
Abstract
The aim of the present study was to describe the first mphC-positive staphylococci, including two Staphylococcus lentus (Sle-087lar and Sle-091lar) and one Staphylococcus xylosus (Sxy-228lar), isolated from samples of animal origin, in Greece. Isolates Sle-087lar and Sxy-228lar were resistant to erythromycin, whereas Sle-091lar was resistant to erythromycin and lincomycin. All three isolates were susceptible to the remaining antibiotics. PCR screening showed that isolate Sle-091lar carried also ermB. For Sxy-228lar, whole-genome sequencing (WGS) and de novo assembly obtained an mphC-positive contig of 57.3-kb exhibiting high similarity with the genome of mphC-negative S. xylosus S170. However, mphC of Sxy-228lar was 91% similar to that found in plasmid pJW2311 from S. xylosus JW2311. Additionally, WGS data showed that Sle-087lar and Sle-091lar harbored mphC-carrying sequences being highly similar to the recently announced genome of the mphC-carrying S. lentus isolate 050AP from Tanzania. However, differences were observed in the mphC environment, suggesting the independent acquisition of the gene by each isolate. Sle-091lar also harbored transposon Tn917, which carries ermB resistance gene, integrated into S. lentus chromosome. These findings indicated that acquisition of resistance genes can lead to the emergence of multiresistant staphylococci, causing animal infections with economic burden.
Introduction
M
Therefore, the aim of the present study was to describe the characterization of the first mphC-positive staphylococci isolated from samples of animal origin, in Greece.
Materials and Methods
Bacterial strains
Milk samples were collected in 30-ml plastic tubes with preservative (bronopol), using specific sampling devices during milking, or by automatic sampling devices on milking robots from clinically healthy ewes from farms located throughout Greece. One hundred twenty-one staphylococci were recovered from ewes, which were diagnosed with subclinical mastitis, based on simultaneous bacterial isolation and increased somatic cell counts (>1.0 × 106 cells/ml). 7 The isolates were identified to species level based on catalase and coagulase production (SLIDEX Staph Plus; bioMerieux, Marcy l'Etoile, France) and by the automated system Vitek II (bioMerieux)
Susceptibility testing
All isolates were tested for susceptibility to various antimicrobial agents (azithromycin, benzylpenicillin, cefoxitin, clindamycin, daptomycin, erythromycin, fosfomycin, fusidic acid, gentamicin, imipenem, levofloxacin, linezolid, moxifloxacin, mupirocin, nitrofurantoin, oxacillin, rifampicin, teicoplanin, tetracycline, tigecycline, tobramycin, trimethoprim/sulfamethoxazole, and vancomycin) by Vitek II and E-test (bioMerieux). Minimum inhibitory concentrations (MICs) were interpreted according to European Committee on Antimicrobial Susceptibility Testing (EUCAST) criteria a .
Detection of resistance determinants
Genes encoding for resistance to macrolides were detected by PCR, using specific primers for ermA, ermB, ermC, ermY, ermF, msrA, msrB, ereA, ereB, lnuA, lnuB, and mphC. 8
Illumina sequencing
Genomic DNAs from the wild isolates were extracted using the DNASorb-B Kit (Sacace Biotechnologies Srl, Como, Italy). Multiplexed DNA libraries were prepared, using the Nextera XT Library Preparation Kit, and 300-bp paired-end sequencing was performed on the Illumina MiSeq platform (Illumina, Inc., San Diego, CA) using the MiSeq v3 600-cycle Reagent Kit. Initial paired-end reads were quality trimmed, using the Trimmomatic tool v0.33, 9 with the sliding window size of 4 bp, required average base quality ≥17, and minimum read length of 48 bases. Then, reads were assembled by use of the de Bruijn graph-based de novo assembler SPAdes v3.9.1, 10 using using k-mer sizes 21, 33, 55, and 77. For sequence analysis and annotation, the BLAST algorithm (www.ncbi.nlm.nih.gov/BLAST), the ISfinder database (www-is.biotoul.fr/), and the open reading frame finder tool (www.bioinformatics.org/sms) were utilized. Comparative genome alignments were performed using the Mauve (version 2.3.1) program. 11
Antibiotic resistance genes were identified using the ResFinder 2.1 tool b with an identity threshold of >90%. 12
Phylogenetic analysis
Comparative genomic analysis of isolates was done based on metrics calculated by QUAST tool (Version 4.5). 13
Nucleotide sequence accession numbers
The nucleotide sequences of mphC-carrying regions from Sxy-228lar, Sle-087lar, and Sle-091lar have been assigned GenBank accession numbers MG557992, MG557990, and MG557991, respectively. The ermB-carrying sequence from Sle-091lar has been assigned GenBank accession number MG557993.
Results
Among 121 staphylococcal isolates, 31 were identified as S. aureus, whereas the remaining 90 were classified to various species of CoNS, with predominance of Staphylococcus chromogenes, Staphylococcus epidermidis, Staphylococcus lentus, and Staphylococcus xylosus. Seven macrolide-resistant staphylococci were detected; three of them, two S. lentus (Sle-087lar and Sle-091lar) and one S. xylosus (Sxy-228lar), all positive for the presence of mphC, were further analyzed. MICs showed that isolates Sle-087lar and Sxy-228lar were resistant to erythromycin (MIC: 8 μg/ml), whereas Sle-091lar was resistant to erythromycin (MIC: 256 μg/ml) and clindamycin (MIC: 64 μg/ml). All three isolates were susceptible to the remaining antibiotics. PCR screening showed that isolate Sle-091lar carried also ermB.
Analysis of Illumina data for the presence of 16S rRNA 14 and tuf sequences 15 confirmed the identification of the isolates as S. lentus and S. xylosus. Analysis of whole-genome sequencing (WGS) data by the ResFinder v2.1 tool 12 revealed that none of the sequenced isolates included additional resistance genes.
Additionally, Illumina sequencing and de novo assembly obtained a unique contig of 57.3-kb carrying mphC for S. xylosus Sxy-228lar. Sequence analysis showed that the 57.3-kb fragment exhibited high similarity (92% coverage, 95% identity) with the genomes of S. xylosus isolates HKUOPL816 and S170 (GenBank accession no. CP013922) (Fig. 1). However, isolates HKUOPL8 and S170 were not found to harbor mphC. The mphC gene of Sxy-228lar was 91% similar to that found in plasmid pJW2311 from S. xylosus JW2311 (GenBank accession no. LT223129). Of note, no mobile elements that could be implicated in the integration of mphC gene in the chromosome of S. xylosus were found in its surroundings.
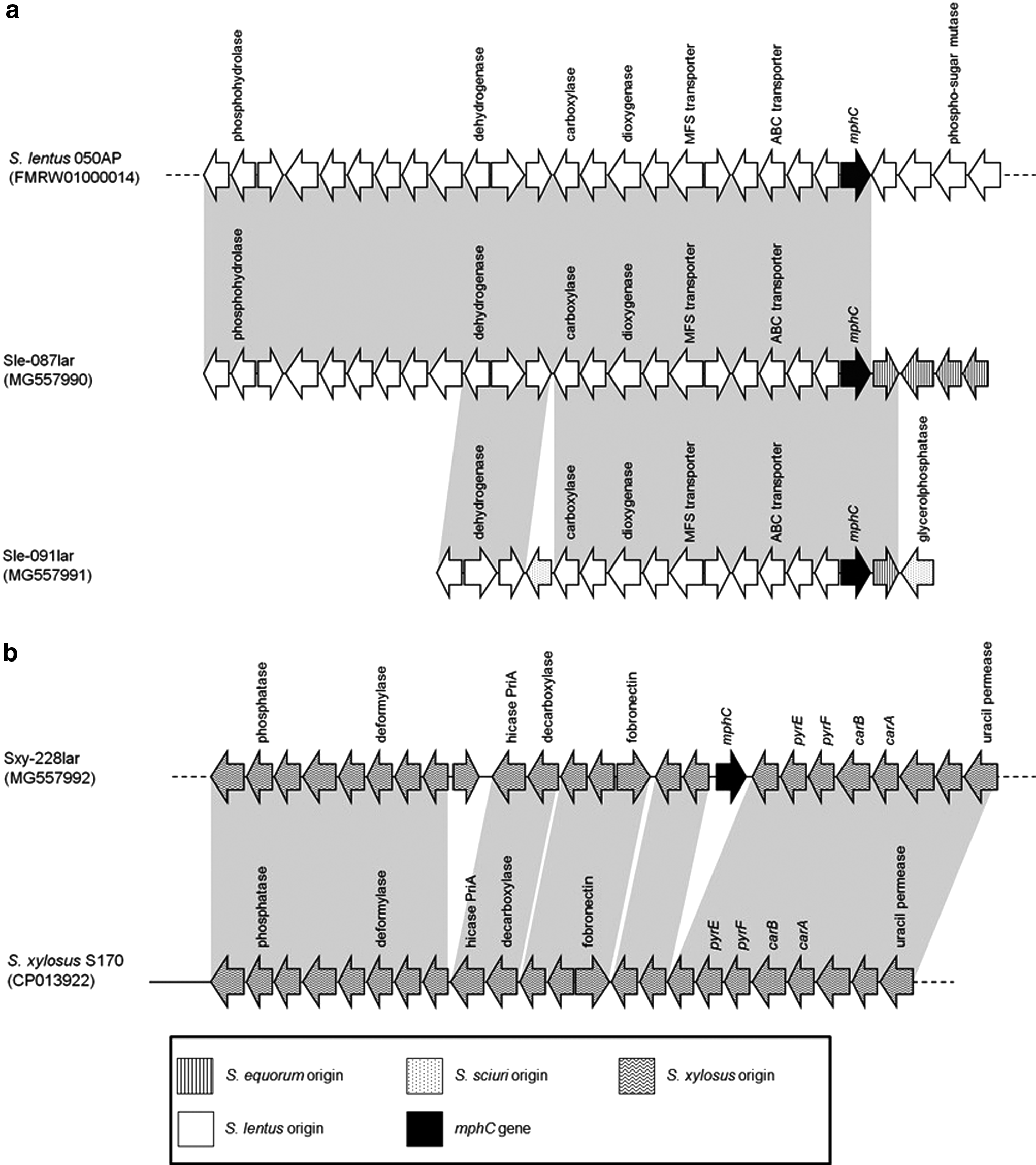
Detailed comparison of the mphC-carrying regions in Staphylococcus lentus isolates Sle-087lar and Sle-091lar
In WGS data of S. lentus Sle-087lar, mphC gene was identified on a unique contig of 26.5-kb, which differed from the observed region in S. xylosus Sxy-228lar. The mphC-carrying sequence of Sle-087lar was highly similar (84% coverage, 99% identity) to the recently announced genome of the mphC-carrying S. lentus isolate 050AP from Tanzania 17 (Fig. 1). However, downstream of mphC, Sle-087lar included a 5.3-kb segment (nt 21172–26498), which was absent from the genome of S. lentus 050AP. Part of this segment, which comprised four coding sequences for proteins with unknown function, has been previously found in the chromosome of Staphylococcus equorum isolate KS1039 from South Korea (GenBank accession no. CP013114).
Sequencing data showed that isolate Sle-091lar harbored an mphC-carrying fragment of 15.2 kb, exhibiting extensive similarity (87% coverage, 99% identity) to the respective sequence of Sle-087lar (Fig. 1). Only two differences between the two mphC-carrying regions were observed. A 0.75-kb sequence (nt 3015–3754), which was previously found in the mphC-positive Staphylococcus sciuri isolates FDAARGOS_285 (GenBank accession no. CP022046) and SNUDS-18, 18 was identified in Sle-091lar. This sequence that encoded a transcriptional regulatory protein was not present in the WGS data of S. lentus isolates Sle-087lar and 050AP. 17 In addition, a 1.4-kb sequence of S. sciuri origin was identified, downstream of mphC gene, in Sle-091lar. Similar sequences (74%) were found in non-mphC-carrying contigs of Sle-087lar and 050AP. Differences in the genetic environment of mphC suggested the independent acquisition of the gene by the different isolates.
Furthermore, Sle-091lar included ermB resistance gene on a unique contig of 62.9-kb. Sequence analysis showed that ermB was part of the transposon Tn917, which was originally described from Streptococcus faecalis DS16, 19 integrated into S. lentus chromosome. Target site duplications of 6 bp (ACATTA) at the boundaries of Tn917 indicated insertion by transposition.
To examine the phylogenetic relationships of S. lentus isolates, Sle-087lar, Sle-091lar, and 050AP, 17 metrics were calculated by QUAST tool (Version 4.5) 13 using WGS data of isolate Sle-087lar as a reference. Comparative analysis showed that genome fractions (i.e., percentage of aligned bases in the reference genome) of isolates Sel-091lar and 050AP were 92% and 90%, respectively. On average, genomic content of Sle-091lar had 254 of mismatches and 13 indels per 100,000 aligned bases compared with reference genome, whereas genomic content of 050AP had 486 mismatches and 22 indels per 100,000 bp. These findings clearly indicated that the studied S. lentus isolates were not related to each other.
Discussion
To our knowledge, this is the first description of mphC-positive S. lentus and S. xylosus isolates from samples of animal origin, from Greece. S. lentus and S. xylosus are staphylococcal species, which are frequently isolated from animals, and cause infections with economic burden. Sequencing data showed that the mphC gene was integrated into the chromosomes of S. xylosus and nonrelated S. lentus isolates. However, the mphC of Sxy-228lar was 89% similar to the respective genes of Sle-087lar and Sle-091lar isolates, suggesting a different origin of the resistance genes. Additionally, differences observed in the environment of mphC genes suggested independent acquisition events. This hypothesis was further supported by the findings of phylogenetic analysis showing that S. lentus isolates were not related to each other. Interestingly, no mobile elements were observed in the surroundings of mphC genes, supposing that homologous recombination could be the mechanism for the acquisition of mphC by S. xylosus and S. lentus. Furthermore, isolate Sle-091lar harbored transposon Tn917, which carries ermB resistance gene.
Conclusion
These findings indicated that acquisition of resistance genes by different staphylococcal species, even by the spread of diverse genetic structure, can lead to the emergence of multiresistant staphylococci, causing animal infections with economic burden.
Footnotes
Acknowledgments
This work was supported by funding from the Research Committee of the University of Thessaly. It was also financed partially by the Medical Research Foundation of the Czech Republic (grant number 17-29239A).
Disclosure Statement
No competing financial interests exist.