
Abstract
Purpose:
The more frequent reports of carbapenem-resistant Enterobacteriaceae have raised the alarm for public health. Apart from the production of carbapenemases, deficiency (decreased or loss of expression) of outer membrane proteins (OMPs) has been proposed as a potentially important mechanism of carbapenem resistance. The aim of the present study was to evaluate the contribution of the major OMPs to carbapenem resistance in Enterobacter aerogenes (CREA) isolates and also investigate the role of small RNAs (sRNAs) in inducing porin-associated permeability defects.
Materials and Methods:
The differential expression of OMPs was analyzed in four clinical CREA isolates. omp35 and omp36 genes were further investigated by whole-genome sequencing, induction of meropenem resistance, sRNA overexpression, OMP complementation assays, and reverse transcription-quantitative PCR.
Results:
All four isolates examined were deficient in omp35 and omp36. Functional restoration of these two genes confirmed their contribution to carbapenem resistance. The meropenem induction assay further revealed that porin deficiency plays a role in carbapenem resistance under antibiotic selection pressure. Single-point mutations in omp36 leading to premature stop codons were detected in two of the isolates. Elevated expression levels of the sRNAs micF and micC were detected in the other two porin-deficient isolates, which were predicted to be potential porin regulators from whole-genome sequencing. Overexpression of micF and micC downregulated the expression of Omp35 and Omp36, respectively.
Conclusions:
Porin deficiency plays an important role in carbapenem resistance among clinical E. aerogenes isolates under regulation of the sRNAs micC and micF. Furthermore, overexpression of micC and micF had a minor to no impact on carbapenem minimum inhibitory concentrations, and thus, the regulatory mechanism is likely to be complex.
Introduction
C
The most common and unequivocal carbapenem resistance mechanism involves the production of carbapenemases, although other mechanisms have been reported, including overproduction of β-lactamases, efflux pumps, porin deficiency, and changes in penicillin-binding proteins. 9 However, the role of porin deficiency in CREA remains controversial. In general, porin deficiency is considered to result in carbapenem resistance in conjunction with the production of β-lactamases.2,10 However, there have been isolated reports of porin deficiency alone leading to carbapenem resistance. CREA has over the past years become a major public health concern. 11 In E. aerogenes, Omp35 and Omp36 porin deficiency or loss has been considered to contribute to carbapenem resistance. 3 Although the mechanism of porin deficiency through gene mutation has been extensively studied in Enterobacteriaceae, other likely mechanisms leading to porin dysfunction remain to be investigated.12,13 One such mechanism is regulation of the expression of outer membrane proteins (OMPs) by relatively small RNAs (sRNAs) at the posttranscriptional level, which has been reported in Escherichia coli isolates under stimulation stress, including oxidative stress, and a change of osmotic pressure, pH, or temperature. 14 Regrettably, the regulation of OMPs by sRNAs in E. aerogenes has not yet been studied, and therefore, the relationship between sRNA regulation and carbapenem resistance remains unknown. To this end, the aim of this study is to first evaluate the contribution of the major outer membrane porins to carbapenem resistance of CREA isolates, and then determine the existence of an sRNA-related mechanism contributing to porin deficiency or loss.
Materials and Methods
Clinical isolates
A total of 164 clinical E. aerogenes strains were isolated from January 2013 to December 2015 at Huashan Hospital (Fudan University, Shanghai, China) and evaluated for carbapenem resistance. Antimicrobial susceptibility testing was performed by the broth microdilution method to determine the minimum inhibitory concentrations (MICs) of imipenem, meropenem, and ertapenem according to the Clinical and Laboratory Standards Institute (CLSI) 2017 guidelines, 15 and species confirmation was performed by amplification of the 16S rRNA gene. 16 Phenotypic testing indicated the presence of a carbapenemase in 14 isolates by using the combined disc test comprising meropenem alone and with ethylenediaminetetraacetic acid (EDTA) and phenylboronic acid (PBA). We confirmed the existence of blaKPC-2 carbapenemase genes by PCR amplification,17,18 except for four isolates (A4, A18, A19, and A23). These four isolates were further screened for β-lactamases (blaTEM, blaSHV, blaCTX-M, and blaOXA-1) and plasmid-mediated AmpC β-lactamases (blaAAC, blaFOX, blaMOX, blaDHA, blaCIT, and blaEBC) by PCR using previously reported primers. 17 The expression of chromosome ampc β-lactamases was confirmed by reverse transcription-quantitative PCR (RT-qPCR). 19 The role of the efflux pump was determined in the presence of the efflux pump inhibitor. 20
Pulsed-field gel electrophoresis
Pulsed-field gel electrophoresis (PFGE) of XbaI (Takara, Shanghai, China)-digested genomic DNA samples of E. aerogenes isolates was performed according to the instruction manual. The DNA fragments were separated by use of a CHEF-Mapper XA PFGE system (Bio-Rad). Electrophoresis was performed for 20 hr at 14°C with pulse times ranging from 3 to 40 sec at 6 V/cm. The DNA fingerprints generated were analyzed according to the criteria proposed by Tenover et al. 21
Meropenem resistance induction
Three carbapenem-susceptible isolates (S1, S2, and S3; meropenem MICs of 0.03 mg/L) were grown overnight at 37°C, and the culture was diluted in Luria-Bertani (LB) broth with a serially increasing concentration of meropenem. The initial concentration of meropenem was equal to 1/2 MIC and was then doubled every 24 hr. 22 The overnight cultures of the three isolates were plated on LB plates containing the same amount of meropenem with LB broth. Mutants showing resistance phenotypes were randomly selected from the plates after incubation at 37°C for 24 hr and then streaked onto new LB plates. Antimicrobial susceptibility testing was then performed again by the broth microdilution method.
Analysis of OMPs
OMPs were isolated according to the rapid procedure proposed by Carlone et al., 23 and the expression of outer membrane porins was confirmed by sodium dodecyl sulfate/polyacrylamide gel electrophoresis (SDS-PAGE) analysis. Porins were loaded onto a 12% SDS-polyacrylamide gel. After a 150-min electrophoresis of 80 V by Mini-PROTEAN Tetra (Bio-Rad, China), the membrane was stained with 0.1% Coomassie brilliant blue (Sangon, Shanghai, China).
The coding sequences of omp35 and omp36, with their respective promoter regions, were amplified as previously described 12 and aligned with the respective sequences of the E. aerogenes strain NCTC 10336 (GenBank accession no. KT894105.1 and KT894106.1, respectively).
Whole-genome sequencing to predict the sRNAs regulating OMP expression
We sequenced the whole genome of the carbapenem-resistant isolate A4 to predict the sRNAs regulating OMP expression. Gene predictions were carried out using the most popular ab initio methods, Glimmer (v3.02) and Prodigal (v2.6.2). The consensus set of genes (with the same start or stop positions in parent scaffolds) predicted by both tools was chosen as candidate gene models. In addition, Infernal (v1.1.1) and CMs from the Rfam database were used to identify rRNAs and sRNAs with default parameters. We combined the sRNA prediction results with known sRNA genes from sRNAMap and the sRNATarBase.v2.0 database as potential regulators. After confirmation of the genomic location of OMP genes (omp35 and omp36) in the sequenced strain, we extracted 1,000 bp upstream and downstream of the gene separately as the presumed regions regulated by sRNAs. All of the sRNAs selected were aligned to the target region using blastn with parameters “-task ‘blastn-short’ -evalue 1e-3,” and alignment blocks longer than 10 bp were considered to be highly confident regulatory regions. MiRanda (version 3.3a) was implemented to predict the sRNA–RNA interaction at the target regions extended with the length of sRNAs on both ends.
OMP complementation assays and sRNA overexpression
The OMP genes omp35 and omp36, and the predicted regulatory sRNAs micF and micC were amplified from a clinical carbapenem-susceptible isolate (S3) with a meropenem MIC of 0.03 mg/L. The omp35 and omp36 genes were cloned into pBad33, an arabinose-inducible vector with chloramphenicol resistance, yielding the recombinants pBad33-omp35 and pBad33-omp36, respectively. The micF and micC genes were cloned into pHSG396, a high expression vector with chloramphenicol resistance, yielding the recombinants pHSG396-micF and pHSG396-micC, respectively. For both complementation and overexpression, competent cells of the clinical isolates were prepared according to standard methods, 24 and recombinant plasmids were introduced by electroporation. Transformants were selected on LB agar containing 50 mg/L of chloramphenicol. The expression of Omp35 and Omp36 in the transformants was induced by 0.2% arabinose (weight/volume), and the expression of all genes was confirmed by real-time PCR and SDS-PAGE analysis.
Reverse transcription-quantitative PCR
RNA was extracted by the QIAgen RNA kit (QIAgen, Shanghai, China) and then immediately reverse-transcribed using the TaKaRa cDNA Synthesis kit (Takara, Shanghai, China). PCR was performed using the QIAgen real-time PCR kit (QIAgen, Shanghai, China) to determine the transcription levels of omp35, omp36, the multidrug efflux pump genes acrB, micC, and micF relative to the level of the housekeeping gene rpoB, according to a previously described protocol. 25 Primer sequences are given in Table 1.
Results
Clinical E. aerogenes isolates
In total, 18 of the 164 E. aerogenes isolates were found to be resistant to carbapenems. blaKPC-2 carbapenemase genes were detected in 14 of these isolates, and therefore, we proceeded with further investigation of the remaining 4 (A4, A23, A18, and A19). The carbapenem MICs of these isolates are shown in Table 2. Preliminary analyses showed that efflux pumps have no contribution to the resistance. Only one isolate harbored the AmpC cephalosporinase gene (blaDHA gene). No overexpression of chromosome AmpC and no β-lactamase genes were detected in the remaining three isolates. Three distinct PFGE groups (types A–C) were observed among the four isolates. A18 and A19 belonged to group A, and A4 and A23 belonged to group B and C, respectively (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/mdr).
Data represent minimum inhibitory concentrations (mg/L).
A4, A18, 18, A23: clinical isolates; S2R, S3R: induced isolates.
ETP, ertapenem; IPM, imipenem; MEM, meropenem.
OMP analysis
SDS-PAGE analysis revealed diverse OMP profiles among the four isolates (Fig. 1). Omp35 loss and decreased expression of Omp36 were observed in isolates A4 and A23. Decreased expression of Omp36 was observed in isolates A18 and A19. Sequencing revealed that omp36 was inactivated due to a point mutation in isolates A18 and A19, creating premature stop codons. No mutation in omp35 or omp36 was found in the other two isolates (A4 and A23).
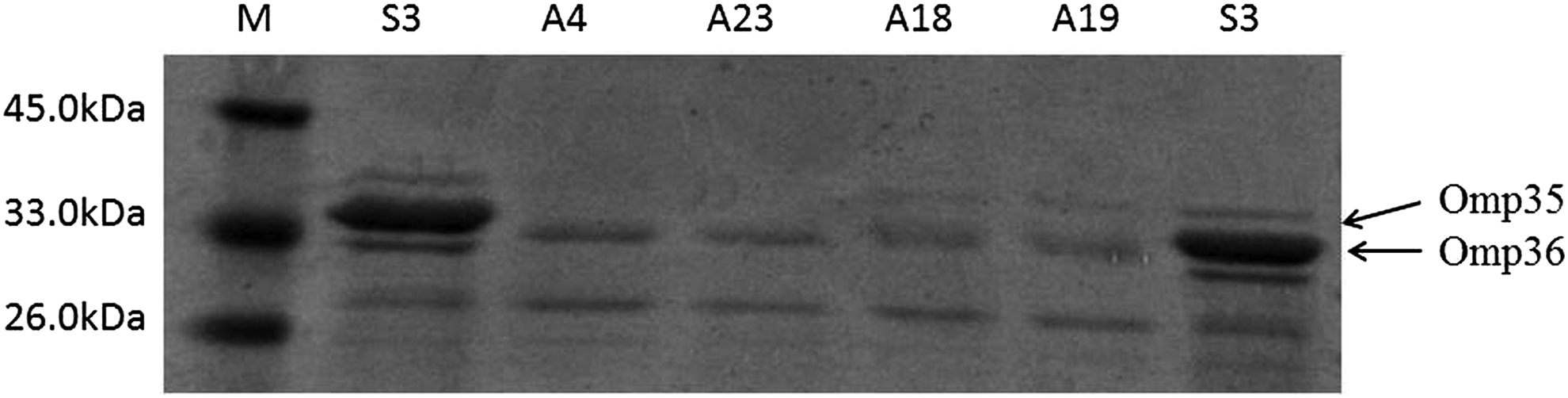
OMP profiles of four clinical Enterobacter aerogenes isolates on a 12% SDS-polyacrylamide gel. The last lane shows the molecular weight marker. S3 is a carbapenem-susceptible clinical isolate. OMP, outer membrane protein; SDS, sodium dodecyl sulfate.
Functional restoration of Omp35 and Omp36
Restoration of Omp35 function in A4 and A23, and of Omp36 function in A18 and A19, resulted in two- to fourfold decreases in the MICs of carbapenems (Table 2). SDS-PAGE analysis confirmed that the missing Omp35 and Omp36 were effectively restored in all transformants (Fig. 2).
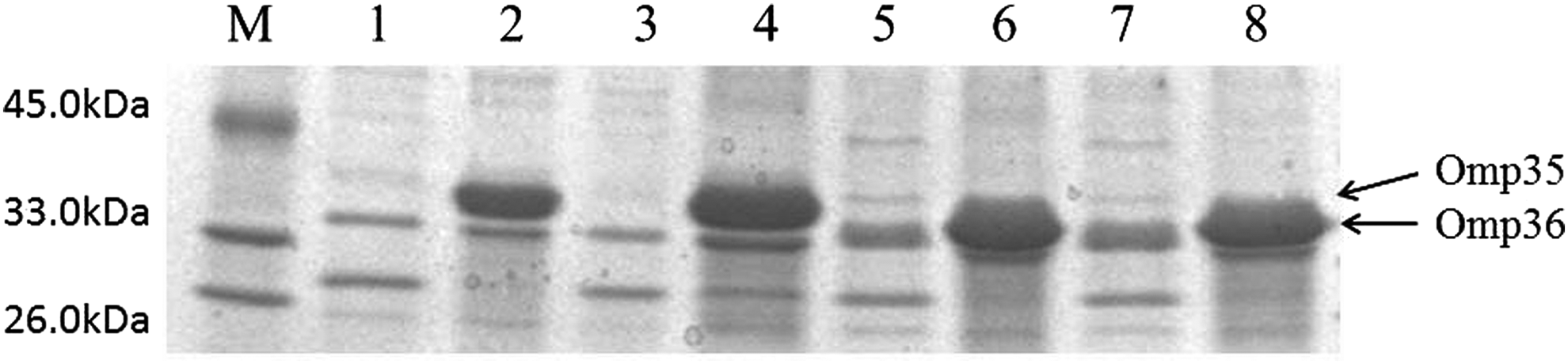
OMP profiles of strains with functional restoration of Omp35 and Omp36. Lane M: protein markers. Lanes 1–8: A4, A4 with functional restoration of Omp35; A23, A23 with functional restoration of Omp35; A18, A18 with functional restoration of Omp36; A19, A19 with functional restoration of Omp36.
In vitro selection of mutants with carbapenem resistance
After 13 days of serial passaging (every 24 hr), the meropenem MIC of the S2 mutant increased from 0.03 to 0.25 mg/L, which was designated S2R. The meropenem MIC of the S3 mutant increased from 0.03 to 8 mg/L, which was designated S3R. However, the S1 isolate maintained a low meropenem MIC throughout the experiment. The MICs of the three carbapenems for isolates S2R and S3R are listed in Table 2. SDS-PAGE analysis revealed loss of the porin Omp36 in S2R, and loss of both Omp35 and Omp36 in S3R. In isolate S3R, the omp35 sequence had a 1-bp deletion after 259 bp and the omp36 sequence had a point mutation at 962 bp, both of which resulted in a premature stop codon. In isolate S2R, the omp36 sequence had multiple single-base insertions, deletions, and substitutions. The similarity of the omp36 sequence between S2R and the parental isolate S2 was only 96%.
Genome sequencing of a carbapenemase-negative and porin-deficient isolate
The whole genome of isolate A4 was sequenced to identify potential sRNAs regulating OMP expression in E. aerogenes. The raw sequence data have been submitted to the SRA database under project ID PRJNA350242. According to the genome sequencing results and compared with the existing gene database, sRNATarBase.v.2.0 and sRNAMap predicted that the sRNAs micF and micC might regulate Omp35 and Omp36 porin expression (Table 2 and Fig. 3).
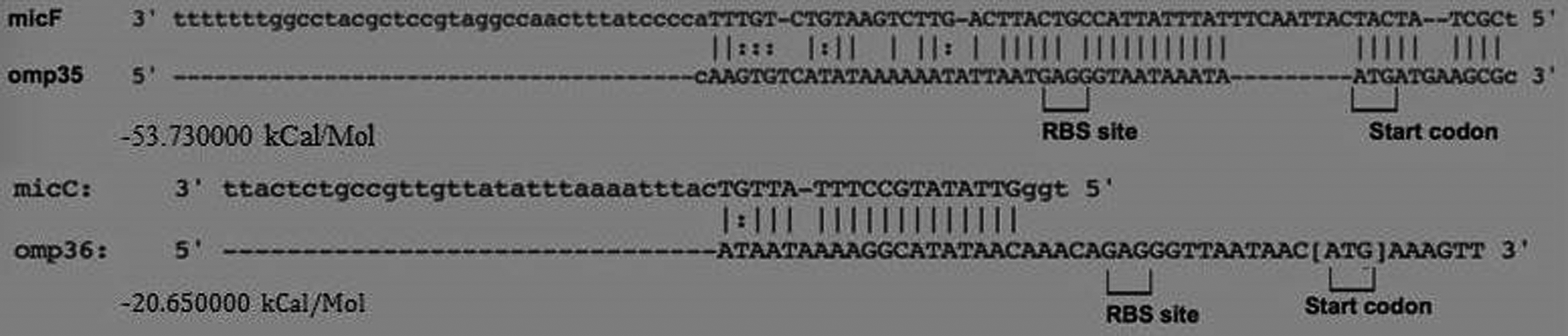
Expression of OMPs and sRNAs. sRNA, small RNA.
RT-qPCR of OMP genes and predicted sRNAs
Compared with the susceptible control strain, the transcription levels of omp36 were fivefold lower in isolates A18 and A19. The transcription levels of omp35 were 13- and 36-fold lower, and those of omp36 were 2–3-fold lower in the A4 and A23 isolates. The micF gene expression level of A4 was 20-fold higher than that of the susceptible control, and the micC gene expression levels of A4 and A23 were 13-fold higher than that of the susceptible control (Fig. 4). The transcription levels of acrB were equivalent between carbapenem-resistant and carbapenem-susceptible isolates.

Alignment block between sRNAs and OMP genes.
Overexpression of micF and micC
Real-time PCR confirmed that micF and micC were overexpressed in all transformants. In the micC-overexpressing isolates, the micF expression level was significantly decreased (Supplementary Fig. S2). SDS-PAGE analysis revealed that micF overexpression resulted in the loss of Omp35 expression, and micC overexpression resulted in downregulated expression of Omp36 (Fig. 5). Furthermore, elevated Omp35 expression was observed in micC-overexpressing isolates. The overexpression of micC resulted in a twofold increase in the MICs of carbapenems, whereas the MICs of carbapenems were maintained when micF was overexpressed (Supplementary Table S1).
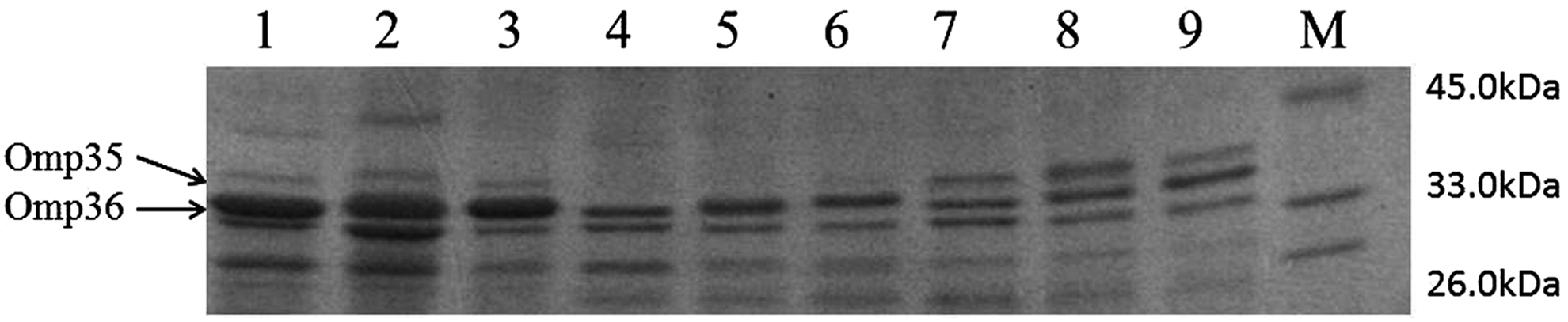
OMP profiles of sRNA-overexpressing isolates. Lanes 1–3: S1, S2, and S3, three clinical carbapenem-susceptible isolates. Lanes 4–6: S1-micF, S2-micF, and S3-micF, micF overexpression isolates. Lanes 7–9: S1-micC, S2-micC, and S3-micC, micC overexpression isolates. Lane M: protein marker.
Discussion
Resistance to carbapenems among E. aerogenes is a serious and growing clinical problem. The most common mechanism of carbapenem resistance is the presence of carbapenemase producing genes. Reduced susceptibility to carbapenems has been attributed to permeability deficiency in conjunction with β-lactamase production (ESBLs or AmpC-type enzymes). 3 Porin loss alone has also been implicated in causing carbapenem resistance,19,26 however, in Enterobacteriaceae, the ability of porin deficiency alone remains a matter of debate. In this study, we detected the absence or reduced expression of two major porins (Omp35 and Omp36) in two CREA isolates, and reduced expression of the porin Omp36 in another two isolates. Several previous studies suggested that Omp36 plays a more important role than Omp35 in carbapenem resistance,2,27 which was supported in the present study since the carbapenem MICs of Omp36-deficient isolates did not change depending on the presence or absence of Omp35. However, functional restoration of Omp36 or Omp35 resulted in two- to fourfold decreases in the MICs of carbapenems. We concluded that although Omp36 deficiency could contribute to carbapenem resistance without loss of Omp35, restoration of Omp35 expression could also enhance carbapenem susceptibility in case of deficiency of both these major porins. Thus, Omp35 loss deserves to occupy an important position in carbapenem resistance. We found that loss or deficiency of these two major porins in combination with AmpC enzyme could lead to a high level of carbapenem resistance. Moreover, in vitro carbapenem resistance induction showed loss of one or both major porins in both mutant isolates S2R and S3R, further highlighting the importance of porin loss in carbapenem resistance. Thus, CREA can occur under a condition of porin deficiency in an environment of carbapenem pressure.
Most cases of porin deficiency were shown to result from a single-base insertion, deletion, or substitution in the coding sequences, leading to premature translational termination, 12 which was further confirmed in the present study. In the clinical isolates A18 and A19 and in the induction isolates S2R and S3R, premature stop codons were found in the omp35 and/or omp36 sequences. Surprisingly, light bands of Omp36 were still detected despite the presence of premature stop codons in isolates A18 and A19. We speculated that this band was due to formation of segmental porins because the stop codon appeared at a rearward position. By contrast, the A4 and A23 isolates had intact porin genes, with no obvious Omp35 porin expression in the outer membrane, while the expression of Omp36 was substantially reduced. Given the previous finding of the sRNA-mediated expression of OMPs at the posttranscriptional level in E. coli, 14 we supposed a similar mechanism of sRNA regulatory OMP expression in these two CREA isolates.
Isolate A4, with an intact porin gene but no Omp35 in the outer membrane, was chosen for further whole-genome sequencing analysis. The sequencing result and sRNA prediction analysis revealed a >10-bp complementary binding block between the omp35 and micF genes, as well as between the omp36 and micC genes, which was supported by real-time PCR. Moreover, the isolate A4 showed overexpression of micF and micC, whereas A23 only showed overexpression of micC. A study in E. coli demonstrated that micF downregulated Omp35 porin expression and micC downregulated Omp36 expression. 28 Indeed, overexpression of micF and micC confirmed that this sRNA regulation mechanism also operates in E. aerogenes. In the micC overexpression isolates, micF expression was markedly decreased and Omp35 porin expression was increased. This suggests an as-yet-unidentified regulation mechanism between micC and micF. However, the overexpression of micF did not increase the MICs of carbapenems for the selected isolates, whereas micC overexpression resulted in twofold increases of the MICs. Omp35 loss was previously shown to be insufficient to cause carbapenem resistance 2 ; indeed, the MICs were maintained when micF was overexpressed. Given that micC overexpression led to reduced Omp36 expression and elevated Omp35 expression, we suggest that the overexpressed Omp35 partially compensated for the pore function of Omp36, resulting in no obvious change in the MICs.
In summary, we identified porin deficiency as an important mechanism in CREA; however, this appears to only lead to a low or intermediate degree of carbapenem resistance. Furthermore, there may be different mechanisms by which porin deficiency contributes to carbapenem resistance. Mutation in coding sequences is one major factor, and sRNA regulation might also play an important role. Although we identified that the sRNAs micF and micC could downregulate the expression of Omp35 and Omp36, respectively, in E. aerogenes, overexpression of the two sRNAs failed to alter the MICs of carbapenems significantly. This suggests a complicated regulatory network influencing carbapenem susceptibility. Thus, the detailed regulatory mechanism of sRNAs requires further study.
Footnotes
Acknowledgment
This work was supported by the National Natural Science Foundation of China (grant no. 81401699 to F.H.).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.