
Abstract
Eleven clinical Klebsiella pneumoniae fluoroquinolone-resistant isolates were tested to access the potential of adjuvant therapies to reduce antimicrobial resistance using fixed concentrations of the chemosensitizers chlorpromazine (CPZ), thioridazine (TZ), phenylalanine-arginine-β-naphthylamide (PAβN), and 1-(1-naphthylmethyl)-piperazine-(NMP) with varying concentrations of antimicrobial agents nalidixic acid (NAL), ciprofloxacin (CIP), moxifloxacin (MXF), tetracycline (TET), and chloramphenicol (CHL). Ethidium bromide dye was used together with the chemosensitizers to investigate permeabilization effects. NMP was assessed for its capacity to reduce the mass of biofilm alone and in combination with CIP and MXF. Of the selected chemosensitizers, NMP exhibited the greatest capacity to reverse resistance and inhibit efflux, based on the concentrations tested. Susceptibility to antimicrobial agents including (fluoro)quinolones, TET, and CHL were found to be increased in the presence of NMP, in a concentration-dependent manner. PAβN also demonstrated similar effects when combined with the chemosensitizers tested. In the case of half of the isolates studied, NMP alone reduced preformed biofilm biomass. Combinations of latter along with CIP or MXF were also found to reduce the mass of preformed biofilm, in the case of only some of the bacterial isolates. The capacity of NMP to reduce antimicrobial resistance could be of relevance as a strategy to limit bacterial colonization on abiotic surfaces.
Introduction
A
K. pneumoniae is commonly associated with hospital-acquired infections such as bloodstream infections, respiratory and urinary tract infections in immunocompromised patients.8,9 This bacterium has also been associated with community-acquired infections with more severe cases being documented in otherwise healthy individuals leading to pyogenic liver abscesses with high mortality rates. 10 After Escherichia coli, this pathogen is the second most commonly occurring etiological agent reported in cases of urinary tract infection. 11 One of the phenotypic features that lends support to this observation is the bacterium's ability to form biofilms, particularly on abiotic surfaces and indwelling medical devices such as catheters. 12 This sessile state confers additional resistance, to the action of antimicrobial compounds, further narrowing the chemotherapeutic options available for subsequent intervention. 13
Currently, there are few new antimicrobial compounds becoming available for clinical use. Therefore, to maintain the efficacy of the existing arsenal of agents, new strategies to preserve their action and to counter resistant bacteria are required. The use of combination therapy with carefully selected chemosensitizers, also known as efflux pump inhibitors (EPIs), is one potential option. The later strategy is designated as an adjuvant therapy and some studies report interesting results in overcoming innate resistance in bacteria of importance to human health. 14
Chemosensitizers are structurally diverse compounds with the capacity to act on different biochemical pathways. Some of these compounds are of vegetable origin such as reserpine, which was isolated from the roots of Rauwolfia vomitoria and demonstrates activity against Bacillus subtilis 15 ; other compounds of animal origin such as EA-371α and EA-371δ isolated from Streptomyces exhibit an inhibitory effect on MexAB-OprM efflux pump activity in Pseudomonas aeruginosa. 16 Alternatively, these compounds could be rationally designed making use of the new bioinformatics tools, as in the case of MBX231917,18 that acts as an inhibitor of resistance-nodulation cell division-type efflux pumps. Chemosensitizers can also be synthetic in nature as in the case of the psychotropic drug thioridazine (TZ) or the neuroleptic drug pimozide. 19 The arylpiperazine, 1-(1-naphthylmethyl)-piperazine (NMP) is a synthetic molecule reported to be effective in reversing fluoroquinolone resistance in resistant isolates of E. coli and other Enterobacteriaceae.20–22 The same ability to reverse resistance was also demonstrated for phenylalanine-arginine-β-naphthylamide (PAβN).23,24 The mechanism by which these compounds act remains to be elucidated. Anecdotal evidence suggests that they act as an inhibitory substrate for one or more efflux pumps with the capacity to bind strongly to the substrate/drug binding pocket thereby blocking its activity and leaving the antimicrobial compound available in the bacterial cytoplasm to exert its full effect. 25
In this study the ability of four chemosensitizers—including two phenothiazines: chlorpromazine (CPZ) and TZ; PAβN and NMP, were studied for their capability to reverse drug resistance, when used in combination with different classes of antimicrobial agent. The study focused on multidrug-resistant (MDR) K. pneumoniae of clinical origin.
Materials and Methods
Chemical compounds
Müeller-Hinton (MH) broth and MH agar, phosphate-buffered saline (PBS) and all the powder antibiotics/antimicrobial compounds, with the exception of moxifloxacin (MXF) and the chemosensitizers, TZ, CPZ, PAβN, and NMP, were obtained from Sigma (Ireland). MXF was purchased from Santa Cruz Biotechnology (Spain). Stock solutions were prepared according to the manufacturers' directions.
Bacterial isolates
Eleven MDR K. pneumoniae clinical isolates obtained from a tertiary teaching hospital (in Dublin, Ireland) 26 and the MDR K. pneumoniae MGH 78578 ATCC™700721 was included as a control in this study. E. coli ATCC™25922 was also included as reference for the antibiotic susceptibility testing.
Chemosensitizers and the measurements of minimum inhibitory concentrations and minimum bactericidal concentrations
The minimum inhibitory concentrations (MICs) for the chemosensitizers TZ, CPZ, PAβN, and NMP along with all of the antimicrobial agents tested including ciprofloxacin (CIP), chloramphenicol (CHL), MXF, nalidixic acid (NAL), and tetracycline (TET), were determined by the two-fold broth microdilution method in 96-well microtiter plates.
Briefly, bacterial overnight cultures were diluted in sterilized PBS to ∼105 CFU/mL and 5 μL were inoculated into a 96-well of the plate containing 100 μL of each chemosensitizer and each of the antimicrobial compounds studied at concentrations prepared, from twofold serial dilutions in MH broth medium. The concentration ranges used (high-to-low) for the chemosensitizers were 400–0.78 μg/mL for TZ and CPZ; 1,000–1.95 μg/mL for PAβN and NMP. For the antimicrobial compounds, the concentration ranges were 512–1 μg/mL, with the exception for MXF wherein the concentration used ranged from 128 to 0.25 μg/mL. Dimethyl sulfoxide (DMSO) was used as a control to test for any solvent effects in this MIC assay. Plates were incubated at 37°C for 16–18 hr and the MIC values recorded thereafter.
Measurement of the minimum bactericidal concentration (MBC) values was performed in MH broth. Five microliters were aliquoted from the latter 96-well plate and reinoculated into a fresh sterile 96-well plate containing fresh MH media. These plates were incubated at 37°C and the results recorded after 16–18 hr.
Ethidium bromide accumulation assay
Intracellular accumulation of the fluorescent dye, ethidium bromide (EtdBr) can be measured using fluorometry. This is a convenient way in which to measure bacterial efflux/membrane permeability in presence of modulating compounds, such as chemosensitizers. 27 Briefly, overnight cultures of K. pneumoniae were centrifuged and washed three times with PBS and OD600nm adjusted to 0.6. EtdBr was added to the samples to a final concentration of 50 μM.
Two hundred microliters of the adjusted cells along with EdtBr were aliquoted into a black bottomed 96-well plate. The selected chemosensitizers were included in the plate at defined concentrations both in the presence or absence of glucose (50 mM). TZ and CPZ were added at final concentrations of 25, 50, and 100 μg/mL; PAβN final concentrations were 10, 20, 100, 250, and 500 μg/mL; and NMP was added at 62.5, 125, and 250 μg/mL. Heat-inactivated bacteria (10 min at 90°C) were used as a control for maximum fluorescence and cells in the presence of PBS alone were used as a background signal, carbonyl-cyanide m-chlorophenylhydrazone (CCCP) was added to a final concentration of 50 and 100 μM (in the presence and absence of 50 mM glucose).
Fluorescence was measured in a Fluoroskan Ascent FL (Thermo Scientfic, Dublin, Ireland) with excitation and emission wavelengths of 518 and 606 nm, respectively. Fluorescence was acquired in cycles of 60 sec, for a total period of 50 min. The compounds were tested in duplicate for this experiment. Duplicates were averaged and plotted using Rstudio (www.rstudio.com) and compared among the isolates based in their average fluorescence units.
Reversal of antimicrobial resistance in planktonic grown cells
To reverse the resistant phenotype of the study isolates, a fixed concentration of each of the selected chemosensitizers was used in combination with each of the antimicrobial compounds. TZ and CPZ were used at 50 μg/mL, PAβN at 20 μg/mL, and NMP was included at 125 and 250 μg/mL. The chemosensitizers were prepared at the specified concentrations in MH broth. Antimicrobial compounds CIP, CHL, MXF, NAL, and TET were diluted individually in each chemosensitzer prepared solution. All of the dilutions tested, were prepared by twofold broth microdilution in 96-well microtiter plates.
Overnight bacterial cultures were diluted in sterilized PBS to ∼105 CFU/mL. Aliquots of 5 μL were then transferred to each well of the 96-well plate that contained 100 μL of each compound mixture. The antibiotic concentration ranges used were 512–1 μg/mL for NAL, TET, and CHL. For CIP and MXF concentration ranges used were 64–0.0625 μg/mL. The solvent DMSO was also used in combination with the antimicrobial compounds and included here to test for any differential effects it might exert on bacterial growth. Plates were incubated at 37°C for 16–18 hr and MIC values recorded.
The determination of the MBC was performed in MH broth, as described earlier. Five microliters were collected from the MICs 96-well plate and reinoculated in a new 96-well plate with fresh MH media. The plates were once again incubated at 37°C and the MBC values obtained recorded after 16–18 hr.
Susceptibility of each isolate was interpreted using CLSI 2016 criteria (as outlined in the M100-S23 document). For MXF, the breakpoints used were those published by the European Committee on Antimicrobial Susceptibility Testing (EUCAST, 2016). All assays were carried out on three independent occasions for each drug-chemosensitizer combination.
Bacterial growth curves in the presence of NMP
Bacterial growth curves were performed by measuring the turbidity of K. pneumoniae in the presence of NMP in MH broth at 0, 125, and 250 μg/mL, respectively. Bacterial overnight cultures were adjusted in MH-NMP solution, at the above concentrations, to an OD600nm 0.005. A volume of 200 μL of each diluted study isolate was aliquoted to a 96-well plate and absorbance was measured at 37°C for a period of 24 hr using a Multiskan™ FC Microplate Photometer (Thermo Scientific, IE) at 620 nm. Each isolate was cultured independently on five different occasions.
Biofilm assay in the presence of NMP and fluoroquinolone compounds
Cultures of each K. pneumoniae study isolate were grown overnight and then adjusted to OD600nm 0.3 in M9 minimal media (NH4Cl [1.9 mM], Na2HPO4 [42.3 mM], KH2PO4 [22 mM], NaCl [8.56 mM], MgSO4 [2 mM], CaCl2 [0.1 mM], and glucose [0.1% w/v]). Following this step, each of these was dispensed (as 200 μL volumes) across a 96-well microtiter plate and incubated statically for 8 hr at 37°C. Plates were then washed with PBS to remove planktonic cells and fresh media was added containing defined concentrations of NMP (0, 250, 500, and 1,000 μg/mL). For the reversal of fluoroquinolone-resistant biofilms, CIP or MXF at the MBC was added in the presence and absence of 250 μg/mL of NMP. Plates were then reincubated statically for 16–18 hr at 37°C. The crystal violet assay was carried out as described previously. 26 This assay was repeated 3 times for each isolate and each assay was carried out over 10 technical replicates. Results were compared against the control well containing fresh M9 minimal media alone.
Statistical analysis
Biofilm data were processed individually for each time point and condition in Excel (Microsoft). Comparisons between conditions were performed using Prism 6 (Graphpad) using ANOVA analysis with Tukey's multiple comparisons test. p-Values of <0.05 were considered significant.
Results
Chemosensitizer characterization
Previously, a study collection of 11 K. pneumoniae of clinical origin and belonging to the clonal groups CG258 (n = 7), CG15 (n = 3), and CG147 (n = 1) were characterized. All were found to be resistant to extended-spectrum β-lactam compounds and fluoroquinolones. 26 The aim of this study was to explore strategies designed to reverse the resistance phenotype to several antimicrobial compounds, using selected chemosensitizers.
Several chemosensitizers along with the determination of their corresponding MICs and their MBCs are shown in Table 1. For the majority of these compounds MIC/MBC values recorded were determined within similar ranges with a maximum of two- or four-fold differences being recorded. The reference isolate K. pneumoniae MGH 78578 demonstrated a lower MIC for TZ and CPZ when compared with the majority of the clinical isolates in this study.
Minimal Inhibitory Concentration and Minimal Bactericidal Concentration
CPZ, chlorpromazine; MBC, minimum bactericidal concentration; MIC, minimum inhibitory concentration; NMP, 1-(1-naphthylmethyl)-piperazine; PAβN, phenylalanine-arginine-β-naphthylamide; TZ, thioridazine.
Assessment of membrane permeability and efflux activity using EtdBr
Study isolates were tested for their ability to accumulate EtdBr in the presence of each of the selected chemosensitizers. These compounds, also known as EPIs,22,24,28 can reduce the capacity of the bacterial cell to expel a broad range of chemically distinct substrates. Accumulation of EtdBr was measured, as a means of indicating the efficacy of each chemosensitizer to permeabilize the bacterial membrane. When the efflux pumping capacity of a bacterial cell or the membrane integrity is compromised, the level of fluorescence emitted will be higher, when compared to a control condition. All isolates were assayed in saline buffer in the presence and absence of glucose. Glucose functions to energize the bacterial cell and supports the activity of energy-dependent efflux systems.
Data obtained (Supplementary Figs. S1.1–S1.12; Supplementary Data are available online at www.liebertpub.com/mdr) highlighted the fact that all chemosensitizers had a general ability to destabilize the cell membrane, as recorded by increases in the measurement of fluorescence. The chemosensitizers CPZ, TZ, and NMP appeared to act in a concentration-dependent manner. The interaction between these chemosensitizers, as determined by this assay, and some of the bacterial isolates showed some differences.
More fluorescence was emitted when using CPZ, followed by TZ, NMP, and then PAβN (Supplementary Figs. S1.1–S1.12). The ability of the chemosensitizer PAβN to permeabilize the cell membrane, as measured by emitted fluorescence, was found to be lower in magnitude, when compared with the other chemosensitizers. The only noticeable effect observed was at a concentration of 500 μg/mL.
The presence of glucose in the medium reduced the emitted fluorescence signals, an effect that was more pronounced when the chemosensitizer TZ was used. CPZ activity was also affected in a similar manner, in particular for the lowest concentrations tested (25 and 50 μg/mL) for all isolates tested with the exception of K. pneumoniae CFS0366 and CFS0370, wherein the effect of glucose resulted in minimal changes in the fluorescence when using CPZ at 50 μg/mL. The same effect was noted with NMP, wherein the presence of glucose resulted in a significant reduction in fluorescence when at lower concentrations (62.5 and 125 μg/mL).
Important to note was the effect of the proton ionophore, CCCP that is known to interfere with the electrochemical gradient in Gram-negative bacteria reducing the production of ATP and increasing the membrane permeability.29,30 The interaction of this compound with K. pneumoniae also demonstrated an isolate-dependent effect with 41% (n = 5 isolates), showing resistance to the action of this compound.
Investigation of four selected chemosensitizers for their ability to reduce the antimicrobial resistance phenotype in K. pneumoniae of clinical origin
To study the effect(s) of these selected chemosensitizers on the susceptibility of the bacterial study isolates, fixed concentrations were used; TZ and CPZ were both used at 50 μg/mL, NMP at 125 and 250 μg/mL. PAβN was used at 20 μg/mL, a concentration that was lower than the MIC arising from toxicity of this compound to human cells. 31 In addition to the (fluoro)quinolone compounds (NAL, CIP, and MXF) evaluated in the presence of the latter chemosensitizers, two other agents, TET and CHL, both of which exhibit distinct resistance mechanisms of action were included.
Overall, when the effects of all chemosensitizers in combination with antimicrobial compounds were evaluated and compared at defined concentrations, NMP demonstrated a greater capacity to reverse resistance of the antimicrobial agents tested followed by PAβN (Table 2 and Supplementary Table S1).
Changes in MIC Values Among the Study Isolates of Klebsiella pneumoniae
Susceptible isolates were not tested in combination with the chemosensitizers (marked with a dash “-”). Statistically significant reductions in the mass of biofilm formed are in bold.
MICs determined according to the CLSI guidelines (CLSI, 2016).
MICs determined according to the EUCAST guidelines (EUCAST, 2016).
50 μg/mL.
20 μg/mL.
250 μg/mL
125 μg/mL.
CHL, chloramphenicol; CIP, ciprofloxacin; MXF, moxifloxacin; TET, tetracycline.
All clinical isolates were originally shown to be resistant to NAL (data not shown) and none of the chemosensitizers used could reverse this phenotype, with the sole exception of isolate K. pneumoniae CFS0364, wherein a susceptible phenotype was obtained when using CPZ. This isolate was characterized by the absence of any observable efflux-dependent mechanism conferring resistance to quinolones. 26 PAβN and NMP (at 250 μg/mL) were the only compounds capable of causing a reduction on the resistance phenotype, for the majority of isolates, of at least four- and six-fold, respectively.
Resistance to CIP and MXF followed the same pattern as for NAL, with NMP being the chemosensitizer that demonstrated the greatest fold-change recorded. Ten and 11 of the 12 study isolates, were found to revert to susceptibility. K. pneumoniae CFS0364 and CFS0369 expressed a resistance phenotype to CIP, and when CPZ was included, MIC when remeasured indicated that the isolate became susceptible. A similar effect was noted for K. pneumoniae CFS0367 and CFS0368 when PAβN was included. No major genomic or selected phenotypic differences were identified in these isolates that could further explain these observations. 26
K. pneumoniae CFS0363 expressed a higher MIC against CIP, and did not respond to the action of the chemosensitizer, in particular to NMP, when compared with the other study isolates. This former isolate was positive for both the oqxAB and kexD efflux pumps along with containing a plasmid-mediated qnrB1 gene.
In the case of TET not all of the study isolates were resistant to this compound. Therefore, only the intermediate and resistant isolates were selected for testing. The majority of the chemosensitizers used demonstrated limited ability to reduce the resistance phenotype with the sole exception of NMP wherein 8- to 12-fold changes in MIC were noted. When considering CHL resistance, both PAβN and NMP were the chemosensitizers where most of the changes in MIC values were recorded. Five of 11 isolates demonstrated a susceptible MIC, when the antimicrobial compounds were used in combination with the NMP and PAβN (Table 2).
The effect of the solvent DMSO, used to dilute NMP was negligible (data not shown) and did not contribute to these reductions. Data obtained from these laboratory assays suggest that, Klebsiella isolates appeared to respond to the action of NMP in a concentration-dependent manner. The action of PAβN was similar to that of NMP even though the compounds were used at different concentrations.
Measurement of the effect of NMP on bacterial growth
The chemosensitizer NMP proved to have a positive effect, reducing the MIC, when combined with certain antimicrobial compounds. To determine whether or not the effect exerted by NMP could cause any impact on bacterial growth, the ability of all 11 clinical isolates was assessed by standard growth assays in broth culture in the presence of 0, 125, and 250 μg/mL of NMP. Data obtained (Supplementary Fig. S2A–F) showed that at the lowest concentration of NMP used, 125 μg/mL, growth appeared to demonstrate a typical shaped curve with no significant changes being observed, with the exception of the isolate K. pneumoniae CFS0368 (Supplementary Fig. S2F). At 250 μg/mL of NMP, all the isolates demonstrated a modest extension of the lag phase, suggesting that bacteria require time to recover from the initial chemosensitizer stress. A stationary growth phase decrease was noted at this concentration suggesting that prolonged exposure to NMP could impair the ability for bacteria to maintain a normal duplication performance.
Study of the effect of NMP on preformed biofilms
Given the ability of NMP to reverse antimicrobial-resistant phenotypes, when combined with fluoroquinolones during planktonic growth along with its effects on stationary phase of bacterial growth (Supplementary Fig. S2) as determined by reductions in MIC values, the ability of NMP to reduce/eliminate preformed biofilms (formed after 8 hr growth) was tested. NMP was used at concentrations equivalent to 0.5× MIC (250 μg/mL), MIC (500 μg/mL) and 2× MIC (1,000 μg/mL) and due to its solubility, higher concentrations of NMP could not be used.
Data presented in Fig. 1 describe the action of NMP to reduce the mass of a preformed biofilm, at the concentrations tested. This effect appeared to be concentration dependent. At 0.5 × MIC, 33% (n = 4) of the isolates showed a statistically significant decrease in biomass, which was higher than 10%. Similarly, at the MIC concentration, 50% (n = 6) of these isolates showed a 10% decrease in biomass (p < 0.05). At 2× MIC concentrations, 67% (n = 8) of the isolates showed a statistically significant decrease in biofilm formation, higher than 70% (p < 0.0001) with two isolates that showed a decrease higher than 50% (p < 0.001).
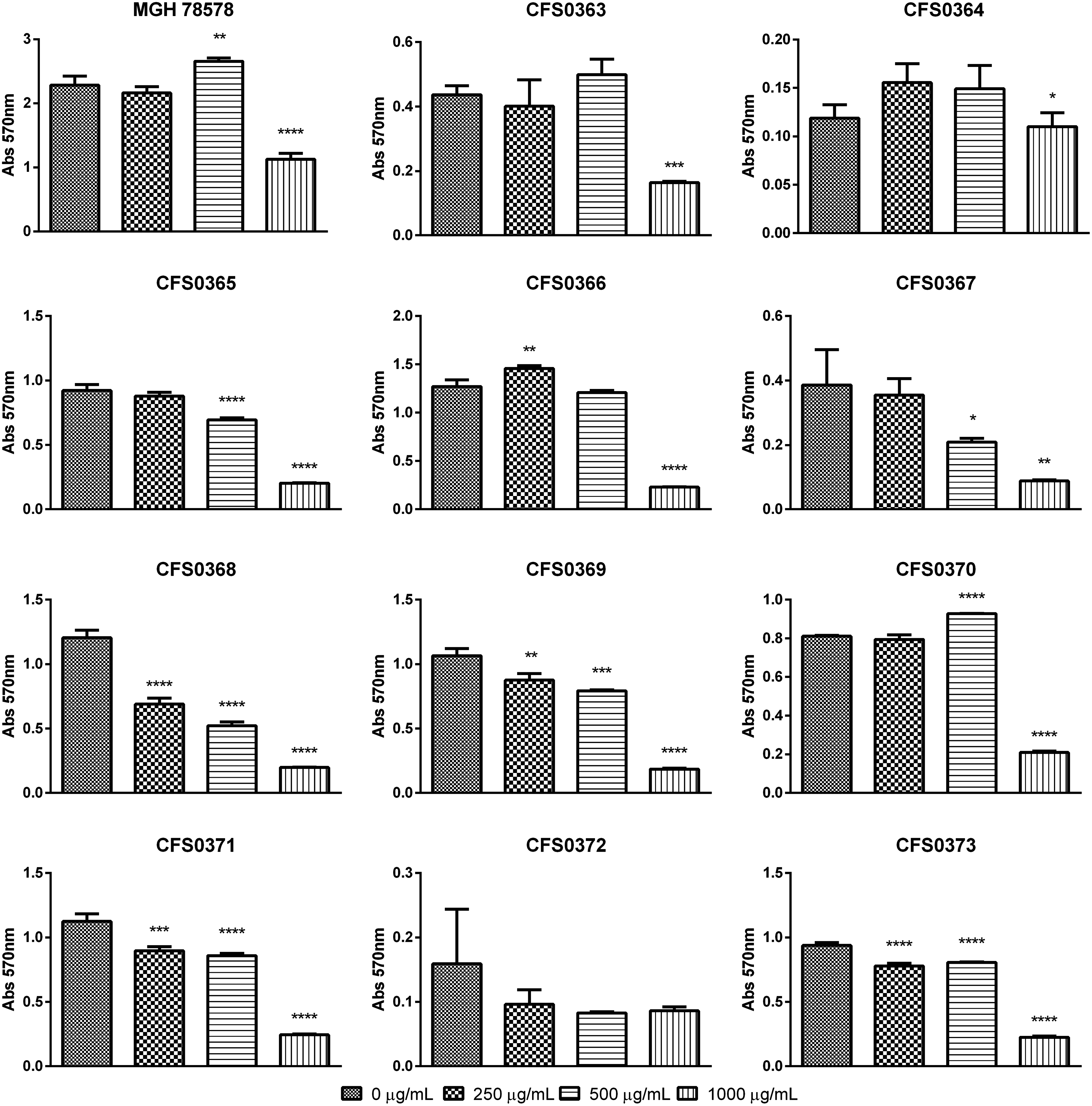
NMPs antibiofilm effect in preformed Klebsiella pneumoniae biofilms grown in M9 minimal media statically at 37°C for 16–18 hr. Biofilm biomass (expressed in OD570nm) was quantified by crystal violet staining. Each isolate was grown in different concentrations of NMP and was subsequently compared with the control (0 μg/mL NMP) using ANOVA analysis with Tukey's multiple comparisons: *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001. NMP, 1-(1-naphthylmethyl)-piperazine.
K. pneumoniae CFS0368 was found to respond greatest to NMP even at 0.5× MIC, a pattern also observed when its bacterial growth was tested in the presence of NMP (Supplementary Fig. S2F).
Fluoroquinolone-resistant biofilm eradication
A study of the effects of the combined action of NMP and fluoroquinolones, CIP and MXF in reducing biofilm formation was carried out. Data obtained were shown in Table 3 along with Supplementary Figs. S3 and S4. Initially, mature biofilms (formed after 8 hr) were grown in M9 minimal media in the presence of the antimicrobial compound alone at the MIC concentration and in combination with NMP at 0.5 × MIC concentration (250 μg/mL).
Biofilm Reduction Assays of Fluoroquinolone Resistance in Klebsiella pneumoniae
Statistically significant reductions in the mass of biofilm formed are in bold.
P < 0.05.
P < 0.01.
P < 0.001.
P < 0.0001.
CIP when used alone was found to reduce the mass of preformed biofilm in 9 of the 12 isolates studied, as determined by CV straining. Two of these isolates showed a statistically significant decrease that was calculated to be higher than 10% with a p < 0.01. The combinatory effect of CIP along with NMP showed that 58% of the isolates (n = 7) exhibited decreased levels of preformed biomass and which three isolates presented a statistical significant decrease that was calculated to be higher than 30%. In contrast, 42% of the study collection increased their biomass by more than 25% (p < 0.01) when in presence of the adjuvant combination.
MXF alone decreased preformed biomass in only one isolate (calculated as a 9.59% decrease) with statistical significance (p < 0.01). A statistically significant increase in biofilm biomass was also noted for two isolates of between 16% and 28%. The combinatory effect of MXF and NMP produced a significant decrease in preformed biofilm, which was calculated at being higher than 20% in five of the study isolates. Two isolates (K. pneumoniae CFS0371 and CFS0373) were able to double their biomass (p < 0.0001) in comparison with the control.
Overall, no common trend toward reduction in biofilm formation was identified when combining CIP or MXF at their MIC concentrations along with NMP at 0.5× MIC. Variability in biofilm formation appeared to be isolate specific.
Discussion
K. pneumoniae is now considered to be an important threat to public health due to its capacity to acquire and express a MDR phenotype. The latter is of particular concern with respect to extended-spectrum β-lactamase-based compounds, carbapenams, fluoroquinolones, and more recently colistin among others.32–34 In this study the potential to reverse some of these resistance phenotypes was explored. Fluoroquinolones are a commonly prescribed class of antimicrobial compound in the medical setting. 35 These drugs target the bacterial enzymes DNA gyrase and DNA topoisomerase IV. Resistance to fluoroquinolones is mainly associated with target gene mutations in gyrA, gyrB, parC, and parE 36 along with efflux mechanisms including AcrAB-TolC, 37 KexD, 38 and the plasmid-encoded oqxAB efflux system. 39 Other resistance mechanisms including plasmid-mediated functions have also been reported and which in some cases act to protect the enzyme from the drug. 40
To assess whether or not adjuvant therapy might be a useful strategy designed to preserve the efficacy of these compounds in clinical medicine, four selected chemosensitizers TZ, CPZ, PAβN, and NMP were studied in combination with NAL, CIP, MXF, TET, and CHL.
Data obtained in this study showed that all bacterial isolates studied exhibited similar MIC values for each of the four selected chemosensitizers. These findings are similar to values previously published for K. pneumoniae21,41,42 and other Gram-negative pathogens such as E. coli,20,22,43 Salmonella,44,45 Acinetobacter baumannii, 46 and Vibrio cholerae. 47
When used in combination with either CIP or MXF, NMP, at concentrations of 250 and 125 μg/mL, demonstrated a notable ability to reduce bacterial resistance in a dose-dependent manner with an average 10-fold decrease in MIC values, recorded for both drugs tested. Similar reductions in MIC values were also recorded when using lower concentrations of NMP in some Gram-negative bacteria.20–22,43,46,48 In contrast, PAβN was the chemosensitizer that demonstrated a greater potency with reductions in the order of four- to eight-fold recorded at a concentration of 20 μg/mL. The latter concentration was set at a lower level compared to the isolates MIC, due to the associated toxicity. 31
Smaller changes in MIC values were also observed with TZ and CPZ when tested in combination with CIP and MFX, [with the exception of two isolates (K. pneumoniae CFS0364 and CFS0369) that appeared to be more susceptible to the action of CPZ] and wherein resistance to NAL and CIP, respectively, was reversed completely. These results suggest that these compounds may be interacting in a bacterial isolate-dependent manner.
Two other antibiotics were also assessed to determine whether the observations made with NMP and fluoroquinolone-based compounds could be extended to other representatives of different drug classes: TETs function to inhibit protein synthesis by preventing the attachment of aminoacyl-tRNA to the 30S ribosomal acceptor (A) site 49 and CHL binds irreversibly to the receptor site on the 50S subunit of the bacterial ribosome, inhibiting peptidyl transferase, resulting in the inhibition of protein formation. 50 In both cases mechanisms of resistance associated with efflux pump activity have also been reported. 51 TET in combination with NMP when used at 250 μg/mL produced a 8- to 12-fold decrease in MIC values. CHL resistance was also positively affected by NMP and also by PAβN, both reversing the resistant phenotype in half the study isolates in this collection.
The effect of individual chemosensitizers on membrane permeability, when used at different concentrations, was assessed by measuring fluorescence emission, following the intracellular accumulation of EtdBr. Data showed different emission patterns. Chemosensitizers CPZ and TZ gave rise to increased dye accumulation followed by NMP. The latter effects appeared to function in a concentration-dependent manner.
Based on these data, it would seem reasonable to hypothesize that the chemosensitizers selected for study may exert their effects on the bacterial cell by different mechanisms. TZ and CPZ appear to exhibit similar effects, based on fluorescent measurements. In contrast, NMP produced reduced fluorescent signal emissions while inclusion of PAβN gave rise to even lower emission signals. Further, interaction of these chemosensitizers with bacterial cells also appeared to demonstrate some strain-specific interactions, with some isolates showing greater susceptibility toward specific chemosensitizers or chemosensitizers/drug combinations. Several possible reasons may account for this and could include differences in the expression of efflux pumps, porins, lipopolysaccharide (LPS), or capsular polysaccharide (CPS) modifications. Another contributing factor to consider is pH. All experiments were performed at neutral pH, the interaction of these chemosensitizers with antimicrobial compounds could be potentiated at higher or lower pH.52,53
Overall, based on the concentrations tested the activity of NMP was capable of reversing MDR phenotypes in K. pneumoniae. These data suggest that NMP could be a potential substrate for a number efflux pumps apart from the known target AcrB,54–56 as it was also able to reduce TET and CHL resistance. Membrane permeabilization effects, as evidenced earlier could also point to a complementary activity of NMP, as seen previously for the chemosensitizer PAβN.24,57
The activity of NMP alone also demonstrated a positive effect in reducing the biomass of an 8 hr preformed biofilm. The effect of NMP was shown to be concentration dependent. Previous publications also reported these effects for NMP and these were concomitant with reductions in TET-resistant E. coli when contained in a biofilm. 58 When combined with antimicrobial compounds, 25% and 42% of the isolates reduced their biomass in the presence of CIP and MXF, respectively.
The combinatorial effect of the selected antimicrobial agents and NMP appeared to increase the mass of biofilm as measured by CV staining, in some cases, a result that contradicts the findings from planktonic experiments. Use of mature biofilms is known to trigger the activation of certain resistance mechanisms in the presence of the antimicrobial compound and this signaling could have a direct impact on how these assays perform.59–61 The biofilm assays reported in this study also showed some strain-specific responses to the action of the chemosensitizer/antimicrobial compounds combination, suggesting once more that differential expression of components such as porins, efflux pumps, LPS, or CPS could play a hitherto underappreciated ole.
In conclusion, the mechanism of action of NMP is not fully elucidated and furthermore due to its serotonin antagonistic characteristics this compound is considered to be currently too toxic for clinical use. 62 Nonetheless, understanding the mechanisms of action of NMP is a useful first step toward the rational design of chemosensitizers similar in structure, but which are less toxic to humans. Currently, and based on these findings, NMP could be used to treat abiotic surfaces to mitigate biofilm formation and contribute to a reduction in the ability of MDR K. pneumoniae to colonize these ecological niches in the clinical setting.
Footnotes
Acknowledgments
The authors would like to acknowledge the financial support through the research grant 11/F/051 provided by the Department of Agriculture, Food and the Marine (DAFM), Ireland. Similarly, the Food Institutional Research Measure (FIRM) Network & Team Building Initiative 2006 is also acknowledged.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.