
Abstract
Fluoroquinolones (FQs) are the drugs of choice for prophylaxis of bacterial infections in immunocompromised cancer patients. This study aimed to investigate FQ resistance and the prevalence of plasmid-mediated quinolone resistance (PMQR) determinants in 239 Gram-negative isolates collected at a tertiary care cancer hospital in Cairo, Egypt. Disc diffusion and broth microdilution tests showed that 70.7% of the isolates were nonsusceptible to ciprofloxacin (MIC50 = 64 μg/ml). Polymerase chain reaction (PCR) revealed that 53.6% of the isolates carried at least one PMQR determinant, of which 23.4% were susceptible to ciprofloxacin. The most prevalent gene, aac(6′)-Ib-cr, was identified in 36.8% of the isolates, while qnr genes were harbored by 31.0% (qnrS, 24.3%; qnrB, 7.1%, and qnrA, 0.4%). The oqxAB genes were only detected in Klebsiella sp. isolates (92.5%). PMQR determinants were more likely detectable among isolates recovered from pediatric patients than adults (59.3% vs. 43.8%) and were significantly associated with ceftriaxone and gentamicin resistance. A combined genetic analysis using random amplified polymorphic DNA-PCR and enterobacterial repetitive intergenic consensus-PCR showed that most of the qnr-positive isolates were not clonal. Findings of the current study raised concerns about the efficacy of prophylactic use of FQs in cancer patients in our region. It also demonstrates the possible role of PMQR-positive ciprofloxacin-susceptible isolates in the dissemination of resistance to other antimicrobial agents and the urgent need to reconsider the existing FQ breakpoints defined by the Clinical and Laboratory Standards Institute.
Introduction
D
Clinically significant high-level FQ resistance usually arises from mutations in the genes coding the target enzymes; DNA gyrase and topoisomerase IV. Other mutations may occur in regulatory genes that control the expression of membrane porins and efflux pumps, leading to reduced intracellular accumulation of FQs. 5 Being fully synthetic, transferable plasmid-mediated quinolone resistance (PMQR) has not been expected to exist. However, in the late 1990s, the first PMQR mechanism, target protection through the pentapeptide repeat family Qnr protein (QnrA), was described. 5 Since then, a variety of other PMQR mechanisms have been described, including other Qnr proteins (QnrS, QnrB, QnrD, QnrC, QnrVC, and the recently discovered QnrE), enzyme inactivation (AAC(6′)-Ib-cr), and drug efflux (QepA and OqxAB).6–8 PMQR genes do not confer FQ resistance by themselves, but rather facilitate the selection of mutants of higher-level resistance. 5
The efficacy of FQ prophylaxis may be limited by the environmental prevalence of resistance and may be compromised when the Gram-negative bacillary resistance increases beyond 20%. 9 Accordingly, to the best of our knowledge, the current study was the first to address the effectiveness of FQ implementation for prophylaxis against Gram-negative bacterial infections in immunocompromised cancer patients in Egypt. For this purpose, we evaluated the recent FQ resistance trend and dissemination of PMQR determinants in Gram-negative isolates recovered from infected patients attending a major cancer hospital in Egypt.
Materials and Methods
Bacterial isolates
A total of 239 Gram-negative clinical isolates were recovered from different clinical specimens collected from infected cancer patients attending the National Cancer Institute (NCI), Cairo University, in the period from November 2014 to July 2015. The NCI is the largest tertiary cancer hospital in Egypt, drawing patients throughout the country. One hundred and fifty isolates (62.8%) were recovered from pediatric patients, while 89 (37.2%) were recovered from adult patients. The majority of isolates were from blood (n = 137; 57.3%), followed by wound swabs (n = 25; 10.5%), pus (n = 23; 9.6%), sputum (n = 18; 7.5%), stools (n = 11; 4.6%), urine (n = 10; 4.2%), drain (n = 8; 3.3%), bronchoalveolar lavage (n = 4; 1.7%), and chest tubes (n = 3; 1.3%). The collected isolates were identified using conventional microbiological techniques and confirmed by Vitek GN cards (bioMerieux, Marcy l'Etoile, France).
Approval of the study protocol was received from the Ethical Review Boards of of Ain Shams University, October University for Modern Sciences and Arts, and the NCI, Cairo University.
Antimicrobial-susceptibility testing
In vitro susceptibility of all isolates to different quinolones was tested by the disc diffusion method according to the Clinical and Laboratory Standards Institute (CLSI) guidelines. 10 Susceptibilities to other antibiotic classes, such as aminoglycosides, cephalosporins, and carbapenems, were also tested. Commercial discs (Oxoid, United Kingdom) of the following antibiotics (μg/disc) were used: nalidixic acid (30), norfloxacin (10), ciprofloxacin (5), levofloxacin (5), gentamicin (10), amikacin (30), ceftriaxone (5), and imipenem (10). The minimum inhibitory concentrations (MICs) of ciprofloxacin and levofloxacin were determined by the broth microdilution method as described by the CLSI. 10 The reference strain Escherichia coli ATCC 25922 was used for quality control.
Polymerase chain reaction screening of PMQR determinants
All isolates were screened for qnrA, qnrS, qnrB, and aac(6′)-Ib genes by multiplex polymerase chain reaction (PCR). Monoplex PCR was used for screening of the efflux pump coding genes; oqxAB and qepA. Plasmid DNA was extracted by the GeneJET Plasmid Miniprep Kit (Thermo Fisher Scientific, Waltham, MA), according to the manufacturer's instructions. Primers used for PCR reactions and their annealing temperatures are listed in Table 1. The gene variant aac(6′)-Ib-cr was further identified by digestion with the restriction enzyme, BseGI (Thermo Fisher Scientific). PCR reactions were carried out in a TAdvanced thermal cycler (Biometra, Göttingen, Germany) in a 25 μl PCR mixture that contained 12.5 pmoles of each primer, 12.5 μl of MyTaq™ Red Mix, and 1 μl (100 nmole) of template DNA, with incubation at 94°C for 5 min; followed by 35 cycles at 94°C for 30 sec, at annealing temperature for 30 sec, and 72°C for 30 sec, and a final extension at 72°C for 5 min. Amplified PCR products were analyzed by gel electrophoresis in 1.5% (w/v) agarose gel containing ethidium bromide (0.5 μg/ml) and visualized on a UV transilluminator. To confirm PCR product identity, amplicons of various genes from six different isolates were selected for sequencing, including qnrA, qnrB, qnrS, aac(6
Ta, annealing temperature.
ERIC, enterobacterial repetitive intergenic consensus; PCR, polymerase chain reaction; RAPD, random amplified polymorphic DNA.
Molecular typing
To investigate the clonal relationship and dissemination of qnr-positive isolates of the same species, two PCR-based fingerprinting techniques, random amplified polymorphic DNA (RAPD)-PCR and enterobacterial repetitive intergenic consensus (ERIC)-PCR, were used.
Genomic DNA was extracted using the Genomic DNA Purification Kit (Thermo Fisher Scientific) according to the manufacturer's instructions. ERIC-PCR was carried out using the ERIC-2 primer as previously described. 17 For RAPD analysis, preliminary assays were carried out on five RAPD primers: 1247, 1252, 1290, 640, and RAPD4 (Table 1). Upon extensive analysis, the primer 1290 produced the most discriminating fingerprints and was selected for use in amplification as described previously. 16 All strains were processed simultaneously for each reaction, and electrophoresis was performed under identical conditions. PCR products were separated in 1.5% (w/v) agarose gels at 90 V for 3 hr. Amplification patterns of both reactions were analyzed using GelComparII software, version 6.5 (Applied Maths, Kortrijk, Belgium). Similarity clustering analyses were performed using UPGMA (unweighted pair-group method with arithmetic mean) and Dice coefficient with 1.5% tolerance.
Statistical analyses
Data analyses, including descriptive statistics, frequencies, and cross-tabulations, were performed using IBM SPSS Statistics 20.0 software (SPSS, Inc., Chicago, IL). Chi-square and Fisher's exact tests were used for comparisons of categorical variables. All tests of significance were two-tailed, and a value of p < 0.05 was considered statistically significant.
Results
Bacterial isolates
During the surveillance period, a total of 239 Gram-negative isolates were recovered from different clinical specimens collected from infected cancer patients. The clinical isolates were identified as E. coli (n = 95; 39.7%), Klebsiella pneumoniae (n = 82; 34.3%), Acinetobacter baumannii (n = 24; 10%), Pseudomonas aeruginosa (n = 19; 7.9%), and other Gram-negative isolates (n = 19; 7.9%) comprising Enterobacter cloacae (n = 8; 3.3%), Enterobacter aerogenes (n = 5; 2.1%), Klebsiella oxytoca (n = 3; 1.3%), Proteus mirabilis (n = 2; 0.8%), and Citrobacter freundii (n = 1; 0.4%).
Prevalence of FQ resistance and concomitant resistance to other antimicrobial agents
Of all tested isolates, 82.0% were nonsusceptible to at least one quinolone, while 66.9% were resistant to all of them. Among different quinolones, nonsusceptibility to nalidixic acid was the most commonly detected (81.2%), followed by ciprofloxacin (70.7%), norfloxacin (69.5%), and levofloxacin (67.4%). The prevalence of ciprofloxacin (CIP) resistance among different species is illustrated in Figure 1.

Prevalence of CIP resistance among different species. CIP MIC50, MIC of ciprofloxacin at which 50% of the isolates were inhibited; other Enterobacteriaceae species include Enterobacter cloacae, Enterobacter aerogenes, Klebsiella oxytoca, Proteus mirabilis, and Citrobacter freundii. MIC, minimum inhibitory concentration.
MIC values of ciprofloxacin and levofloxacin ranged from 0.125 to 256 μg/ml with MIC50 of 64 and 16 μg/ml, respectively. Low-level resistance to ciprofloxacin (MIC: 0.25–1 μg/ml) was detected in 8.7% of isolates, while 51.4% showed high-level resistance (MIC >32 μg/ml). Unexpectedly, higher prevalence of ciprofloxacin resistance was detected among isolates from pediatric patients than those from adults (74.7% vs. 64.0%, p = 0.08).
Prevalence of PMQR genes
At least one PMQR determinant was detected in 53.6% of the clinical isolates. The aac(6′)-Ib-cr gene was the most frequently detected (36.8%), as shown by restriction analysis of PCR products of aac(6′)-Ib (114). The qnr genes were detected in 31.0% of the isolates. The qnrS gene was more common than qnrB (24.3% vs.7.1%), while only one P. mirabilis isolate was positive for the qnrA gene. The qepA gene was not detected in tested isolates. The prevalence of PMQR genes in different bacterial species is shown in Figure 2.
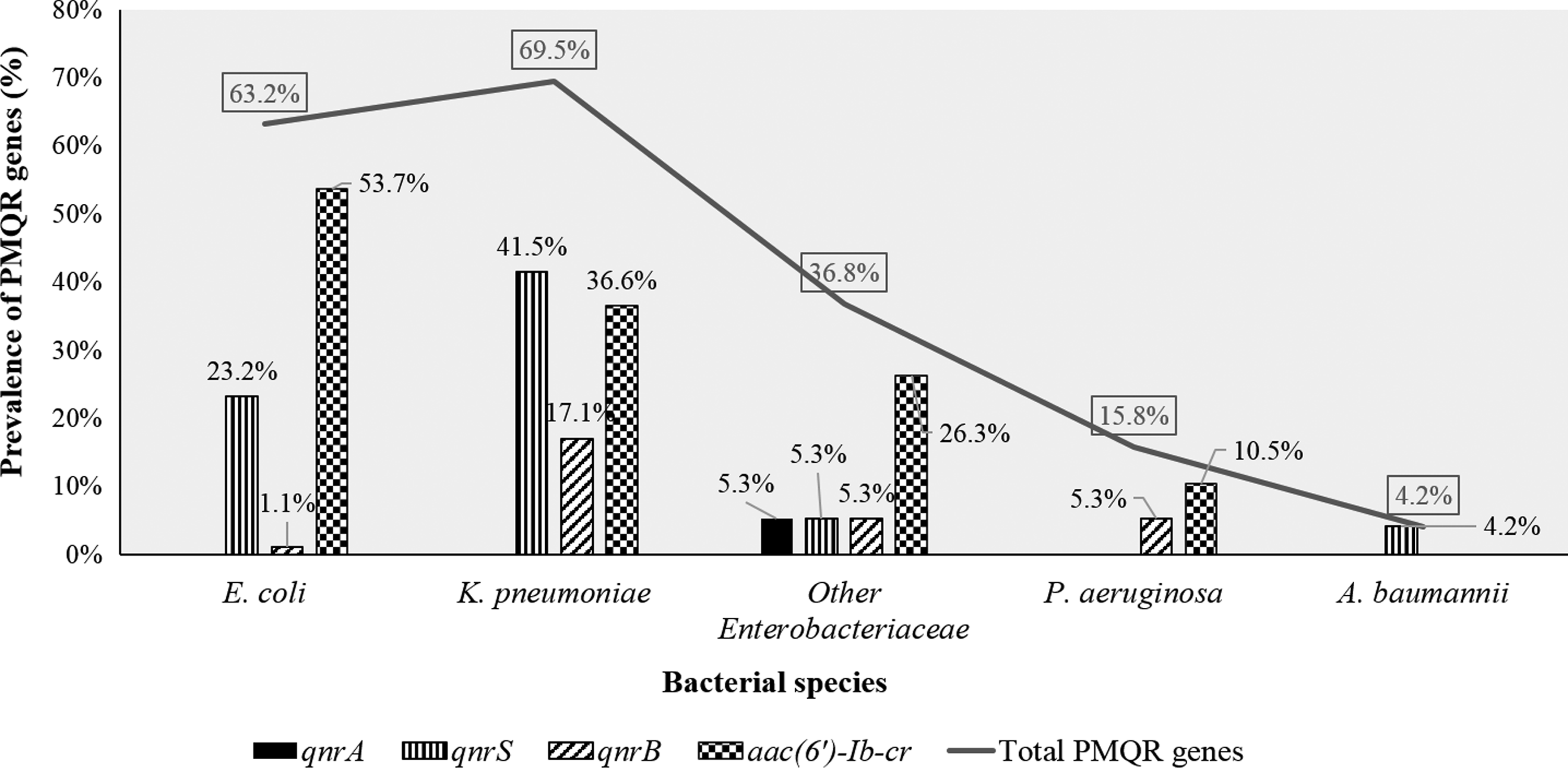
Prevalence of PMQR genes in different bacterial species. PMQR, plasmid-mediated quinolone resistance; other Enterobacteriaceae species include E. cloacae, E. aerogenes, K. oxytoca, P. mirabilis, and C. freundii.
Despite their absence in other species, oqxA and oqxB were detected in 92.5% of Klebsiella sp. isolates (95.1% of K. pneumoniae and 100% of K. oxytoca). Nevertheless, oqxA and oqxB-positive Klebsiella sp. isolates were not considered as PMQR positive and were not a subject for subsequent analyses.
Among PMQR-positive isolates, 32(24.8%) carried two PMQR genes, while combinations of three genes were detectable in 1.6% of isolates. No significant association was found between qnr genes and aac(6′)-Ib-cr, as shown in Table 2.
qnr-positive, isolates positive for any of the qnr genes.
In comparison with isolates recovered from adult patients, a significantly higher PMQR prevalence was found among pediatric patients (43.8% vs. 59.3%, p = 0.02). With respect to specimen types of isolates, PMQR genes were detected in isolates from almost all specimen types, while the majority of PMQR genes (61.7%) were detected in isolates recovered from blood cultures.
Significantly higher ciprofloxacin nonsusceptibility was detected among PMQR-positive isolates than those without PMQR genes (76.6% vs. 64.0%, p = 0.03). Nevertheless, 23.4% of PMQR-positive isolates were susceptible to ciprofloxacin. Furthermore, ciprofloxacin susceptibility was higher among isolates carrying qnr genes (35.0%) than those carrying aac(6′)-Ib-cr (24.1%) as the only PMQR determinants. The MIC50 values of various bacterial species associated with different combinations of PMQR genes are shown in Table 3. Overall, the MIC50 of ciprofloxacin among PMQR-positive isolates was higher than that of isolates without PMQR (64 vs. 32 μg/ml). Notably, PMQR determinants were detected in isolates showing a ciprofloxacin MIC as low as 0.125 μg/ml, that is, 16-fold lower than the cutoff value set by the CLSI for ciprofloxacin-reduced susceptibility (2 μg/ml for intermediate susceptibility and ≥4 μg/ml for full resistance). 10
Percentages were calculated with reference to the number of isolates of each species.
Percentages were calculated with reference to the number of isolates harboring corresponding PMQR genes.
CIPR, ciprofloxacin resistant; CIP MIC50, MIC at which 50% of the isolates were inhibited; other Enterobacteriaceae species include Enterobacter cloacae, Enterobacter aerogenes, Klebsiella oxytoca, Proteus mirabilis, and Citrobacter freundii.
MIC, minimum inhibitory concentration; PMQR, plasmid-mediated quinolone resistance.
PMQR determinants were significantly correlated with ceftriaxone and gentamicin resistance, as revealed by the chi-square test. Susceptibilities of ciprofloxacin-resistant isolates and PMQR-positive isolates to other antimicrobial agents are shown in Table 4.
CIPR, ciprofloxacin-resistant isolates; CIPS, ciprofloxacin-susceptible isolates; GENR, gentamicin-resistant isolates; AMKR, amikacin-resistant isolates; CROR, ceftriaxone-resistant isolates; IMPR, imipenem-resistant isolates; PMQR+, PMQR-positive isolates; PMQR-, PMQR-negative isolates.
Sequencing of qnrS, qnrB, aac(6
Clonal relatedness of qnr-positive isolates
For intraspecies genetic diversity analysis, qnr-positive isolates were grouped into three categories: group 1 (G1), qnrS-positive K. pneumoniae isolates; group 2 (G2), qnrS-positive E. coli isolates; and group 3 (G3), qnrB-positive K. pneumoniae isolates. Single qnr-positive isolates belonging to different species were excluded from fingerprinting analysis.
A total of 33 genetic profiles were generated by ERIC-PCR analysis of 34 qnrS-positive K. pneumoniae isolates. Moreover, a total of 24 ERIC types (G1E1–G1E24) were identified based on 85% similarity levels. Isolates with ≥85% similarity were considered to be variants of a given type. On the other hand, RAPD-PCR showed 32 distinct profiles that were classified into 27 RAPD types (G1R1–G1R27). ERIC-PCR analysis of 22 qnrS-positive E. coli isolates showed 22 distinct patterns and 16 ERIC types (G2E1–G2E16). The same number of patterns was obtained by RAPD PCR, but with more RAPD types (n = 17). Regarding the qnrB-positive K. pneumoniae isolates, ERIC-PCR analysis showed 14 distinct profiles classified into 7 ERIC types, while more types were identified by RAPD-PCR (n = 10). For some isolates, their RAPD profile showed 85% similarity or more, they could be differentiated by ERIC-PCR, and were reallocated into different ERIC types. Similarly, isolates of the same ERIC type could also be differentiated by RAPD-PCR. Accordingly, dendrograms based on the combined fingerprints generated by ERIC- and RAPD-PCRs were constructed (Fig. 3A–C).
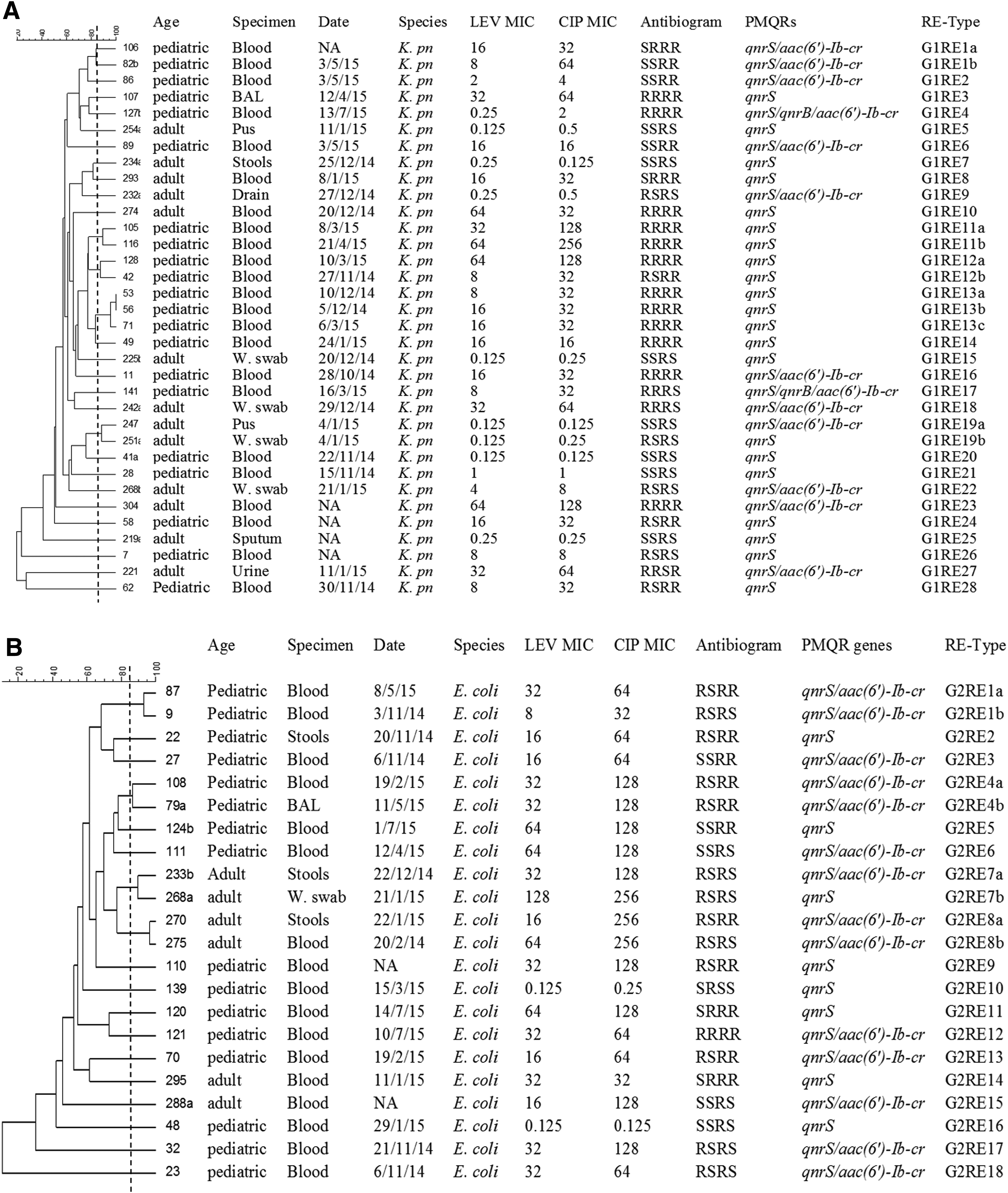
Dendrograms constructed using the combined ERIC-RAPD patterns of qnr-positive isolates.
Discussion
FQs have been widely used as the drugs of choice for antimicrobial prophylaxis of bacterial infections in cancer patients. 18 However, the problem of FQ resistance is becoming increasingly serious with their extensive use. Accordingly, regular regional review of bacterial causes of infections and incidence of antimicrobial resistance is imperative for implementing effective antibiotic policies. 1 Although some surveys have provided data on FQ resistance and the presence of PMQR genes in clinical isolates from Egypt and other Arabic countries, 19 lack of adequate data on bacterial infections of cancer patients was the reason for conducting the current study.
Among 239 Gram-negative isolates from infected patients with cancer included in the current study, a considerably high prevalence of FQ resistance was detected. The highest prevalence of ciprofloxacin resistance was shown by A. baumannii isolates (95.8%), followed by E. coli (83.2%) and K. pneumoniae (62.2%), than other Enterobacteriaceae (31.6%). P. aeruginosa isolates showed the lowest resistance to ciprofloxacin (26.3%). Such a high FQ resistance was accompanied by the existence of PMQR determinants in 53.6% of the clinical isolates. In comparison with other studies on noncancer patients in Egypt,20,21 the prevalence of qnr genes and aac(6′)-Ib-cr in K. pneumoniae isolates in the current study was much lower than that reported by El-Badawy et al. (56.1 vs. 74.% and 36.6 vs. 61.0%, respectively). Another Egyptian study conducted on extended-spectrum beta lactamase (ESBL)-producing E. coli has also reported a slightly higher prevalence of qnr genes compared with E. coli isolates in our study (26.6 vs. 24.2%), while the prevalence of aac(6′)-Ib-cr was much lower (23.3 vs. 53.7%). 21
Of all detected PMQR determinants, aac(6′)-Ib-cr was the dominant one (36.8%). Similar findings were reported by Hassan et al. and other studies in Arabic countries.22,23 In contrast to the mentioned studies that reported high prevalence of the qnrB gene, the qnrS gene was the most frequently detected among qnr genes in this study. Contrary to previous studies,20,24 the qnrA gene was detected in a P. mirabilis isolate (0.4%) from a wound infection. An even higher prevalence of the qnrA gene (16.6%) was reported by Hassan et al. in E. coli isolates from Egypt and in K. pneumoniae isolates from Kuwait (3.7%) than in this study, in which the qnrA gene was not detected in any of the mentioned species. It is worth mentioning that some qnr genes were not screened in the current study and a higher prevalence of such determinants may actually exist in our collection.
Although the qepA gene could be detected before in K. pneumoniae clinical isolates from Egypt 20 and E. coli isolates from Algeria, 25 qepA was not detected in the current study. Consistent with our finding, some worldwide studies also reported the absence of the qepA gene.26,27 Previous studies showed that oqxAB genes are mostly located on the chromosome of Klebsiella sp. and that their presence may not correlate with FQ resistance, probably due to different expression levels. 28 Accordingly, oqxA- and oqxB-positive Klebsiella sp. isolates were not considered among the PMQR-positive isolates in the current study.
While a significant association between qnr genes and aac(6′)-Ib-cr was reported by other studies,24,29,30 such association was not detected in the tested isolates, indicating that such PMQR determinants can circulate independently.
PMQR determinants were identified among almost all tested species, including nonfermenters such as P. aeruginosa and A. baumannii, a fact that may argue for their interspecies transferability. The current finding also confirms previous knowledge about the higher prevalence of PMQR determinants among members of Enterobacteriaceae than nonfermenters (63.3% vs. 9.3%, p < 0.001) possibly due to the higher contribution of other resistance mechanisms such as target site mutations and hyperactive efflux pumps.5,31 Among the Enterobacteriaceae members, PMQR determinants were found more frequently in K. pneumoniae isolates. Similar findings were previously reported in K. pneumoniae isolates recovered from noncancer patients.32,33
Unexpectedly, higher ciprofloxacin resistance and PMQR prevalence were detected among isolates from pediatric patients than those from adults (74.7% vs. 64.0% and 59.3% vs. 43.8%, respectively) and such findings may suggest the misuse or inevitable prescription of FQs in this population. Another possible explanation is that the coselection of PMQR determinants might be promoted by the administration of cephalosporins or aminoglycosides. Similar findings in a previous study were attributed to their increased contact with adults as well as exposure to some environmental factors such as food. 34 Lower prevalence of PMQR determinants (32.1%) was reported in ESBL-producing Enterobacteriaceae isolates recovered from pediatric patients in Mexico 35 as well as in E. coli and K. pneumoniae (9.7%) isolates recovered from a pediatric hospital in Korea. 36
Notably, PMQR-positive isolates revealed higher prevalence of ciprofloxacin nonsusceptibility (76.6% vs. 64.0%) and higher ciprofloxacin MIC values (MIC50 = 64 vs. 32 μg/ml) than PMQR-negative isolates. This is in line with others reports suggesting that the presence of PMQR genes could facilitate the accumulation of further mutations leading to a higher level of FQ resistance. 36
The most alarming finding in our study along with other studies32,37 was the detection of PMQR genes in isolates classified as ciprofloxacin susceptible according to CLSI guidelines. This might justify the lower prevalence of PMQR determinants reported from other studies conducted on ciprofloxacin-resistant isolates only. 33 The clinical significance of this finding arises from the possible selection from low- to high-level resistance and subsequent treatment failure upon use of FQs for treatment of infections caused by such isolates, as previously reported.38,39 The mentioned threat is greatly underestimated by the current CLSI breakpoints and implies the urgent need for reducing ciprofloxacin nonsusceptibility cutoff values to an MIC of 0.125 μg/ml.
Moreover, the results showed that about 91.1% of isolates harboring more than one PMQR gene were nonsusceptible to ciprofloxacin and 67.6% of them showed a high level of resistance. This finding may support the hypothesis that the accumulation of more than one PMQR determinant in one isolate may lead to FQ resistance.5,33,38,40
A long-standing question about quinolone resistance has been its close association with resistance to other agents, particularly β-lactams and aminoglycosides. 41 Upon analysis of such association, ciprofloxacin resistance was found to be significantly associated with resistance to each of gentamicin, amikacin, ceftriaxone, and imipenem (p < 0.001). Meanwhile, PMQR determinants significantly correlated with ceftriaxone (p = 0.002) and gentamicin resistance (p = 0.006). A possible explanation is the fact that PMQR genes are mostly colocated with other resistance determinants (ESBL, AmpC-type β-lactamase, and carbapenemase genes) within transposons and/or integrons in multidrug resistance plasmids.5,6 We might therefore agree with the idea that β-lactams, quinolones, and aminoglycosides can act as potential coselectors for antimicrobial resistance. Accordingly, the use of FQs for prophylaxis may deprive clinicians of the freedom of selecting an effective therapy for bacterial infections in cancer patients.
The diversity of ERIC and RAPD profiles obtained suggests that dissemination of PMQR-positive isolates among K. pneumoniae and E. coli was unlikely to be caused by the spread of specific clones. Instead, PMQR genes were widely distributed among genetically unrelated isolates in different clusters. This is probably due to the horizontal transfer of plasmids carrying such genes, which was reported by some previous studies as well.26,32 Nevertheless, some isolates with an identical pattern were recovered from different patients at different times, suggesting low-level clonal dissemination of these strains. Moreover, a single case of infection control problem was evident in the ICU where two qnrB-positive K. pneumoniae isolates with more than 85% genetic identity were recovered in the same week from two patients.
In a study by Lim et al. in 2009 on ESBL-producing E. coli isolates, ERIC-PCR was more discriminative than RAPD. 42 In contrast, for Serratia marcescens genotyping, Patton et al. found more profiles with RAPD than with ERIC-PCR. 43 With respect to our isolates, although more RAPD types were identified, some isolates whose RAPD profiles showed 85% similarity or more could be differentiated by ERIC-PCR. Accordingly, a combined analysis using fingerprints from the two typing techniques was done. This was also reported by previous studies44,45 to be more discriminating than individual techniques.
Conclusion
High prevalence of FQ resistance was detected among Gram-negative bacteria causing infections in cancer patients in our region. This was accompanied by a high prevalence of PMQR genes that was unexpectedly higher among pediatric patients. PMQR genes were detectable in genetically diverse isolates that were in some cases susceptible to ciprofloxacin. Resistance to FQs was often coexpressed with resistance to other classes of antimicrobials employed in management of bacterial infections in cancer patients.
In summary, findings emerging from several aspects of our study raised concerns about the efficacy of FQ use for prophylaxis of bacterial infections in cancer patients in our region where a high prevalence of FQ resistance was detected. In this concern, it would be recommended to consider the discontinuation of FQ use as prophylactic agents, application of antibiotic rotation strategies, or a combination of ciprofloxacin with oral nonabsorbable antibiotics such as colistin. Infection control practices and antimicrobial stewardship programs are also crucial in combating antimicrobial resistance. Detection of PMQR genes in ciprofloxacin-susceptible bacteria, particularly in isolates displaying a phenotype of decreased susceptibility, sheds light on the urgent need to reconsider the existing FQ breakpoints defined by the CLSI.
Footnotes
Acknowledgments
The authors are grateful to the Microbiology and Immunology department, Faculty of pharmacy, Ain Shams University, as well as the Microbiology and Immunology department, Faculty of pharmacy, October University for Modern Sciences and Arts, for providing the laboratory facilities for this work. The authors would also like to acknowledge the Microbiology Laboratory of the National Cancer Institute, Cairo University, for providing the clinical specimens.
Disclosure Statement
No competing financial interests exist.