
Abstract
Aeromonas spp. are opportunistic pathogens related to multiple infectious diseases in ornamental fishes. In the present study, the antimicrobial susceptibility, resistance genes, and integrons of 65 goldfish-borne Aeromonas spp. were evaluated. The isolates were identified as A. hydrophila (n = 30), A. veronii (n = 32), and A. punctata (n = 3) by gyrB sequencing. The antimicrobial susceptibility testing of the isolates designated that most of the isolates were resistant to amoxicillin (100.00%), nalidixic acid (100.00%), ampicillin (98.46%), tetracycline (92.31%), rifampicin (86.15%), and cephalothin (61.54%) and each of the isolates showed multiple antimicrobial resistance phenotype (resistant to ≥3 classes of antimicrobials). PCR amplification of antimicrobial resistance genes revealed that the plasmid-mediated quinolone resistance gene, qnrS, was the most prevalent (73.85%) among the isolates. The other antimicrobial resistance genes were detected in the following proportions: qnrB (26.15%), aac(6′)-Ib-cr (4.60%), tetA (16.92%), tetE (21.54%), aac(6′)-Ib (29.23%), and aphAI-IAB (7.69%). The IntI gene was found in 64.62% isolates, and four class 1 integron gene cassette profiles (incomplete dfrA1, catB3-aadA1, dfrA1-orfC, and qacE2-orfD) were identified. These data suggest that goldfish-borne Aeromonas spp. serve as a reservoir of antimicrobial resistance genes and class 1 integrons.
Introduction
A
Goldfish (Carassius auratus) are among the most popular freshwater ornamental fish in the world, because of their attractive coloration and high adaptability to the environment. Due to the high demand, intensive culture of goldfish has been expanded, which led to a high incidence of infectious diseases. In Asia, Aeromonas spp., Flavobacterium spp., and Edwardsiella spp. are causing a huge economic loss in the ornamental fish industry by disease outbreaks. 3 Among them, Aeromonas infections are predominantly observed in goldfish. 4
Aeromonas species are the autochthonous microflora of freshwater, sewage, and brackish water environments, which have been also considered as the important pathogens in warm and cold-blooded animals. 2 They have been associated with a variety of local and systemic infections in humans, even in immunologically competent hosts. 5 Up to date, there are 30 accepted species of Aeromonas that have been known in the world. Among them, A. hydrophila, A. veronii, and A. punctata are the most common in diseased fish. 6 They are the causative agents of many diseases, including motile aeromonad septicemia, motile aeromonad infection, hemorrhagic septicemia, red pest, and red sore in freshwater ornamental fish. 3
In ornamental fish industry, antimicrobials have been used by the fish producers as a preventive treatment of bacterial diseases, while fish have been harvested and handled. They have also been used by the retailers or owners to control bacterial infections in ornamental fish. 7 However, the excessive use of antimicrobials increases clonal selection, which facilitates the spread of antimicrobial-resistant bacteria. 8 The emergence of antimicrobial resistance among the aquatic-borne Aeromonas spp. gained lots of attention in the recent years (Table 1).
Mobile elements such as integrons, plasmids, and transposons are responsible for a wider distribution of antimicrobial resistance determinants by horizontal gene transfer among the bacteria. 19 Of these, integrons have become very important, as they can capture several antimicrobial-resistant gene cassettes, which contribute to a multiple antimicrobial resistance. 20 The amino acid sequences of integrases (IntI) divide integrons into distinct classes. The intI1, intI2, and intI3 are designated as class 1, class 2, and class 3 integrons, respectively, which are associated with mobile genetic elements. 21 The class 1 integron is the most common type of integron class in gram-negative bacteria and often found in Aeromonas spp. 22
The risk factors especially related to the transferability of ornamental fish-borne antimicrobial-resistant bacteria should be determined. However, there were no studies reported about the antimicrobial resistance gene profiling and characterization of integron gene cassettes in Aeromonas spp. isolated from goldfish and other ornamental fish in Korea. Therefore, the aim of the study was focused on the determination of antimicrobial resistance patterns, antimicrobial resistance genes, and class 1 integron gene cassettes of Aeromonas spp. isolated from goldfish.
Materials and Methods
Isolation and identification of bacteria
A total of 65 Aeromonas spp. used in this study were isolated from goldfish, which were purchased from pet shops in Seoul, Korea.
The whole body of fish was homogenized with 5 ml of sterile physiological saline. One milliliter of each homogenized sample was enriched in 9 ml of alkaline peptone water at 35°C for 24 hr. Enriched samples were streaked onto a plate of Aeromonas agar base (MB Cell, Los Angeles, CA) and incubated at 35°C for 24 hr. The colonies presumed as Aeromonas were subcultured on tryptic soy agar (MB Cell, Los Angeles, CA) overnight at 35°C and subjected to several biochemical tests such as oxidase, starch hydrolysis, indole, and H2S tests to identify the genus Aeromonas.
For species identification, PCR and sequencing of the gyrB gene were done according to the previous report. 23 The gyrB sequence data were analyzed by using BLAST option in GenBank (http://blast.ncbi.nlm.nih.gov).
Antimicrobial susceptibility testing
The antimicrobial susceptibility pattern of each isolate was determined against 18 antimicrobials by disk diffusion test using commercially available antimicrobial disks (Oxoid, United Kingdom).
The antimicrobials included ampicillin (10 μg), amoxicillin (30 μg), cephalothin (30 μg), cefoxitin (30 μg), cefotaxime (30 μg), imipenem (10 μg), nalidixic acid (30 μg), ciprofloxacin (5 μg), levofloxacin (10 μg), ofloxacin (5 μg), tetracycline (30 μg), doxycycline (30 μg), trimethoprim/sulfamethoxazole (1.25/23.75 μg), rifampicin (5 μg), gentamicin (10 μg), amikacin (30 μg), kanamycin (30 μg), and chloramphenicol (30 μg). Escherichia coli ATCC 25922 strain was used as the quality control. 7 Antimicrobial susceptibility testing and interpretation of results were done according to the recommendations of Clinical and Laboratory Standards Institute. 24
PCR amplification of antimicrobial resistance genes
For the detection of antimicrobial resistance genes, PCR amplification of extended spectrum β-lactamase (ESBL) (blaTEM, blaSHV, and blaCTX), tetracycline resistance (tetA, tetB, and tetE), aminoglycoside resistance [armA, aphAI-IAB, aac(3)-IIa, and aac(6′)-Ib)], and plasmid-mediated quinolone resistance (PMQR) (qnrA, qnrB, and qnrS) genes were performed.
The PCR primers and conditions are summarized in Table 2. PCR products were detected by electrophoresis in 2% (w/v) agarose gels. The PMQR gene, aac(6′)-Ib-cr, was identified by sequencing and BLAST comparison with GenBank.
ESBL, extended spectrum b-lactamase; PMQR, plasmid-mediated quinolone resistance.
Detection of integrase genes and class 1 gene cassettes
The PCR was performed for the detection of integrase 1 (IntI1) and integrase 2 (IntI2) genes by the specific primers described in Table 2.
The gene cassettes of IntI1-positive strains were detected by the amplification of 5′ cassette segment (5′-CS) and 3′ cassette segment (3′-CS) primers. The PCR products were sequenced and analyzed by means of BLAST option in GenBank.
Results
Identification of Aeromonas spp.
According to the gyrB gene sequencing, The Aeromonas isolates were identified as three species, including A. hydrophila (n = 30), A. veronii (n = 32), and A. punctata (n = 3).
Antimicrobial susceptibility patterns
The antimicrobial susceptibility patterns of Aeromonas spp. are shown in Fig. 1. All the isolates were resistant to at least four antimicrobials belonging to ≥3 structural classes.
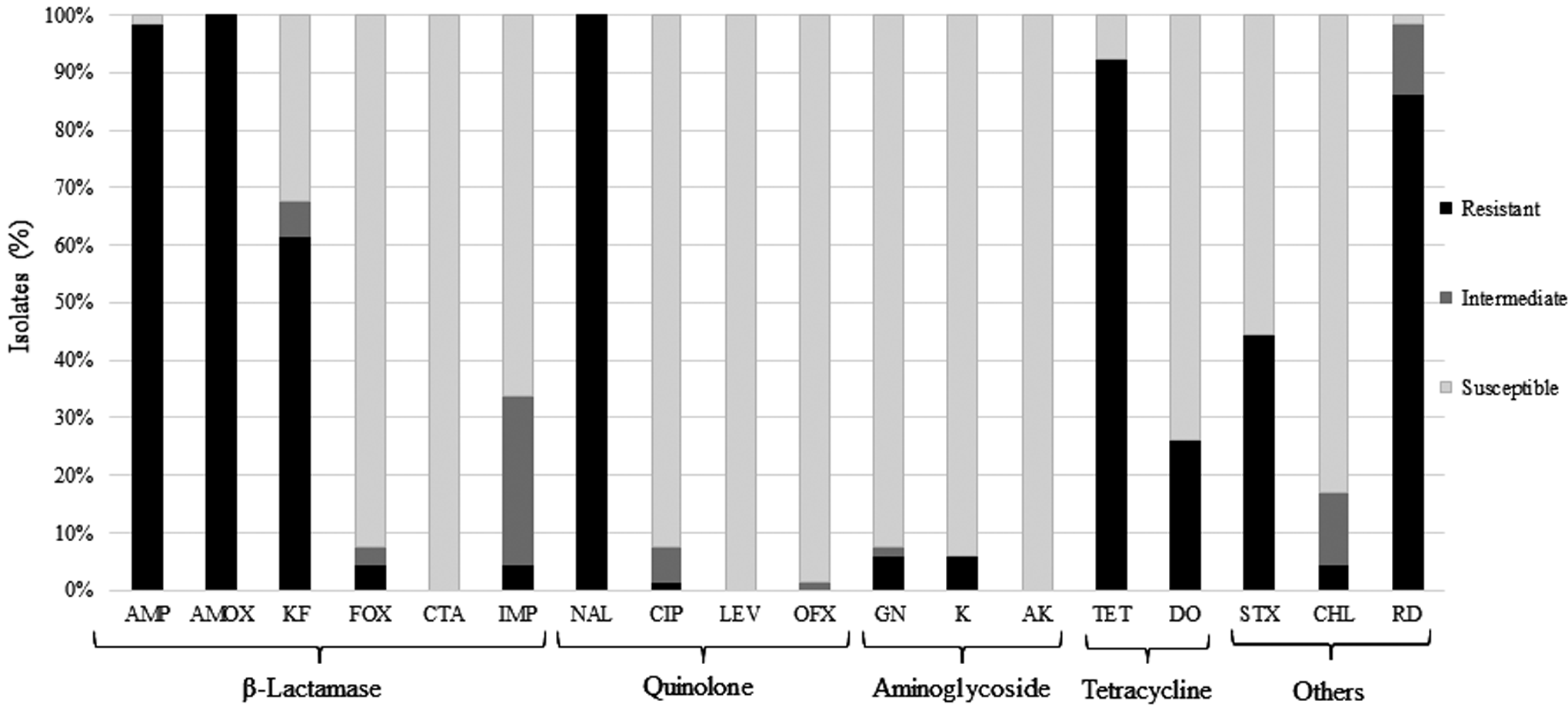
Antimicrobial susceptibility patterns of 65 Aeromonas spp. isolated from goldfish. Antimicrobials, including ampicillin, amoxicillin, cephalothin, cefoxitin, ceftriaxone, imipenem, nalidixic acid, ciprofloxacin, levofloxacin, ofloxacin, gentamicin, kanamycin, amikacin, tetracycline, doxycycline, trimethoprim/sulfamethoxazole, chloramphenicol, and rifampicin are designated as AMP, AMOX, KF, FOX, CTA, IMP, NAL, CIP, LEV, OFX, GN, K, AK, TET, DO, STX, CHL, and RD, respectively.
The resistance was prevalent for amoxicillin (100.00%), nalidixic acid (100.00%), ampicillin (98.46%), tetracycline (92.31%), rifampicin (86.15%), and cephalothin (61.54%). The resistance rates to trimethoprim/sulfamethoxazole, doxycycline, gentamicin, imipenem, cefoxitin, kanamycin, chloramphenicol, and ciprofloxacin were 44.62%, 26.15%, 6.15%, 4.61%, 4.61%, 6.15%, 4.61%, and 1.54%, respectively. The isolates showed intermediate resistance to cephalothin (6.15%), cefoxitin (3.08%), imipenem (29.23%), chloramphenicol (12.31%), rifampicin (12.31%), ciprofloxacin (6.15%), ofloxacin (1.54%), and gentamicin (1.54%). All isolates were susceptible to ceftriaxone, amikacin, and levofloxacin.
Detection of antimicrobial resistance genes
The PMQR genes, qnrS, qnrB, and aac(6′)-Ib-cr, were detected in 48 (73.85%), 17 (26.15%), and 3 (4.60%) isolates, respectively (Table 3). The tetracycline resistance gene, tetA and tetE were present in 11 (16.92%) and 14 (21.54%) isolates. The aminoglycoside resistance genes, aac(6′)-Ib and aphAI-IAB, were found in 19 (29.23%) and 5 (7.69%) isolates, respectively.
Aeromonas hydrophila, Aeromonas veronii, and Aeromonas punctata are designated as AH, AV, and AP, respectively.
Antimicrobials; Ampicillin (10 μg), amoxicillin (30 μg), cephalothin (30 μg), cefoxitin (30 μg), cefotaxime (30 μg), imipenem (10 μg), nalidixic acid (30 μg), ciprofloxacin (5 μg), levofloxacin (10 μg), ofloxacin (5 μg), tetracycline (30 μg), doxycycline (30 μg), trimethoprim/sulfamethoxazole (1.25/23.75 μg), rifampicin (5 μg), gentamicin (10 μg), amikacin (30 μg), kanamycin (30 μg), and chloramphenicol (30 μg) are marked as AMP, AMOX, KF, FOX, CTA, IMP, NAL, CIP, LEV, OFX, TET, DO, STX, RD, GN, AK, KN, and CHL, respectively.
I, intermediate resistance.
The ESBL (blaTEM, blaSHV, and blaCTX-M), aminoglycoside resistance [armA and aac(3′)-IIa], tetracycline resistance (tetB), and PMQR (qnrA) genes were not detected in any isolates.
Identification of integrase genes and gene cassettes
From 65 Aeromonas spp., 42 (64.62%) isolates were positive for intI1 gene. Among intl1 positive isolates, 15 (35.71%) carried class 1 integron gene cassettes. Four integron cassette profiles were identified according to the size of the PCR amplicons and designated as IP (IP-1 to IP-4) (Table 4).
Three resistance gene cassette arrays were identified, including catB3-aadA1 (1,200 bp), dfrA1-orfC (600 bp), and qacE2-orfD (1,015 bp) by gene sequencing (Fig. 2). The gene cassette arrays comprising catB-aadA1, dfrA1-orfC, and qacE2-orfD were detected in 3, 3, and 1 isolates, respectively. Eight strains amplified a 250 bp PCR amplicon confirmed to be the partial dfrA1 gene by sequencing (Table 4). The intI2 gene was not detected in any of the tested strains.

Genetic structure of class 1 integrons. The attI and attC sites of gene cassettes are designated as open and black circles, respectively. Three gene cassettes arrays are shown below the variable region (patterned box). Intl1, integrase gene; qacEΔ1, qac encoding marginal resistance to quaternary ammonium compounds; sul1, sulfonamide resistance gene; orf, open reading frame.
Discussion
In aquaculture, disease diagnosis is presumptive and antimicrobials are generally administered in the absence of trusted antimicrobial resistance data. The abusive use of unregistered antimicrobials is common because only a few antimicrobial agents have been approved for the disease control in Korean aquaculture. 28 Therefore, the study about the emergence of antimicrobial resistance and resistance genes in goldfish-borne Aeromonas spp. owns significance.
In this study, most of the Aeromonas strains showed multiple antimicrobial resistance phenotypes. The isolates showed 100% resistance against amoxicillin and nalidixic acid. This result is in line with a previous report where every fish-borne Aeromonas spp. also showed resistance against amoxicillin and nalidixic acid. 13 The isolates were highly resistant to tested β-lactams in this study. The Aeromonas spp. are naturally resistant to β-lactams because of the expression of chromosomal β-lactamases. 29 The high resistance rates were also observed against tetracycline, trimethoprim/sulfamethoxazole, and doxycycline. This is likely to be due to the extensive use of these antimicrobials in ornamental fish industry.7,30 Some isolates showed reduced susceptibility to chloramphenicol, ciprofloxacin, and ofloxacin although they have been banned for the treatment of aquatic animal diseases in Korea. 28
The abundance of PMQR genes was observed among the tested strains. The qnrS gene (73.85%) was the most prevalent among the isolates. The high presence of qnrS gene was found in Aeromonas spp. isolated from ornamental fish in United Kingdom. 31 In addition, the occurrence of qnrS gene was widely seen in Aeromonas spp. of different aquatic species and aquatic environments, which means the homogenous distribution of this gene among the Aeromonas spp. in different sources and geographical locations.17,32
The aminoglycoside resistance gene, aphaAI-IAB, was detected only in aminoglycoside-resistant isolates, whereas most of the aminoglycoside susceptible isolates contained aac(6′)-Ib gene. However, the isolates harboring aac(6′)-Ib gene exhibited multiple antimicrobial resistance phenotypes. The aminoglycoside resistance genes are generally located in the large plasmids, which also harbor the antimicrobial resistance genes of different antimicrobial groups.33,34
The existence of tetA and tetE genes was observed among the tetracycline resistance strains. According to the previous studies, tetA and tetE genes are the most common tetracycline resistance determinants, which are often found in Aeromonas spp.35,36 The prevalence of tetA and tetE genes was also reported in several ornamental fish-borne Aeromonas isolates. 31 However, the tetB gene could not be identified in any isolates. The absence of tetB gene was previously reported in goldfish-borne aeromonads.37,38 The presence of tetB gene was occasionally observed in Aeromonas spp. isolated from cold-water ornamental fish, whereas the higher presence of tetB gene could be detected in Aeromonas spp. isolated from tropical ornamental fish.7,31
The mobile element related to class 1 integron (intI1) gene was present in 64.62% of the tested isolates. This prevalence of class 1 integron-related intI1 gene was higher than the prevalence reported by another study, where 19.6% of IntI1-positive isolates contained gene cassettes in Aeromonas spp. isolated from different aquatic animals in China. 21 Only 35.71% of IntI1-positive isolates carried gene cassettes, which indicate the lack of gene cassettes in most of the isolates. The lack of gene cassettes might result from one of the three reasons, which are the complete absence of resistance genes in gene cassettes, nonexistence of 3′ conserved region, and higher amplicon size to amplify in conventional PCR method.39,40
The catB3 gene and aadA1 genes were observed in the integron 1 gene cassettes. The catB3 and aadA1 are responsible for chloramphenicol and streptomycin resistance. The isolates carrying catB3 gene showed susceptibility to chloramphenicol in this case. This could be due to the low level of acetyl coenzyme, which led the bacteria to be susceptible against chloramphenicol. 41 The aadA1 gene positive isolates showed susceptibility to the tested aminoglycosides, although streptomycin was not used in this study. Roe et al. 42 observed that the isolates harboring aadA1 gene showed resistance to other aminoglycosides rather than streptomycin. The trimethoprim resistance encoded dfrA1 gene was found in combination with an unknown functional open reading frame, orfC. The dfrA1-orfC array was also identified in Vibrio cholerae isolated from human clinical cases in India and Salmonella enterica from both humans and animals in Australia.43–45 The incidence of same gene cassette array in different species indicates the transferability of gene cassettes in intra- and interspecies level, which confirms the dissemination of antimicrobial resistance genes among the bacteria through horizontal gene transfer. 41
A partial dfrA1 gene was the most common gene cassette found among the intI1-positive isolates. It was suggested that the partial or incomplete gene has been picked up by the disturbance of recombination event during the integron transferring process, which provides no advantages for the bacteria. 46
The qacE2-orfD gene cassette was detected only in one isolate. The qacE2 gene relates to a larger efflux of quaternary ammonium compounds, and the orfD gene has no known function. The qacE2 gene is associated with multiple antimicrobial resistance and can be horizontally transferred via plasmids among the bacteria and transmitted together with other antimicrobial-resistant genes. 47
The studies suggest that goldfish is a reservoir for Aeromonas spp. comprising multiple antimicrobial resistance phenotypes, diverse antimicrobial resistance genes, and class 1 integrons. The association of multiple antimicrobial resistance with integrons expands the risk of coselection and persistence of other resistance genes under the selective pressure executed by the application of antimicrobials. Thus, the epidemiology and evolution of plasmids related to the antimicrobial resistance genes of goldfish-borne Aeromonas spp. should be further studied. Considering the rapid growth of the ornamental fish industry, the antimicrobial use for prophylactic purposes must be substituted by better husbandry and transportation.
Footnotes
Disclosure Statement
No competing financial interests exist.