
Abstract
Flies play an important role as vectors in the transmission of antimicrobial-resistant bacteria (ARB) and are hypothesized to transfer ARB between internal and external livestock housing areas. The aim of this study was to understand the role that flies may play in the maintenance of ARB in the farm environment. We first evaluated the fate of ingested antimicrobial-resistant Escherichia coli harboring a plasmid containing antimicrobial-resistance genes (ARGs) throughout the housefly (Musca domestica) life cycle, from adult to the subsequent F1 generation. Antimicrobial-resistant E. coli was isolated from different life cycle stages and ARG carriage quantified. The ingested E. coli persisted throughout the fly life cycle, and ARG carriage was maintained at a constant level in the housefly microbiota. To clarify the transmission of ARB from flies to livestock, 30-day-old chickens were inoculated with maggots containing antimicrobial-resistant E. coli. Based on the quantification of bacteria isolated from cecal samples, antimicrobial-resistant E. coli persisted in these chickens for at least 16 days. These results suggest that flies act as a reservoir of ARB throughout their life cycle and may therefore be involved in the maintenance and circulation of ARB in the farm environment.
Introduction
A
Synanthropic insects, including flies, inhabit the areas surrounding farms where they act as vectors for the dissemination of many microorganisms, including ARB. 4 We have previously proposed that flies not only act as a mechanical vector of ARB in the farm environment, but also as a biological vector, involved in the horizontal gene transfer of antimicrobial resistance genes (ARGs) harboring plasmids in their intestines.5–7
Recently, Pava-Ripoll et al. 8 have demonstrated that pathogenic bacteria ingested by adult flies are transmitted vertically to the next generation. However, the dynamics of ARB maintenance and the carriage of the ARG-carrying plasmids over the fly life cycle remain unclear. In the absence of antimicrobial selective pressure, the growth rates of ARB are typically inferior relative to wild-type strains 9 and they should therefore be eliminated by natural selection. However, other selective pressures may exist in the fly, such as host immunity through antimicrobial peptides that would contribute to the maintenance of ARB. 10 If it is found that ARB and ARGs can persist throughout the life cycle of a fly, the fly would play an important role as a reservoir that maintains ARB in the farm environment.
In livestock-rearing environments, marked attention is paid to the carriage of microorganisms between internally and externally housed livestock to control the spread of ARB and livestock-associated infectious diseases. However, this strategy has not been entirely successful because the transmission routes for many microorganisms have not yet been clarified.11,12 A previous study suggested that the application of fly screens to broiler farms reduced the introduction of Campylobacter spp. into the internal housing area within chicken enclosures. 13 Based on this observation, we propose that a fly-mediated transmission route for the internally housed livestock might consist of maggots hatched from eggs laid by flies that originated in the externally housed livestock. As maggots often contain microorganisms carried by their parental flies, 8 livestock infection by these microorganisms in close contact with maggots is a distinct possibility.
To further investigate this possibility, the aim of this study was to clarify the role of flies in maintaining ARB in the farm environment. We evaluated the fate of ingested antimicrobial-resistant Escherichia coli and ARGs-carrying plasmids throughout the fly life cycle. In addition, to elucidate the possible transmission routes of ARB from flies to livestock, we orally administered maggots containing antimicrobial-resistant E. coli to chickens and quantified these bacteria over time in cecal samples.
Materials and Methods
Houseflies
Chemical Specialties Manufacturers Association (CSMA) housefly strain (Musca domestica) was provided by Sumika Life Tech, Ltd. (Osaka, Japan), and maintained for multiple generations in nets at 25°C under a 14-h light–10-h dark cycle in an isolator. 7 The houseflies were provided distilled water and a 1:1 mixture of skim milk and sugar. E. coli TC7-1 strain used in this study, which was derived from cattle feces as previously described,7,14 and ARGs harbored by the ingested bacteria (tetA and blaCTX-M-2), were not naturally carried by this housefly strain.
Housefly bacterial ingestion and rearing
An overview of the experimental design is shown in Table 1. Approximately 100 adult houseflies (1 day after emergence) were transferred to a cage (15 × 15 × 90 cm) on day 1. The houseflies were fed with suspensions of E. coli TC7-1 strain (resistant to cefazolin, cefotaxime, spectinomycin, and tetracycline) that carried a plasmid containing two ARGs (tetA and blaCTX-M-2) twice a day, on days 1–5, ad libitum. 7 To prepare the bacterial suspensions, cultures for ingestion were grown in 25 mL of Tryptic Soy Broth (Bacto Peptone; BD Biosciences, Franklin Lakes, NJ) at 37°C for 14–18 hours and then centrifuged for 15 minutes at 1,800 g. The supernatants were discarded and the pellets resuspended in 5 mL of a milk–sugar solution that had been previously sterilized at 115°C for 20 minutes. Suspensions were then incubated for 1 hour at 37°C. Houseflies were provided with suspensions on a sterile plate for ingestion.
Experimental Design of the Housefly-Rearing Experiment
After the bacteria were ingested, houseflies were transferred to a new cage. Skim milk, sugar, and sterilized distilled water were provided for 24 hours ad libitum on day 6. On day 7, an oviposition substrate was placed in the cage to collect housefly eggs. The substrate contained a 1:7:14 (w/w/w) mixture of autoclaved rodent diet CE-2 (CLEA Japan, Inc., Tokyo, Japan), autoclaved bran (Oriental Yeast, Tokyo, Japan), and sterilized distilled water. Once the housefly eggs were laid, the oviposition substrate was recovered from the cages. The housefly eggs were surface sterilized with sodium hypochlorite (HONCHO CHEMICAL, INC., Tokyo, Japan) and ethanol (Wako Pure Chemical Industries, Ltd., Osaka, Japan), as previously described, 7 and then transferred to a new oviposition substrate. The substrate, containing hatched maggots, was placed in an isolator at 25°C with a 14-h light–10-h dark cycle until pupation (ca. 6 to 7 days).
Next, the pupae were surface sterilized as previously described and transferred to a Petri dish (90 mm) to allow the F1 adult houseflies to emerge, avoiding crosscontamination with the substrate. The pupae in the Petri dish were placed in a new cage under the same conditions until the emergence of F1 adult houseflies (ca. 4 to 5 days).
Houseflies were collected over several days for bacterial isolation and DNA extraction. Specifically, adult houseflies were collected on day 0, before ingestion of bacteria and day 7, after oviposition; eggs were collected on day 7, maggots were collected on days 10 and 12, pupae were collected on days 14 and 16; and F1 adult houseflies were collected on days 18 or 19. Maggots from day 10 were surface sterilized and stored at −80°C until needed for the experimental chicken model. At −80°C, the maggots died; however, the ingested E. coli strain TC7-1 survived in the maggots as shown by the similar viable counts (colony-forming units—CFUs) before and after storage. Housefly rearing with bacterial ingestion was repeated three times.
As a control for checking the involvement of bacterial ingestion in the development of a control group of houseflies under the same conditions, the development of houseflies reared without bacterial ingestion was conducted as for those with bacterial ingestion. A difference in the development of houseflies was not observed.
Bacterial isolation and DNA extraction
Surface-sterilized individual adult houseflies, maggots, pupae, F1 adult houseflies, and 10 pooled eggs were each homogenized in 0.5 mL of phosphate-buffered saline (PBS) (Wako Pure Chemical Industries, Ltd.). The persistence of the ingested bacteria was evaluated in three homogenized samples from each sampling day by plating the serial dilution of samples onto Deoxycholate Hydrogen Sulfide Lactose (DHL) agar (Nissui Pharmaceutical, Tokyo, Japan) supplemented with 4 μg/mL of cefotaxime (Sigma-Aldrich, St. Louis, MO). 7 After incubation for 24 hours at 37°C, the colonies were counted to determine viable counts (CFU/sample). In addition, the last distilled water rinses after surface sterilization were plated on Tryptic Soy Agar (Bacto Peptone) to confirm the complete removal of bacteria from each sample surface.
DNA was extracted from surface-sterilized samples using the ISOFECAL Kit (Nippon Gene, Tokyo, Japan), according to the manufacturer's instructions. Three extracted DNA samples from each sampling day were used in a quantitative PCR (qPCR).
qPCR was performed to determine the copy number of tetA and 16S rRNA genes. To generate standard curves, the tetA and 16S rRNA genes of E. coli strain TC7-1 were amplified using tetA gene-specific primers (forward: 5′-GCGCCTTTCCTTTGGGTTCTC-3′ and reverse: 5′-CGTGATCGGGAGTATCTGGCTG-3′) that were designed according to the GenBank tetA sequence (accession no. KT950741), in addition to 16S rRNA gene universal primers. 15 The PCR products were cloned into a pTA2 vector and inserted into E. coli strain DH5α using the TArget Clone™ Kit (TOYOBO, Osaka, Japan), according to the manufacturer's instructions. Transformants harboring the recombinant plasmid were selected and the plasmids extracted using the QIAprep Spin Miniprep Kit (Qiagen, Hilden, Germany). The concentrations of purified recombinant plasmid DNA in the samples were determined spectrophotometrically (BioSpectrometer; Eppendorf, Hamburg, Germany).
qPCR was performed using SYBR® Premix Ex Taq™ II (TaKaRa, Shiga, Japan) in 20 μL reactions containing 5 μL of template DNA and 0.4 μM of each primer.16,17 The thermal cycling conditions included a holding stage for 30 seconds at 95°C, followed by 40 cycles of 5 seconds at 95°C (denaturation), and 30 seconds at 50°C (for tetA) or 60°C (for 16S rRNA) (annealing and extension) in a LightCycler 480 system (Roche, Mannheim, Germany), according to the manufacturer's instruction.
Experimental chicken model
Animal experimentation protocol was approved by President of Kitasato University through the judgment of the Institutional Animal Care and Use Committee of Kitasato University (approval no. 15-135). 18 The chickens were reared in individual wire cages in an isolation building with a nonmedicated layer ration and water supplied ad libitum.
Chickens (30 days old; “Day 30”) were assigned to three groups (n = 5 per group). On days 30 and 32, the chickens were orally inoculated in the following groups; group 1 was a model for the fly-associated ARB transmission route and consisted of 10 pooled maggots containing E. coli strain TC7-1 (ca. 103 CFU/maggot), prepared as previously described; group 2 was a positive control consisting of a suspension of E. coli strain TC7-1 in saline (109 CFU/chicken); and group 3 was a negative control inoculated with saline.
Cecal droppings or contents were collected on days 31, 32, 33, 36, 38, 40, 43, and 46. The samples were homogenized in PBS. ARB were quantified by performing serial dilution in PBS and plating onto DHL supplemented with 32 μg/mL of cefazolin (Sigma-Aldrich) and 32 μg/mL of spectinomycin (Sigma-Aldrich) for E. coli strain TC7-1. The colonies were counted after incubation for 24 hours at 37°C to determine the viable cell count [CFU/cecal sample (g)].
Data analysis
Data are presented as box-and-whisker plots, where the maximum and minimum values are displayed as whiskers. The interquartile ranges indicate quartile distribution [the distance between the third (75%) and first (25%) quartiles] and are displayed as boxes.
Statistical significance was determined using a Mann–Whitney U test, with the significance threshold set at p < 0.05.
Results
Persistence of the ingested ARB in houseflies throughout their life cycle
The ingested antimicrobial-resistant E. coli strain persisted in the houseflies throughout their life cycle, from adult houseflies, as they proceeded through oviposition and metamorphosis, to the next generation of adult houseflies. Adult houseflies (day 7) harbored 102–105 CFU of the ingested antimicrobial-resistant E. coli per housefly (Fig. 1A). After oviposition, five of the nine (55.6%) pooled egg samples (day 7) contained the ingested strain, with a mean CFU per sample of 2.1 × 101. On days 10 and 12, this value increased to 1.7 × 103 and 8.9 × 103 CFU/maggot, respectively. After the maggots had pupated, the counts decreased to an average of 101 CFU/pupae (days 14 and 16). Five of the nine (55.6%) newly emerged F1 adult houseflies (days 18 to 19) harbored 1.3 × 101 CFU/housefly.

Persistence of antimicrobial-resistant Escherichia coli and antimicrobial resistance genes in houseflies throughout their life cycle.
No bacteria were detected in any of the final distilled water rinse from the last step of the surface sterilization procedure.
Persistence of the ingested plasmid-encoded tetA gene in houseflies throughout their life cycle
The results of qPCR revealed that there were between 103 and 1010 copies of tetA gene per adult housefly (Fig. 1B). The tetA gene was detected in five of the nine (55.6%) pooled egg samples, with a mean number of 1.1 × 103 copies/sample. Later in the fly life cycle, the tetA gene copy number increased from 5.4 × 104 to 7.7 × 104 copies/maggot. Copy number was then maintained throughout pupation, although at somewhat lower levels of 1.4 × 103 to 8.2 × 103 copies/pupae on days 14 and 16, respectively. Finally, the tetA gene was identified in five out of the nine (55.6%) newly emerged F1 adult houseflies, with an average of 2.1 × 103 copies/housefly.
To assess the potential persistence of the tetA gene in houseflies throughout their life cycle, the ratios of tetA to 16S rRNA gene copies were determined (Fig. 1C). The mean copy number of the 16S rRNA gene ranged from 105 to 109 copies/sample [adult houseflies, 3.0 × 108; eggs, 1.9 × 105; maggots, 1.7 × 108 (day 10) and 7.2 × 108 (day 12); pupae, 6.2 × 107 (day 14) and 1.8 × 108 (day 16); and F1 adult houseflies, 1.2 × 108]. The mean ratio of tetA to 16S rRNA gene copies ranged from 4.2 × 10−5 to 8.5 × 10−3 per sample, except in adult houseflies (2.2 × 10−1 per sample).
Persistence of the ingested bacterial strain in the chicken model
After inoculation, E. coli strain TC7-1 was continuously detected in cecal samples from days 31 to 46 in groups 1 and 2 (Fig. 2). The strain was not detected in samples from the negative control group (group 3). After inoculation with maggots harboring E. coli strain TC7-1 on days 30 and 32, the mean concentration of bacteria in cecal droppings of the test chickens (group 1) first increased (1.6 × 105 CFU/g on day 31, and 3.6 × 107 CFU/g on day 33), and then gradually decreased (from 7.8 × 106 CFU/g on day 36 to 1.3 × 106 CFU/g on day 43) (Fig. 2A). On day 40, E. coli strain TC7-1 was not detected in one of the five cecal droppings and on day 43, it was not detected in two of the samples. The strain was detected in all cecal content samples on day 46 (1.8 × 104 CFU/g).
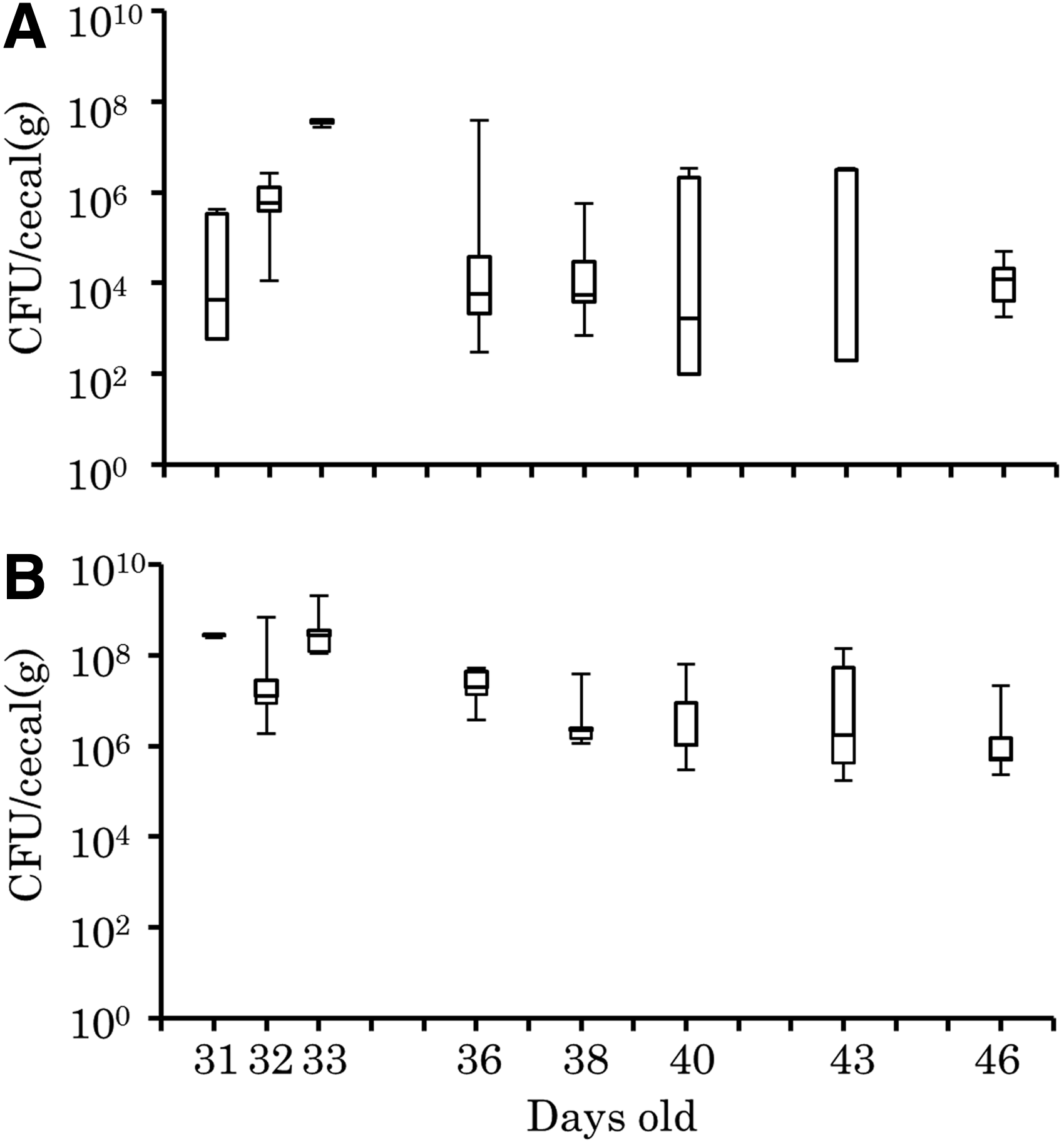
Maintenance of antimicrobial-resistant Escherichia coli strain TC7-1 in the intestine of orally inoculated chickens.
In group 2, the amount of E. coli strain TC7-1 in the cecal samples was 108 CFU/g following inoculation. This decreased gradually from 2.7 × 107 CFU/g on day 36 to 4.0 × 107 CFU/g on day 46 (Fig. 2B).
Discussion
Our study has revealed that antimicrobial-resistant E. coli can be vertically transmitted from adult houseflies to the next generation, despite previous studies reporting that the microbiota of the housefly intestine changes at different developmental stages.19–21 In particular, the morphology and microbiota of flies were dramatically changed by the metamorphosis during the developmental stage of the pupa20,22; however, we found that the constant of microbiota, including ingested bacteria, could be transmitted from maggots and pupae to the adult houseflies in the present study. The fly intestinal microbiota can be affected by various factors during the life cycle, including behavior and biological defense mechanisms.22,23 The persistence of ARB in our study suggests that ARB were selected and persisted in flies throughout their life cycle. In addition, as flies can travel >10 km owing to their strong flight capability, 24 they could act as a vector to widely disseminate ARB. 6 This is compounded by the persistence of ARB throughout the life cycle of the fly.
In this study, the transmission of ingested bacteria to housefly eggs and subsequent stages of the fly life cycle was observed despite surface sterilization of eggs and pupae before hatching or emergence, respectively. We found no bacteria in the final distilled water rinse after surface sterilization, suggesting that ingested bacteria had contaminated the internal chorion of the egg. In a previous study, the transmission of ingested bacteria from adult houseflies to eggs has been demonstrated by directly plating surface-sterilized eggs. However, the presence of ingested bacteria in surface-sterilized eggs was not evaluated by histology or transmission electron microscopy 8 and some bacteria, including ARB, may be transmitted to the progeny owing to strong adhesion to the egg surface. Therefore, in our study, although ingested bacteria were not detected from all of the housefly eggs, they could, however, transmit to the next generation.
A relatively consistent maintenance of ARGs (as assessed by the tetA/16S rRNA gene ratio) was detected in the microbiota of houseflies during a single generation in our study. Horizontal gene transfer of plasmids carrying ARGs between genera in the housefly intestine, including pathogenic and environmental bacteria, is therefore a potential risk that could contribute to the emergence of new ARB. 7 This would facilitate the maintenance of antimicrobial resistance plasmids in the environment. 25 However, antimicrobial resistance plasmids generally impose a fitness cost in the absence of antimicrobial selective pressure.9,26 Despite this, no significant difference in the viability of ARB carrying antimicrobial resistance plasmids and antimicrobial-susceptible bacteria in the fly intestine has been observed. 17 The constant presence of ARGs and their antimicrobial resistance in the housefly intestine may thus arise from a selective pressure in the fly.
In this study, chickens were infected with maggots that harbored antimicrobial-resistant E. coli, which persisted throughout the fly life cycle. This is supported by another study demonstrating that ARB were easily maintained in a broiler flock upon ARB infection. 27 ARB derived from livestock could therefore enter and circulate in the farm environment through transmission between internal and external livestock housing areas through flies and other insects.4,6 These results suggest that vector control in farms is vital to decrease the risk of dissemination and maintenance of harmful microorganisms, including ARB.
In addition, we found that chickens carried ARB until they were 46 days old after infection at 31 days of age. Broiler chickens are usually shipped at ∼50 days of age. 27 Therefore, the persistence of ARB in chicken intestines could lead to the presence of residual ARB on retail meat and subsequent transmission into humans through the food chain. 28 In addition, antimicrobial resistance plasmids could potentially be transferred to enteric bacteria residing in the chicken intestine, 29 raising the possibility of ARG dissemination across genera. To prevent the spread of ARB from livestock to humans through food or other routes, our study suggests that ARB and vector control in the farm environment is essential. 30
In conclusion, flies can act as both a vector for the transmission of ARB and as a reservoir throughout their life cycle. Our data also suggest that they are a vector for ARB transmission between internal and external livestock housing areas. On farms, especially in the poultry industry, livestock are bred in an all-in/all-out replacement system. Therefore, it is important to inhibit the influx of microorganisms and flies from the external livestock housing areas during the replacement period. Hygiene management, including the control of insects, may prevent the maintenance and circulation of ARB in the farm environment.
Footnotes
Acknowledgments
This work was supported by MEXT KAKENHI grant number 24590754. The authors thank Sumika Life Tech, Ltd. for providing houseflies (Musca domestica) (CSMA strain).
Disclosure Statement
No conflicts of interest to declare.