by a FLC-resistant clinical isolate. 29 Both tested strains were preserved at −80°C in 1 mL of YPD broth (Yeast Extract Peptone Dextrose Broth, Biolife, Milan, Italy) supplegmented with 30% sterile glycerol (Centralchem, Bratislava, Slovakia). The storage broth for CY 1123 and SC5314 contained 64 and 0.25 μg/mL of FLC (Mycomax; Zentiva, Prague, Czech Republic), respectively. Before each experiment, strains were revived on YPD plates.
Susceptibility testing was determined by the broth microdilution method according to the protocol of EUCAST (European Committee on Antimicrobial Susceptibility Testing). 30 Briefly, strains were subcultured from the stock sample preserved at −80°C on a YPD agar plate without FLC (C. albicans SC5314) or with the addition of 64 μg/mL of FLC for C. albicans CY 1123 and 0.25 μg/mL for SC5314, respectively. After cultivation at 37°C for 24 hr, the cell suspensions were prepared in RPMI 1640 medium (Roswell Park Memorial Institute Medium; AppliChem, Darmstadt, Germany) supplemented with 2% D-glucose (Centralchem) and buffered with 0.165 M MOPS (morpholinopropane sulfonic acid; Serva, Heidelberg, Germany) to pH 7.0 using a hemocytometer. The final tested concentration of strains was 10 5 cells/mL. The tested concentrations of FLC were prepared in the range from 0.125 to 64 μg/mL. Ninety-six-well flat-bottomed polystyrene microtiter plates (Sarstedt, Nümbrecht, Germany) were filled with 100 μL of the cell suspension (2 × 10 5 cells/mL) and with 100 μL of agents in RPMI 1640 medium. The microtiter plates were incubated at 37°C. After 24 hr, optical density was measured at 570 nm using a plate reader (Dynex MRX-TC Revelation, Denkendorf, Germany). Susceptibility was evaluated in terms of MIC50 (MIC inhibiting the growth of the strain in the presence of the agent by 50% compared with the control sample without antifungal agent), and MIC80 representing the concentration that inhibits growth of FLC-resistant C. albicans by 80% compared with the control sample without agent. Each experiment was repeated at least three times with at least four parallel samples in each experiment.
Determination of possible resistance mechanisms to FLC by qPCR
Total RNA was isolated from C. albicans planktonic cells with an RNeasy Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer's instructions. The RNA Clean and Concentrator™-5 kit (Zymo Research, Irvine, CA) was used for cleaning RNA samples. All RNA samples were treated with DNase I, RNase-free (Thermo Scientific, Waltham, MA) to prevent contamination with genomic DNA. The cDNA of each strain was synthesized using the Maxima First-Strand cDNA Synthesis Kit for RT-qPCR (Thermo Scientific) according to the manufacturer's instructions. The synthesized cDNA samples were stored at −20°C until used. The PCR primers used to amplify and identify the C. albicans CDR1, CDR2, MDR1, ERG11, as well as ACT1 genes are summarized in Table 1. All primers were synthesized by Metabion international AG (Planegg, Germany). The cDNA samples were mixed with Maxima SYBR Green/ROX qPCR Master Mix (2 × ) (Thermo Scientific) and qPCR was performed using a 7900HT Fast Real-Time PCR System (Thermo Scientific). The cycling conditions were as follows: 1 cycle of 10 min at 95°C; followed by 40 cycles of 15 sec at 95°C, and 1 min at 58°C for all genes. After amplification, a melting curve was analyzed to ensure the absence of primer dimers; the dissociation cycle was 15 sec at 95°C, 15 sec at 58°C, and 15 sec at 95°C. The level of gene expression was calculated using the 2−ΔΔCT method with respect to the housekeeping gene ACT1. Samples were compared with the control, which was represented by the standard strain C. albicans SC5314 cultivated without FLC and normalized to 1. Each experiment was repeated at least three times with three parallel samples in each experiment.
List of Nucleotide Sequences of Used Primers
Primers for CDR1, CDR2, and MDR1 genes were used according to Yu et al. 20
Primers for the gene ERG11 was designed by Marek Korenčák (this work).
Primers for the gene ACT1 was designed by Imrich Hikkel (this work).
Fluorescence diS-C3(3) diagnostic assay for monitoring MDR pump activity
DiS-C3(3) fluorescence assay is very suitable for monitoring the activity of the MDR pumps, for which the fluorescent probe diS-C3(3) (3,3′-dipropylthiacarboxycyanine; Fluka, Prague, Czech Republic) is a substrate, such as the Saccharomyces cerevisiae pumps Pdr5p and Snq2p31,32 or Cdr1p and Cdr2p in C. albicans. 33 In the absence of an isogenic pump-deficient strain (taken as a negative control), the determination of pump activity is based on comparing the probe accumulation in cells in the presence and absence of an inhibitor, such as beauvericin (Cayman Chemical Company, Tallinn, Estonia).
After incubation for 16 hr in YPD broth at 37°C, SC5314 and CY 1123 cells were harvested, washed twice with double-distilled water, and resuspended in citrate-phosphate buffer of pH 6.0 to OD578 = 0.1 (citric acid and Na2HPO4. 12 H2O; Lachema, Neratovice, Czech Republic). The potentiometric fluorescent probe diS-C3(3) (10−5 M stock solution in ethanol) was added to 3 mL of yeast cell suspension to a final concentration of 2 × 10−8 M and fluorescence emission spectra of the cell suspensions were measured every 2–5 min in a FluoroMax—4 spectrofluorimeter (Horiba JobinYvon, Edison, NJ) equipped with a xenon lamp. The excitation wavelength was 531 nm, fluorescence range 560–590 nm, duration of one spectral scan 20 sec, and scattered light was eliminated with an orange glass filter with a cutoff wavelength at 540 nm.
The rate and extent of intracellular probe accumulation, the so-called staining curve, records the dependence of the fluorescence emission maximum wavelength λmax on the duration of staining.34,35 When appropriate, beauvericin, an effective inhibitor of both the Cdr1p and Cdr2p pumps, 33 was added 10 min before the addition of the diS-C3(3). The samples were kept at room temperature and occasionally gently stirred. The Cdr1 and Cdr2 pump activity of the standard and FLC-resistant C. albicans strains was determined at four different times (t = 30, 60, 90, and 120 min) from the staining curves of both strains in the presence and absence of 10 μM beauvericin. The measure of pumps' activity, Δλmax/t (nm), was obtained by subtracting the λmax/t value for the strain in the absence of beauvericin from the corresponding λmax/t value of the same strain with pumps inhibited by beauvericin.
When appropriate, the tested FAR was added to the desired final concentration after 11 min of staining. The extent of FAR-mediated probe transport inhibition was calculated from the following equation:
where A and B denote the difference between the equilibrium λmax values of yeasts of both strains in the presence and absence of FAR, respectively.
Susceptibility testing FAR alone and in combination with FLC
Susceptibility testing was performed according to the EUCAST protocol, as was mentioned above. The stock solution of FAR (75 mM, Sigma-Aldrich, Steinheim, Germany) prepared in 96% ethanol (Centralchem) was diluted to the following concentrations: 1 mM, 500, 400, 300, 200, 100, 80, 60, 50, and 30 μM (corresponding to 222, 111, 89, 67, 44, 22, 18, 13, 11, and 7 μg/mL). The effectiveness of FAR was determined for both strains in terms of MIC50, as was previously described for azoles. For determining the impact of FAR in combination with FLC, two concentrations of FAR were tested (60 and 100 μM). FLC was prepared in twofold serial dilutions with final concentrations from 0.125 to 64 μg/mL. Microtiter plates were filled with 100 μL of the cell suspension (2 × 10 5 cells/mL), 50 μL of FLC, and 50 μL of FAR. The microtiter plates were incubated at 37°C for 24 hr and then optical density was measured using a reader. The results were evaluated in terms of MIC50 for the combination of FAR with FLC. Each experiment was repeated at least three times with at least four parallel samples in the experiment. Interaction between FAR and FLC was also evaluated using FICI that was calculated as the sum of concentration of FAR tested in combination with FLC/MIC50 of FAR alone and MIC50 of FLC in combination with FAR/MIC50 of FLC alone. The FICI index was evaluated according to the following criteria: FICI of <0.5, synergy; FICI of ≥0.5 and <1, partial synergy; FICI of 1, additive; FICI of ≥2 and <4, indifferent; and FICI of >4, antagonism.36,37
Statistical analysis
Values are mean from three separate experiments. Bars are standard deviations. The statistical comparison between samples was performed using one-way analysis of variance (ANOVA; free online software in-silico.net). Differences were considered statistically significant at p < 0.05 (*), strongly significant at p < 0.01 (**), and extremely significant at p < 0.001 (***).
Results
Susceptibility testing
The standard strain C. albicans SC5314 was susceptible to FLC with an MIC50 of 0.5 μg/mL. The MIC50 for the FLC-resistant strain was determined to be 8 μg/mL, whereas the MIC80 was 64 μg/mL. Thus, this strain was preserved in parallel on a plate without FLC and with a concentration of 64 μg/mL. The MIC50 of the strain cultivated with and without the presence of FLC was determined to be 8 and 64 μg/mL, respectively.
Determination of changes in expression of CDR1, CDR2, MDR1, and ERG11 genes by qPCR
According to the MIC results, a subinhibitory concentration of FLC (0.25 μg/mL) was selected for C. albicans SC5314 and a concentration of 64 μg/mL was used for the FLC-resistant strain, CY 1123. These concentrations were selected for the cultivation of yeasts subjected to qPCR analysis. Figure 1 summarizes the results from qPCR expressing the relative change in the regulation of the selected genes with respect to the housekeeping gene ACT1. The results showed a slight upregulation of the CDR1 and CDR2 genes for C. albicans SC5314 in the presence of FLC during overnight cultivation. However, the FLC-resistant strain expressed a markedly increased expression of the same genes compared with the control sample of CY 1123 without FLC. The MDR1 gene was only slightly upregulated in C. albicans SC5314, whereas the upregulation was markedly higher in the CY 1123 strain. On the other hand, this gene was downregulated in the CY 1123 strain cultivated without FLC. The final observed gene was ERG11, which also exhibited upregulation in both strains in the presence of FLC, but was downregulated in the CY 1123 strain cultivated without FLC.

Relative change in expression of genes: CDR1
Overexpression of Cdr1p and Cdr2p is associated with increased efflux of fluorescent substrate from cells
To test the increased efflux activity of Cdr1p and Cdr2p in CY 1123 cells relative to the susceptible standard strain SC5314, their fluorescent substrate diS-C3(3) was used. 33 The diS-C3(3) assay confirmed a significantly higher activity of the Cdr1 and Cdr2 pumps in cells of the CY 1123 strain, which persisted even 2 hr after the addition of the probe, compared with their low activity in the standard susceptible strain, C. albicans SC5314 (Fig. 2).

High and stable Cdr1 and Cdr2 pump activity in FLC-resistant strain C. albicans CY 1123 compared with standard strain C. albicans SC5314 detected by their benchmark pump substrate–fluorescent probe diS-C3(3). Staining curves of the standard strain C. albicans SC5314
FAR effectively reduces diS-C3(3) efflux from the cells
To monitor the effect of FAR on the Cdr1p- and Cdr2p-mediated efflux of the probe diS-C3(3), the compound was added to the stained cells of both strains at a concentration range of 20–100 μM (Fig. 3A, B). The addition of FAR to cells diminishes the difference in staining levels between the strains of SC5314 and CY 1123, demonstrating the concentration-dependent character of the probe transport inhibition. The data indicate that FAR reduced the activity of both Cdr pumps, resulting in a decreased MIC50 to FLC in the FLC-resistant strain CY 1123.
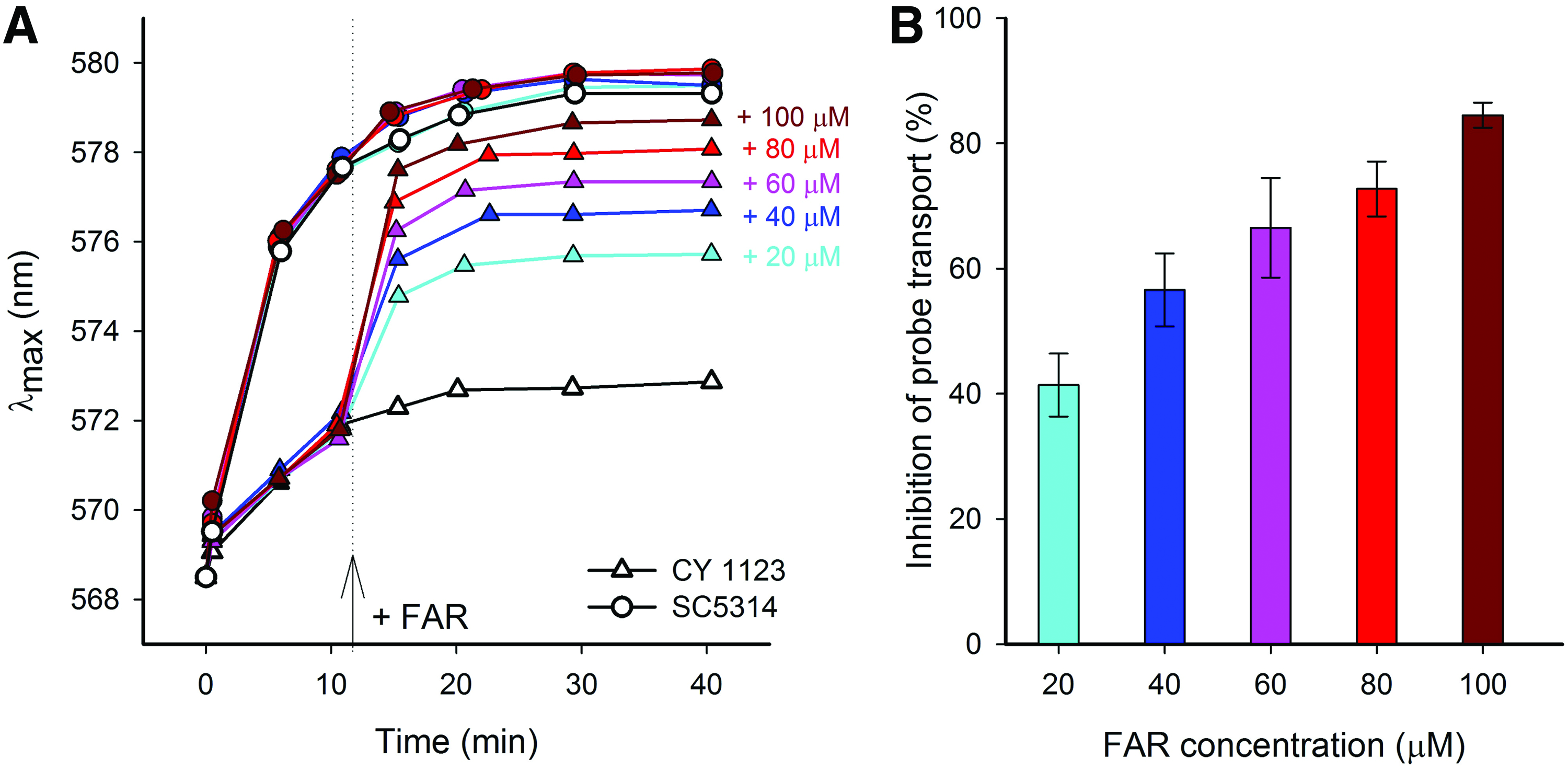
FAR is an effective probe transport inhibitor of Cdr1 and Cdr2 pumps.
Effectiveness of FAR alone and in combination with FLC
The susceptibility of these C. albicans strains to FAR alone was tested for nine different concentrations. The concentration which reached 50% inhibition was 1 mM, but for testing the combination with FLC, two concentrations of FAR (60 and 100 μM) were used (Table 2). In the FLC-susceptible strain SC5314, FAR did not reduce MIC values, and the FICI was unchanged. Changes in MICs were observed in the FLC-resistant strain CY 1123, the MIC50 for FLC decreased from 8 to 4 μg/mL in the presence of 100 μM FAR (FICI = 0.6) when the strain was not initially exposed to FLC. The lower concentration of FAR was less efficient (FICI = 1.06). However, a markedly reduced MIC50 was determined for CY 1123 in the presence of both tested concentrations of FAR (from 64 to 8 μg/mL; FICI = 0.185; synergic effect) when this strain was previously exposed to 64 μg/mL of FLC.
Effectiveness of Fluconazole Alone and in Combination with 60 and 100 μM Farnesol on Candida albicans Strains Determined According to Minimal Inhibitory Concentrations50 Values
C. albicans SC5314 and C. albicans CY 1123 were cultivated in the presence of 0.25 and 64 μg/mL of FLC, respectively. Bold values exhibited a shift in MIC50.
FAR, farnesol; FLC, fluconazole.
Discussion
FLC is used extensively for chemoprophylaxis and the treatment of systemic fungal infections because of its favorable oral bioavailability and safety profile.38,39 Subsequent FLC resistance has been described in a high percentage of AIDS patients.40,41 Several major mechanisms leading to azole resistance have been elucidated: reduced drug intracellular accumulation, decreased target affinity for the drug, and counteraction of the drug's effect.4,9,39 The latter mechanism includes an upregulation of the ERG11 gene leading to an intracellular increase in the target protein or change in other genes involved in sterol metabolism. However, the most common mechanism is connected with ABC transporters or with MFS, responsible for decreasing the intracellular concentration of azoles. 9 Modulation of these transporters could be a possible way to revert resistant isolates to a susceptible mode.7,25
In this work, a FLC-resistant clinical isolate of C. albicans was studied. The first goal was to explore a potential mechanism of azole resistance. In general, the upregulation of the CDR1 and CDR2 genes after previous exposure to FLC is a well-known phenomenon in clinical isolates.10,42 As expected, only weak upregulation of the CDR1, CDR2, and MDR1 genes was observed in the standard strain. However, the FLC-resistant strain after cultivation in the presence of FLC (64 μg/mL) exhibited a markedly increased expression of the same genes compared with cultivation without FLC. While the upregulation of the CDR1 gene was expected, in agreement with the previous observations of many authors, a high expression of CDR2 occurs less frequently.10,42 Moreover, the expression of this efflux transporter seemed to be constitutively upregulated, as an increased regulation was also observed in this strain without cultivation in the presence of FLC. The additive activities of Cdr1p and Cdr2p against FLC have already been demonstrated, suggesting that the concomitant overexpression of CDR1 and CDR2 genes in azole-resistant C. albicans strains could strengthen their resistance.42,43
Another gene, MDR1, was downregulated in these strains cultivated without FLC, but its expression increased in the presence of FLC. Thus, the activity of all efflux transporters was examined in the studied strain. Another assumed mechanism was also observed–overexpression of the ERG11 gene. While the SC5314 strain was sensitive to a higher concentration of FLC, overproduction of the lanosterol 14α-demethylase or point mutation of the ERG11 gene was not expected. However, in the CY 1123 strain, a contribution of this resistance mechanism could not be excluded, as the ERG11 gene was upregulated in the presence of a high concentration of FLC compared with the same strain without FLC.
An experiment focused on the measurement of diS-C3(3) efflux confirmed the results obtained with qPCR. DiS-C3(3) is a substrate of the transport proteins Cdr1p and Cdr2p like FLC. While the susceptible strain was only able to exclude a low amount of diS-C3(3), the activity of efflux pumps in the FLC-resistant strain CY 1123 was about 18 times higher than that of the SC5314 strain. It has been shown that FAR significantly inhibits Cdr1p- and Cdr2p-mediated probe transport. The results also suggest that FAR effectively modulates both Cdr transporters, as the inhibition of probe transport through the pumps expressed by cells of the strain CY 1123 was over 85% in the presence of 100 μM FAR (Supplementary Fig. S1). However, the effect of FAR proved to be concentration dependent.
The study of Sharma and Prasad 24 provided the first evidence that FAR is a specific modulator of the efflux mediated by ABC multidrug transporters, such as Cdr1p and Cdr2p of C. albicans and ScPdr5p of S. cerevisiae. They reported that FAR at a nontoxic concentration (100 μM) synergized with FLC, ketoconazole, miconazole, and AMB. Similarly, Cordeiro et al. 25 tested the in vitro susceptibility of 45 resistant clinical isolates of Candida spp. to azoles–FLC and itraconazole, AMB, and caspofungin. They found that FAR improved the activity of traditional antifungals, demonstrated by decreased MICs. In our study, the effect of 100 μM FAR was evident in FLC susceptibility testing, because the combination FAR-FLC reduced the value of MIC50 in the FLC-resistant strain from 8 to 4 μg/mL. Moreover, when the CY 1123 strain was previously exposed to FLC, the reduction in MIC50 was higher; from 64 to 8 μg/mL of FLC. The FICIs for this strain confirmed an effect of FAR, as they were in the range from additive to synergic, depending on the concentrations used. Despite this large reduction in resistance, the strain remained resistant to FLC. This could be due to the activity of the Mdr1 pump that was upregulated not only after exposure to the high concentration of FLC, but also in its absence. In the standard strain, no shift in MIC50 for FAR was observed. These results suggested that FAR is able to effectively decrease resistance to FLC, but in a concentration-dependent manner. However, if C. albicans expresses additional resistance mechanisms to azoles than those associated with the ABC transporters, FAR alone does not eliminate resistance.
In conclusion, FAR seems to be a suitable substrate affecting the function of ABC efflux transporters that could be an attractive option in therapy for preventing the development of resistance to FLC and/or modulating FLC resistance in C. albicans clinical isolates.
Footnotes
Acknowledgments
The authors wish to thank their colleagues, Marek Korenčák and Dr. Imrich Hikkel for the design of the primers. This work was supported by the Slovak Research and Development Agency under contract No. (APVV-15-0347) and by the grant VEGA (1/0537/19), supported by the Ministry of Education, Science, Research, and Sport of the Slovak Republic.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.