
Abstract
Salmonella spp. are health-threatening foodborne pathogens. The increasingly common spread of antibiotic-resistant Salmonella spp. is a major public healthcare issue worldwide. In this study, we wished to explore (1) antibiotic or polypeptide combinations to inhibit multidrug-resistant Salmonella bredeney and (2) the regulation of cross-resistance and collateral sensitivity of antibiotics and polypeptides. We undertook a study to select antibiotic combinations. Then, we promoted drug-resistant strains of S. bredeney after 15 types of antibiotic treatment. From each evolving population, the S. bredeney strain was exposed to a particular single drug. Then, we analyzed how the evolved S. bredeney strains acquired resistance or susceptibility to other drugs. A total of 105 combinations were tested against S. bredeney following the protocols of CLSI-2016 and EUCAST-2017. The synergistic interactions between drug pairings were diverse. Notably, polypeptides were more likely to be linked to synergistic combinations: 56% (19/34) of the synergistic pairings were relevant to polypeptides. Simultaneously, macrolides demonstrated antagonism toward polypeptides. The latter were more frequently related to collateral sensitivity than the other drugs because the other 13 drugs sensitized S. bredeney to polypeptides. In an experimental evolution involving 15 drugs, single drug-evolved strains were examined against the other 14 drugs, and the results were compared with the minimal inhibitory concentration of the ancestral strain. Single drug-evolved S. bredeney strains could alter the sensitivity to other drugs, and S. bredeney evolution against antibiotics could sensitize it to polypeptides.
Introduction
A
Food pathogens, especially Escherichia coli O157 and Salmonella enterica serotype Typhimurium, are encroaching on humans with increasing morbidity and mortality. 7 MDR food pathogens and resistant genes have been spreading in China and worldwide,8,9 leaving conventional clinical treatments against bacterial infection compromised. 10 MDR pathogens of various types, such as moulds, 11 parasites, 12 and viruses, 13 will be difficult to treat. In particular, the threat posed by gram-negative bacteria has been reported,14,15 suggesting that more effective clinical strategies are needed urgently in this area.5,16
Researchers have investigated antimicrobial interactions in Staphylococcus aureus and E. coli and demonstrated the sensitivity and cross-resistance of certain drugs.17–20 However, such interactions between antibiotics are dependent upon the drugs and bacterial species involved. 21
Salmonella spp. are known to cause food poisoning, 22 and MDR strains can be lethal. Li et al. reported that MDR Salmonella spp. isolated from food products are major threats, with 59.4% of Salmonella typhimurium strains resistant to more than five antibiotics. 23
Antibiotics are widely used in the livestock industry, where Salmonella infection can be common. 24 Farmers tend to add antibiotics into feedstuff to promote animal growth for financial reasons. Animals are the main consumers of antibiotics, which are provided for growth improvement 16 and disease control. 25 A survey conducted in China showed that nearly half of annual antibiotic usage was in the livestock industry. 26 Antibiotic-resistant organisms or genes transferring to humans via foodstuffs occur in a specific way. That is, if animals are given feedstuff containing antibiotics, some antibiotics will be absorbed after oral administration and the other unabsorbed antibiotics will be excreted in the feces. Some of the absorbed antibiotics will be metabolized, and the breakdown products may be excreted in feces and urine. 27 The manure of animals will be reapplied as fertilizers, and the manure from antibiotic-overdosed animals can become threats to water and soils.28,29 Humans are the main consumers of food-producing animals and edible plants, so there is a high risk that antibiotics will enter the human body and accumulate. This indirect ingestion of antibiotics can be highly dangerous to humans. 30 If farmers follow withholding times for antibiotics, no antibiotics or residues should be present in meat or other animal products. Owing to the overuse of antibiotics against bacterial infections in humans and animals, Salmonella spp. have evolved considerable resistance, with MDR strains emerging. 31 In recent years, MDR Salmonella infection has become a major issue. 32 Some researchers have suggested that the progress made in new strategies against Salmonella infection lags far behind the pace of bacterial evolution. 33 Several studies focusing on the drug-resistant mechanism of Salmonella have been undertaken, and theories on the resistance in a single drug have been elucidated. 34 However, the interaction between drugs is considered to be complicated and incompletely understood.
Multidrug combination therapies against bacterial infection have been widely studied and reported. With regard to methicillin-resistant S. aureus (MRSA) infection, 17 Gonzales et al. combined three β-lactam drugs, meropenem, piperacillin, and tazobactam, targeting cell wall synthesis to inhibit bacterial growth. Bakthavatchalam et al. tested the efficacy of a combination of cefixime and ofloxacin against S. typhimurium and observed a synergistic effect. 35 Chang et al. used a combination of cefotaxime and ciprofloxacin to inhibit nontyphoid salmonellae and achieved favorable results. 36
Salmonella bredeney has been reported to be a seriously infectious serotype.37–39 We undertook a study to select antibiotic combinations. Then, we promoted drug-resistant strains of S. bredeney after the treatment of 15 antibiotics. From each evolving population, the S. bredeney strain was exposed to a certain single drug. Then, we analyzed how the evolved S. bredeney strains acquired resistance or susceptibility to other drugs.
Materials and Methods
Materials
S. bredeney CDC-1 was an isolate derived from cold dishes. It was isolated, characterized and donated by Jiangsu Provincial Centre for Disease Control and Prevention (Jiangsu, China). All antibiotics and polypeptides that we applied (Table 1) were prepared from the powder stocks and solvents recommended by the Clinical and Laboratory Standards Institute in 2016 (CLSI-2016). 40 Antibiotic solutions were sterilized using a filter (pore size, 0.22 μm) and stored at −20°C before use.
Antibiotics and Polypeptides Used
CLSI, Clinical and Laboratory Standards Institute; MIC, minimal inhibitory concentration.
Methods
Minimal inhibitory concentration test
All experiments were undertaken in Mueller–Hinton broth according to CLSI-2016. A MIC assay was conducted with the broth dilution method following CLSI recommendations. In brief, S. bredeney CDC-1 was inoculated in Luria broth and incubated overnight in 37°C with shaking at 180 rpm. Then, the cultures were diluted to 0.5 McFarland (≈2 × 108 colony-forming units (CFU)/mL). After 100-fold dilution, the strain was placed in 2-fold-diluted antibiotic gradient plates and incubated at 37°C without shaking for 18–20 hr. The endpoint was determined by optical density at 600 nm (OD600) >0.1. The experiment was conducted in triplicate. The drugs used are shown in Table 1.
Synergistic interactions of antibiotic combinations
An antibiotic combination assay was conducted using the broth dilution method following the protocol described in the work by Gonzales et al. 17 with some modifications. Drug pairings were tested by combination with each of the 15 single drugs to make double combinations. The highest concentration of each drug was determined by the MIC against the S. bredeney CDC-1 strain that we tested before. It was four- or eightfold of the MIC. Thus, the 15 drugs were formulated into 105 pairwise combinations. Each drug combination was prepared with Mueller–Hinton broth in a twofold dilution series down 10 rows in 96-well plates with 100 μL in each well. The overnight incubated S. bredeney CDC-1 strain was diluted to 0.5 McFarland and mixed 1:100 with fresh Mueller–Hinton broth. Then, it was added to the 96-well plates containing drug-combination gradients with a volume of 100 μL in each well. The 96-well plates were incubated at 37°C without shaking for 18–20 hr. The endpoint was also determined by OD600 > 0.1. Each of the drug pairings was tested in triplicate.
The synergistic activity of combined drugs was determined by the fractional inhibitory concentration index (FICI)
41
:
where MIC[A]AB signifies the concentration of drug A in the combined drug AB, and MIC[A] signifies the MIC of the tested strain against the single drug A. FICI ≤0.5 denoted synergy; 0.5 < FICI <4.0 denoted no interaction; and FICI ≥4.0 denoted antagonism.
S. bredeney evolution
Evolution experiments were conducted in a twofold dilution gradient in 96-well plates with a final volume of 100 μL of liquid culture per well. In brief, the original overnight bacterial cultures were diluted at 0.5 McFarland standard (≈2 × 108 CFU/mL) and then diluted 100-fold before addition onto gradient antibiotic plates. After incubation for 20 hr without shaking at 37°C, the culture in the columns at the highest concentration of antimicrobial agent with an OD >0.2 was diluted 200-fold and propagated into a fresh plate with gradients of antibiotic concentration. The concentration was recorded each time before propagation over 37 days.
The highest concentration of the antibiotic in the first column was determined by the MIC of the evolved strain. It was maintained at least fourfold of the MIC to ensure selective pressure. The evolution experiment for 15 drugs was done in duplicate.
Cross-resistance and collateral sensitivity
The evolved S. bredeney strain in each of the 15 drugs selected (Table 1) was inoculated onto 96-well plates with the other 14 drugs in a twofold dilution gradient. It was incubated at 37°C for 18–20 hr without shaking according to the protocol of the MIC test mentioned above. In parallel, the ancestral strain was plated at the same dilution into the same antibiotic gradients as a negative control. This experiment was carried out in triplicate.
Results
Antibiotic resistance of S. bredeney CDC-1
According to the MIC results of S. bredeney CDC-1 against 15 drugs (Table 1), the S. bredeney strain showed considerable resistance to ampicillin, streptomycin, and tetracycline according to CLSI-2016. It also showed resistance to colistin according to guidelines set by the European Committee on Antimicrobial Susceptibility Testing in 2017. 42 S. bredeney CDC-1 revealed resistance to at least one antibiotic from each class tested.
Synergistic and antagonistic interactions of drug pairings
The screening of the 15 drug combinations revealed synergistic and antagonistic interactions (Table 2). One-hundred and five interactions were detected with the 15 drugs, with 5 showing antagonism, 34 showing synergy, and 66 neither antagonism nor synergy, with FICI values ranging from 6 to 0.094. Antagonistic interactions occurred frequently and exclusively between polypeptides and macrolides. Otherwise, polypeptides were more likely to be linked to synergistic combinations: 19 out of 34 of the synergistic pairings involved polypeptides.
The Antibiotic Combinations Involved in the Synergistic Test and the Fractional Inhibitory Concentration Index of the Different Drug Combinations
FICI ≤0.5 indicates synergy; 0.5 < FICI <4.0 indicates no interaction; FICI ≥4.0 indicates antagonism.
FICI, fractional inhibitory concentration index.
Moreover, antibiotics in the same class could demonstrate synergistic (streptomycin–amikacin, FICI = 0.375), antagonistic (colistin–polymyxin B, FICI = 6), or no interaction (aureomycin–tetracycline, FICI = 1).
S. bredeney evolution
S. bredeney CDC-1 evolved in the presence of 15 drugs in six classes (polypeptides, macrolides, aminoglycosides, tetracyclines, β-lactams, and quinolones). Evolution occurred in all populations in 37 days. The extent of these changes was shown in Fig. 1.
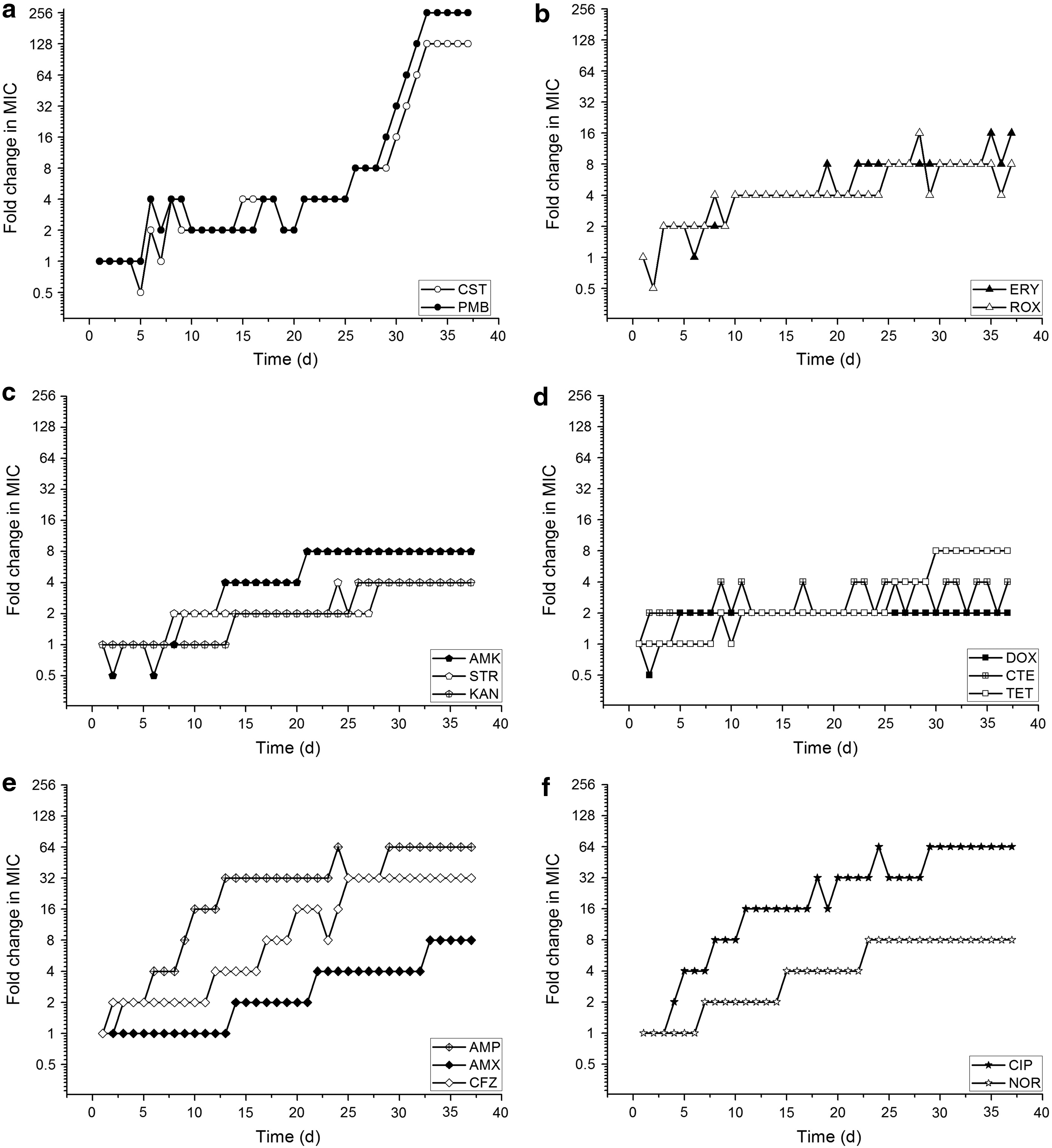
Fold change in MIC against antibiotics in Salmonella bredeney CDC-1 during the evolution experiment. The lines and symbols in the same shade represent antibiotics in the same class. The various shades indicate the antibiotics of polypeptides, macrolides, aminoglycosides, tetracyclines, β-lactams, and quinolones, respectively
S. bredeney was originally resistant to all the tested drugs and the MICs against β-lactams (ampicillin, amoxicillin, and cefazolin) increased the fastest. The MICs of the strains increased slowly against polypeptides (colistin and polymyxin B) in the first 25 days, but accelerated after that time. The increase in MIC against doxycycline was slow, with only twofold change in MIC from day 5 to 37. The MIC increase against tetracyclines was the most unstable among all the classes tested. MICs against aureomycin varied from two- to fourfold from day 9. Finally, the MICs were maintained to at least fourfold of their original values after S. bredeney evolution.
Cross-resistance and collateral sensitivity
The interactions between antibiotics and polypeptides in S. bredeney CDC-1 were displayed in Fig. 2. Twelve types of drug-evolved strains became cross-resistant to ampicillin and no sensitive strains evolved. Ten types of drug-evolved strains gained cross-resistance to tetracycline and one sensitive strain evolved. Thirteen drug-evolved strains became sensitive to colistin and polymyxin B, respectively. Notably, strains exposed in the other drug lines acquired higher MIC against a certain drug, but showed lower MIC against colistin and polymyxin B. We suggest that S. bredeney evolution in other drugs could be inhibited readily by colistin and polymyxin B.

Interaction networks of different antibiotics in S. bredeney CDC-1. Circles with the same pattern indicate antibiotics from the same class (e.g., circles with diagonal lines represent macrolides, which includes erythromycin and roxithromycin). Solid arrows indicate cross-resistance
Antibiotics from the same class showed cross-resistance in most cases. Such cross-resistance was found between four drug pairings: aureomycin–doxycycline (tetracyclines), amikacin–kanamycin (aminoglycosides), amikacin–streptomycin (aminoglycosides), and ampicillin–amoxicillin (β-lactams). Moreover, “mutual cross-resistance” (i.e., previous exposure of S. bredeney CDC-1 to one drug conferred a higher MIC to another drug and vice versa) was displayed more frequently between antibiotics of the same class. This effect was shown in five drug pairings: colistin–polymyxin B (polypeptides), erythromycin–roxithromycin (marcrolides), doxycycline–aureomycin (tetracyclines), ciprofloxacin–norfloxacin (quinolone), and ampicillin–cefazolin (β-lactams).
Drugs from different classes could also confer cross-resistance and even mutual cross-resistance: kanamycin–norfloxacin, erythromycin–ampicillin, and aureomycin–cefazolin. β-lactam antibiotics showed cross-resistance more frequently than those in the other classes.
“Reciprocal collateral sensitivity” refers to bacterial strains that have evolved against a single drug and become sensitive to another drug and vice versa. We found eight drug pairings that fitted this description: roxithromycin–colistin, doxycycline–colistin, roxithromycin–polymyxin B, streptomycin–ciprofloxacin, ciprofloxacin–polymyxin B, colistin–norfloxacin, roxithromycin–amikacin, and colistin–aureomycin.
All the drugs tested except polypeptides could sensitize S. bredeney CDC-1 to colistin and polymyxin B. Hence, use of antibiotics was more likely to result in collateral sensitivity against polypeptides than against the other drugs.
Of the 15 drugs tested, each drug showed no interaction with at least 1 other drug except for polypeptides. However, a single drug did not show complete indifference to all the other drugs indicating that, once an antibiotic was used, it could interact with other drugs and result in resistance or sensitivity.
Discussion
In this work, the MDR strain S. bredeney CDC-1 was used to screen for combinational drug pairings. Among the 105 drug pairings screened, five combinations (4.8%) showed antagonistic interactions, 34 pairings (32.4%) showed synergistic interactions, and 66 pairings (62.8%) demonstrated no interaction. The most favorable combination was streptomycin–polymyxin B (FICI = 0.094) (Table 2).
Compared with similar laboratory-based evolution experiments in E. coli, some differences were seen. Kanamycin and ampicillin displayed reciprocal collateral sensitivity in E. coli according to Lázár et al., 18 but demonstrated mutual cross-resistance in S. bredeney in our work. In the work by Gonzales et al., the β-lactams (meropenem, piperacillin, and tazobactam) showed collateral sensitivity against MRSA. 17 However, according to our findings, antibiotics from the same class showed mutual cross-resistance more often and with no collateral sensitivity. Thus, we consider that the evolutionary response of bacteria toward antibiotics is probably conserved in certain species.
In our study, β-lactam-evolved strains did not show collateral sensitivity to macrolides and vice versa. The use of β-lactams (ampicillin, amoxicillin, and cefazolin) evolved S. bredeney strains that became more resistant to erythromycin, but showed little interaction with roxithromycin. Simultaneously, the erythromycin-evolved strain became more resistant to the three β-lactam antibiotics, and the roxithromycin-evolved strain increased its MIC only against cefazolin and ampicillin, suggesting that β-lactam- and macrolide-evolved resistance could cause cross-resistance to each other. Our results also demonstrated that, although the general mechanism of resistance of the two classes differed, the resistance could also be mediated by other mechanisms. Also, roxithromycin, an erythromycin-modified antibiotic, showed fewer interactions with β-lactams than with erythromycin.
Tetracyclines bind to ribosomes and inhibit the elongation phase of protein synthesis. Ribosomal protection proteins promote resistance to tetracyclines, but may not be the only mechanism of resistance. 43 Halling-Sørensen et al. investigated enzymes which can degrade tetracyclines in certain bacterial species. 44 Also, it has been suggested that in gram-positive and gram-negative bacteria, efflux pumps have important roles in tetracycline resistance. 45 The mechanism of bacterial resistance to marcrolides is similar to that of tetracyclines in several ways.46,47 Hence, the cross-resistance observed between tetracyclines and macrolides in our study may be associated with the mechanism of resistance mentioned above.
Quinolones are broad-spectrum antibacterial agents. They block the activity of DNA gyrase and topoisomerase IV to inhibit bacterial viability. Acquisition of bacterial resistance is based mainly in the chromosomal mutations that result in the loss of a target for antibiotics to bind to. 48 Also, the mechanism of resistance of quinolones could be mediated by efflux pumps. 49 In our study, adaptation to norfloxacin showed an increase in MIC against eight antibiotics and a decrease in MIC against only two polypeptides. The adaptation to ciprofloxacin, however, sensitized S. bredeney to seven antibiotics, and S. bredeney became more resistant to only two β-lactams. Mediation by efflux pumps could explain the increasing resistance, but the bacterial interaction with drugs was mediated by different mechanisms. Hence, the decrease in MIC of quinolone-evolved strains could involve other mechanisms that merit further study.
Polymyxin B and colistin were found to be mutually cross-resistant because they belong to the same drug class and have a similar mechanism of resistance. The latter has been reported to be associated with changes in regulation of lipopolysaccharide (LPS) production in the bacterial wall and alterations in the targets for these two drugs to bind to. 50 In our study, polymyxin B- and colistin-adapted S. bredeney strains evolved resistance to the β-lactams ampicillin, amoxicillin, and cefazolin, the resistance mechanism of which has been reported to be the degradation and modification of β-lactamases.
Moreover, adaptation to polymyxin B and colistin exerted different effects on the other drugs tested. Polymyxin B and colistin were sensitized to ciprofloxacin and roxithromycin; adaptation to polymyxin B resulted in sensitization to kanamycin and norfloxacin; and adaptation to colistin resulted in sensitization to doxycycline and aureomycin.
Interestingly, the other 13 drugs all sensitized S. bredeney to polymyxin B and colistin. It is thought that polymyxin B and colistin inhibit bacterial growth by electrostatic interactions with LPS on bacterial membranes. 51 However, the mechanism of action of this collateral sensitivity requires further investigation.
For the aminoglycosides used in the present study (kanamycin, amikacin, and streptomycin), the evolved S. bredeney strain may have modified (possibly by methylation) the target ribosome that binds with the aminoglycoside.52,53 In this way, the altered ribosome could impede its function, and the gene expression associated with cell membranes could be affected, making cell membranes more vulnerable. Hence, the electrostatic interaction of colistin and polymyxin B targeting LPS on the surface would be more effective.
The present study suggested that the S. bredeney strains tested adapted to the other 13 drugs after they became sensitized to the polypeptides colistin and polymyxin B (Fig. 2b). Simultaneously, synergistic combinations were associated most frequently with polypeptides (Table 2). Nineteen out of 34 synergistic drug pairings were related to colistin and polymyxin B. Also, of the 27 combinations that involved colistin and polymyxin B, 10 were related to colistin and 9 were related to polymyxin B.
There were 105 drug pairings and 210 interactive relationships (Fig. 3). We found that that 32.4% (34), 4.8% (5), and 62.8% (66) drug pairings displayed synergy, antagonism, and no interaction, respectively. Each of the drug pairings covered relationships: cross-resistance, collateral sensitivity, or no interaction. Thus, there were 34 synergistic interactions, 5 antagonistic interactions, and 66 noninteractions. Also, interactive relationships tended to be distributed randomly in the three drug-combination groups. Even though the other 13 drug-evolved S. bredeney strains were sensitized to the polypeptides colistin and polymyxin B, a combination of colistin and polymyxin B displayed different efficacies. Polymyxin B–erythromycin (FICI = 5) and colistin–erythromycin (FICI = 6) were the two worst interactions and polymyxin B–streptomycin (FICI = 0.094) and colistin–streptomycin (FICI = 0.125) were the best. These observations suggested that collateral sensitivity could not ensure a synergistic effect of the antibiotic combinations. Hence, these synergistic and antagonistic interactions were not directly relevant to the collateral sensitivity and cross-resistance of the antibiotics.
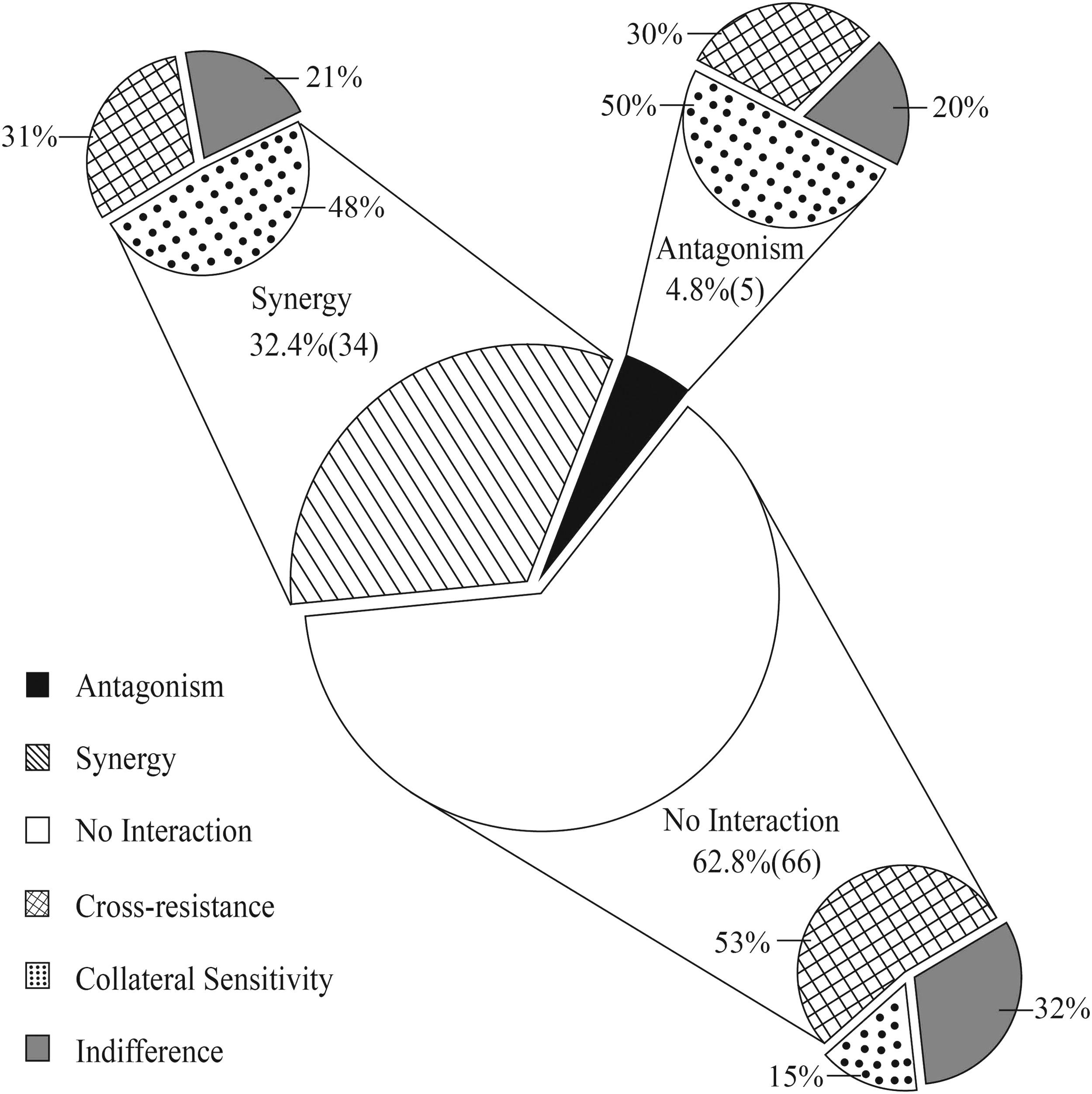
Antibiotic combination and mutual interaction. The pie chart in the center demonstrates the results of antibiotic combination. The black part denotes antagonism, diagonal lined pattern indicates synergy, and white denotes no interaction. The other three pie charts represent mutual interactions of the drugs in the drug pairings. The checkered pattern denotes cross-resistance, the black dots indicate collateral sensitivity, and gray part denotes no indifference. For example, among the 68 synergistic drug pairings, which formed 132 mutual interactions, 48% showed collateral sensitivity, 31% showed cross-resistance, and 21% showed no interaction.
Evolution of a bacterial strain within a laboratory is different from that in the environment. In the former, the bacterial population evolves in different drug selection conditions done in parallel. Hence, the different drug conditions are separate and not likely to affect each other. However, in the environment, the situation could be more complicated. Bacteria could be exposed to various antimicrobial agents simultaneously and evolve in different ways.
In the present study, we initiated a series of experiments targeting MDR S. bredeney, including drug-combination selection, experimental evolution, and cross-interaction tests. We obtained a cross-interaction network and a table showing synergy among two polypeptides (polymyxin B and colistin) and 13 antibiotics against S. bredeney. Bacterial evolution against antimicrobial agents is a serious problem, especially for Salmonella spp., because they are important infectious foodborne pathogens. Some of the monotherapy against salmonellae has failed in clinical settings, including treatment using third- or fourth-generation cephalosporins which were effective previously.54,55 Hence, we used MDR S. bredeney in our research. Drug-combination methods against salmonellae have been studied by many researchers. As mentioned above, Chang et al. used a combination of cefotaxime and ciprofloxacin to inhibit nontyphoid salmonellae, and the two drugs showed a synergistic effect. 36 Bakthavatchalam et al. used a combination of cefixime and ofloxacin against S. typhimurium. 35 However, clinical drug-combination treatment against salmonellae has barely been reported. This knowledge gap may be because research on the activity, efficacy, and side effects of drug combinations is lacking. Experimental studies and clinical trials are needed to fill this knowledge gap.
In this way, our results will be complementary to previous work on drug combinations and interactions against Salmonella spp. It may also be helpful for controlling the dispersal of MDR bacteria in the future.
Footnotes
Acknowledgments
The authors thank the native English-speaking scientists of Elixigen Company (Huntington Beach, California) for editing our article. This work was supported by the Social Development Program of Jiangsu Province (grant number BE2012746), the Independent Innovation Program of Jiangsu Province [grant number CX (12)3087], and the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
Disclosure Statement
No competing financial interests exist.