
Abstract
This study was designed to evaluate the resistance phenotype and genotype of wild type (WT)-, cefotaxime (CET)-, and ciprofloxacin (CIP)-induced Salmonella Typhimurium ATCC 19585, CIP-resistant Salmonella Typhimurium ATCC 19585, Salmonella Typhimurium CCARM 8009, and Salmonella Typhimurium KCCM 40253 before and after exposure to pH 4.5, 4% NaCl, and heat at 42°C. The susceptibilities of WT Salmonella Typhimurium ATCC 19585 and WT Salmonella Typhimurium KCCM 40253 to all antibiotics tested in this study were decreased after CET and CIP induction with the exception with kanamycin, meropenem, and polymyxin B. The highest β-lactamase activities were 2.8 and 3.3 nmol/(min·mL), respectively, at the WT- and CET-induced Salmonella Typhimurium CCARM 8009. FT-IR spectra were found to be dominant at the region from 1,700 to 1,500 cm−1 corresponding to proteins such as amides I, II, and III. The relative expression levels of efflux pump-related genes (acrA, acrB, and TolC), porin-related gene (ompC), virulence-related gene (stn), adhesion-related gene (fimA), and stress-induced alternative sigma factor (rpoS) varied in the antibiotic resistance and stress exposure. This study provides useful information for understanding the antibiotic resistance profile, physicochemical property, and gene expression pattern in Salmonella Typhimurium in association with the induction of antibiotic resistance and exposure to environmental stresses.
Introduction
O
The effective treatment of MDR Salmonella infections remains a high priority in clinical area.7,9 Fluoroquinolones and cephalosporins are commonly used to treat Salmonella infections. 9 Recently, combination therapy has been developed to enhance the antibiotic efficacy against MDR bacteria. 10 Although the combination therapy is beneficial in the treatment of mixed bacterial infections, there are still questions with regard to the cross-resistance in MDR bacteria. 11 Bacteria can encounter different conditions, including physical (heat, pressure, and osmotic shock), chemical (acids, salts, and oxidants), and biological (bacterial competition and secondary metabolites) stresses, leading to the induction of cross-protective responses. However, relatively few studies have investigated the patterns of phenotypic and genotypic variations in MDR bacteria exposed to environmental stresses. Therefore, the objective of this study was to evaluate the antibiotic resistance profiles, physicochemical properties, and gene expression in cefotaxime (CET)- and ciprofloxacin (CIP)-induced Salmonella Typhimurium when exposed to acidic, heat, and osmotic stresses.
Materials and Methods
Bacterial strains and culture conditions
Strains of Salmonella Typhimurium ATCC 19585, Salmonella Typhimurium CCARM 8009, and Salmonella Typhimurium KCCM 40253 were purchased from American Type Culture Collection (ATCC, Manassas, VA), Culture Collection of Antibiotic Resistant Microbes (CCARM, Seoul, Korea), and Korean Culture Center of Microorganism (KCCM, Seoul, Korea), respectively. The CIP-resistant Salmonella Typhimurium ATCC 19585 was also used in this study. All strains were cultivated in trypticase soy broth (TSB) (BD; Becton, Dickinson and Co., Sparks, MD) at 37°C for 20 hr, centrifuged at 5,000 g for 20 min at 4°C, and diluted with phosphate-buffered saline (PBS; pH 7.2) to 108 colony-forming unit (CFU)/mL before use.
Induction of antibiotic-resistant strains
Salmonella Typhimurium ATCC 19585, CIP-resistant Salmonella Typhimurium ATCC 19585, Salmonella Typhimurium CCARM 8009, and Salmonella Typhimurium KCCM 40253 were exposed to CET and CIP to induce antibiotic-resistant strains according to a stepwise selection assay. 12 In brief, all strains were successively cultured in TSB and trypticase soy agar (TSA) containing serially increased concentrations of CET and CIP from 1/2 minimal inhibitory concentrations (MICs). After several passages, the resistance stability of CET- and CIP-induced resistant Salmonella Typhimurium ATCC 19585, Salmonella Typhimurium CCARM 8009, and Salmonella Typhimurium KCCM 40253 was confirmed through 10 culture passages in antibiotic-free TSB. 13
Antimicrobial susceptibility assay
The antibiotic susceptibilities of wild type (WT)-, CET-, and CIP-induced Salmonella Typhimurium ATCC 19585, CIP-resistant Salmonella Typhimurium ATCC 19585, Salmonella Typhimurium CCARM 8009, and Salmonella Typhimurium KCCM 40253 were determined according to a broth microdilution assay with modification. 14 Antibiotic stock solutions were prepared to obtain a final concentration of 10.24 mg/mL. The stock solutions (100 μL each) were serially (1:2) diluted with TSB from 512 μg/mL in 96-well microtiter plates containing ∼106 CFU/mL of each strain. The prepared microtiter plates were incubated at 37°C for 18 hr. After incubation, the MICs were determined at the lowest antibiotic concentration at which no visible growths were observed.
β-Lactamase activity assay
The β-lactamase activity of WT-, CET-, and CIP-induced Salmonella Typhimurium ATCC 19585, CIP-resistant Salmonella Typhimurium ATCC 19585, Salmonella Typhimurium CCARM 8009, and Salmonella Typhimurium KCCM 40253 cells exposed to pH 4.5, 4% NaCl, and heat at 42°C for 30 min was measured by using a nitrocefin hydrolyzing assay kit (BioVision, Inc., Milpitas, CA) according to the instruction manual. The cell-free supernatants were incubated with 50 μL of reaction mixture containing 48 μL of β-lactamase assay buffer and 2 μL of nitrocefin at 37°C for 30 min. The absorbance was measured at 515 nm using a microplate reader (BioTek Instruments, Inc., Winooski, VT). A standard curve was prepared at 0, 5, 10, 15, 20, 30, 40, and 50 nmol of hydrolyzed nitrocefin standard. The β-lactamase activity was expressed as nmol/(min·mL). 15
FT-IR analysis
The biochemical properties of WT-, CET-, and CIP-induced Salmonella Typhimurium ATCC 19585, CIP-resistant Salmonella Typhimurium ATCC 19585, Salmonella Typhimurium CCARM 8009, and Salmonella Typhimurium KCCM 40253 were evaluated using an FT-IR spectrometer (Excalibur series; Bio-Rad, Cambridge, MA) equipped with a UMA-500 microscope. The cultured cells were lyophilized and used for FT-IR analysis in transmittance mode. FT-IR spectra were collected in wave numbers ranging from 4,000 to 500 cm−1 with a resolution of 4 cm−1.
Environmental stress susceptibility assay
The growth curves of WT-, CET-, and CIP-induced Salmonella Typhimurium ATCC 19585, CIP-resistant Salmonella Typhimurium ATCC 19585, Salmonella Typhimurium CCARM 8009, and Salmonella Typhimurium KCCM 40253 were determined to evaluate the bacterial viability and behavior under stress conditions. In brief, the bacterial cells (105 CFU/mL) were exposed to pHs (4.5, 5.0, 7.0, and 9.0) for 24 hr, NaCl concentrations (0%, 1%, 1.5%, 2%, 3%, 4%, and 5%) for 24 hr, and temperature at 42°C for 15 hr. The optical density of each culture was measured at a wavelength of 600 nm at different time intervals. The growth curves were analyzed using Nonlinear Curve Fitting Function of Microcal Origin® 8.5 (Microcal Software, Inc., Northampton, MA).
RNA extraction and complementary DNA synthesis
Total RNAs from WT-, CET-, and CIP-induced Salmonella Typhimurium ATCC 19585, CIP-resistant Salmonella Typhimurium ATCC 19585, Salmonella Typhimurium CCARM 8009, and Salmonella Typhimurium KCCM 40253 exposed to pH 4.5 for 24 hr at 37°C, 4% NaCl for 24 hr at 37°C, and heat at 42°C for 15 hr were extracted according to the protocol of RNeasy Protect Bacteria Mini Kit (Qiagen, Hilden, Germany). The bacterial cells (500 μL) were mixed with 1 mL of RNAprotect Bacteria Reagent and centrifuged at 5,000 g for 10 min. The harvested cells were lysed with a buffer containing lysozyme and then mixed with ethanol to extract RNA using an RNeasy mini column. To remove genomic DNA (gDNA), the extracted RNA was mixed with gDNA wipeout buffer and incubated at 42°C for 2 min. The extracted RNA was mixed with reverse transcriptase, RT buffer, and RT primer mix and incubated at 42°C for 15 min followed by 95°C for 3 min to synthesize complementary DNA (cDNA) according to the QuantiTect Reverse Transcription procedure (Qiagen).
Quantitative RT-PCR assay
The prepared cDNA was used as a template for quantitative RT-PCR assay. The reaction mixture (10 μL of 2× QuantiTect SYBR Green PCR Master, 2 μL of each primer, and 2 μL of cDNA, and 4 μL of RNase-free water) was amplified using an iCycler iQ™ System (Bio-Rad Laboratories, Hemel Hempstead, UK). The primers used in this study are listed in Table 1. The mixture was denatured at 95°C for 30 sec, followed by 45 cycles of 95°C for 5 sec, 55°C for 20 sec, and 72°C for 15 sec. The relative gene expression levels were estimated using the comparative method. 16
F, forward; R, reverse.
Statistical analysis
All experiments were carried out in duplicate for three replicates. The obtained data were analyzed by the Statistical Analysis System (SAS) software. The general linear model and Fisher's least significant difference procedures were used to determine significant mean differences among the strains tested at p < 0.05.
Results
Antibiotic susceptibility of antibiotic-induced resistant Salmonella Typhimurium
The MICs of selected antibiotics against WT-, CET-, and CIP-induced Salmonella Typhimurium ATCC 19585, CIP-resistant Salmonella Typhimurium ATCC 19585, Salmonella Typhimurium CCARM 8009, and Salmonella Typhimurium KCCM 40253 were determined as given in Table 2. The antibiotic susceptibility patterns after CET and CIP induction varied in the degree of antibiotic resistance of WT strains. The susceptibilities of WT strains to most antibiotics were decreased after the induction of antibiotic resistance. However, the CIP-induced Salmonella Typhimurium CCARM 8009 was more sensitive to ampicillin, cephalothin, kanamycin, meropenem, oxacillin, penicillin, piperacillin, streptomycin, and tetracycline than WT Salmonella Typhimurium CCARM 8009. The β-lactamase activities were measured in WT-, CET-, and CIP-induced Salmonella Typhimurium ATCC 19585, CIP-resistant Salmonella Typhimurium ATCC 19585, Salmonella Typhimurium CCARM 8009, and Salmonella Typhimurium KCCM 40253. No β-lactamase activities were observed for all strains with the exception of WT- and CET-induced Salmonella Typhimurium CCARM 8009, showing 2.8 and 3.3 nmol/(min·mL), respectively.
CET, cefotaxime; CIP, ciprofloxacin; MIC, minimal inhibitory concentration; WT, wild type.
Variability in adaptive resistance phenotype to environmental stresses
The WT-, CET-, and CIP-induced Salmonella Typhimurium ATCC 19585, CIP-resistant Salmonella Typhimurium ATCC 19585, Salmonella Typhimurium CCARM 8009, and Salmonella Typhimurium KCCM 40253 were cultured in different pHs (4.5–9.0), NaCl concentrations (0% to 5%), and temperature (42°C) to evaluate the growth and survival (Figs. 1–3). Compared with the WT strains, the decreased growths were observed at the CET- and CIP-induced resistant strains at pH 7 (Fig. 1I–L) and 0% NaCl (Fig. 2B–D), whereas no significant decrease in the growth was observed at pHs 4.5 and 5.0 (Fig. 1A–E), NaCl at 4% and 5% (Fig. 2Q–X), and temperature at 42°C (Fig. 3A–D).

Growth curves of WT- (circle), CET- (diamond), and CIP (square)-induced Salmonella Typhimurium ATCC 19585
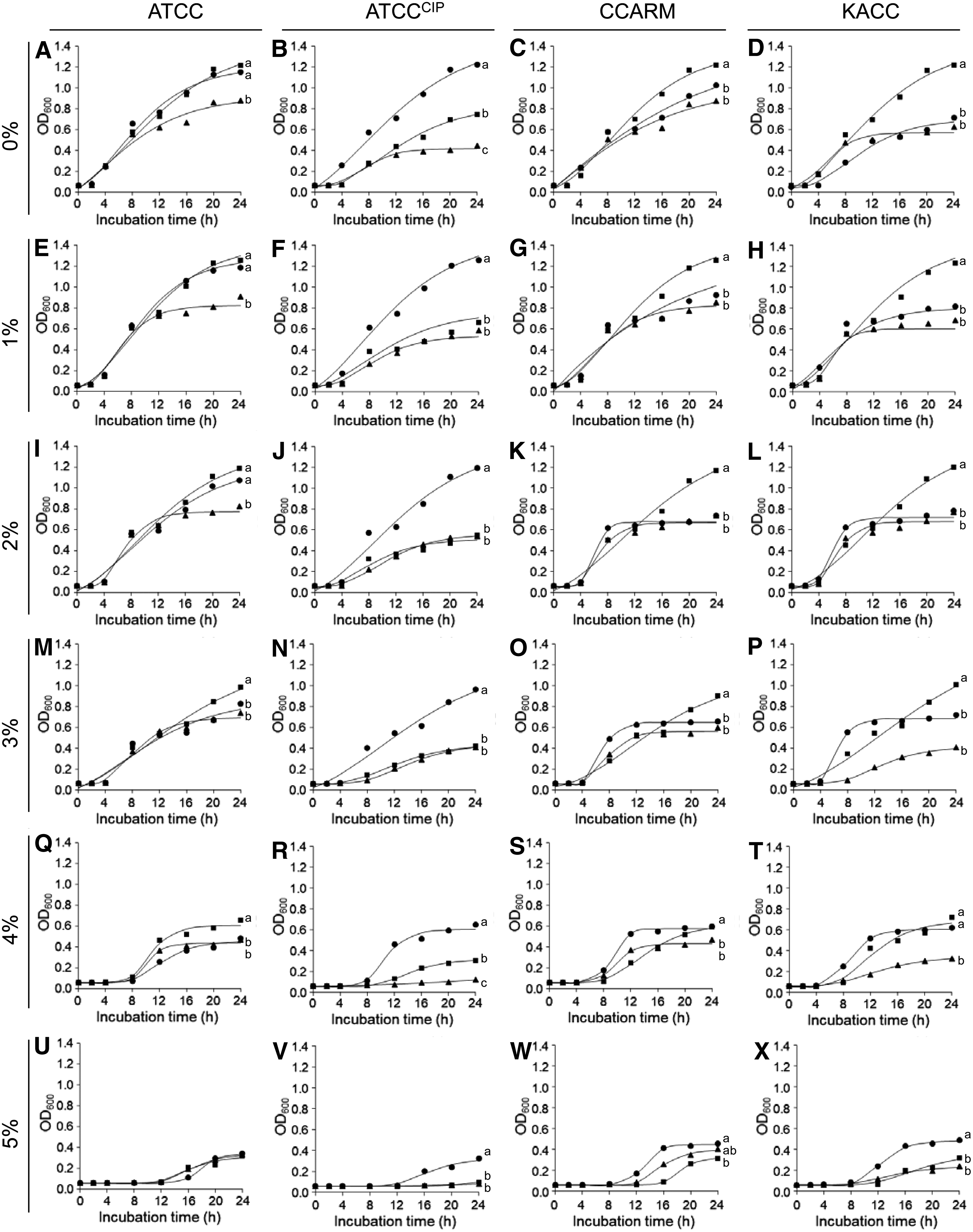
Growth curves of WT- (circle), CET- (diamond), and CIP (square)-induced Salmonella Typhimurium ATCC 19585
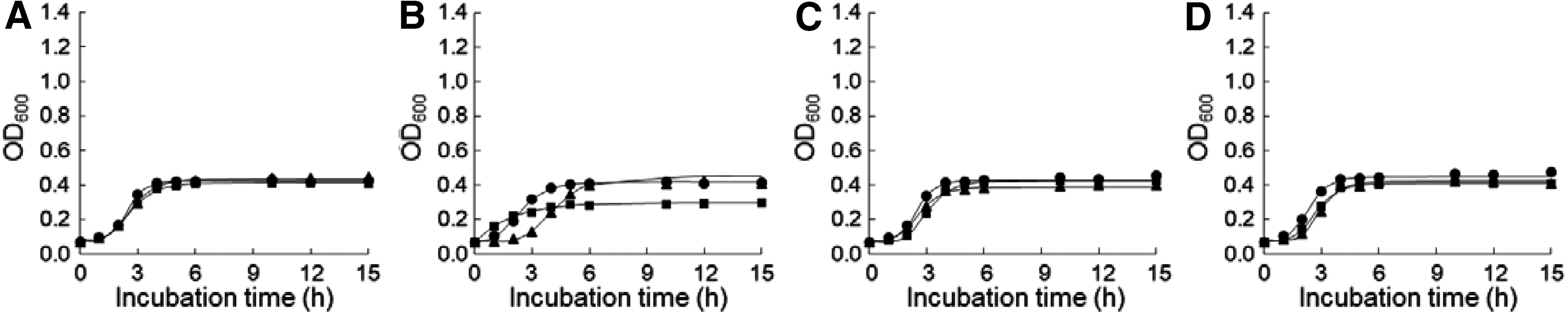
Growth curves of WT- (circle), CET- (diamond), and CIP (square)-induced Salmonella Typhimurium ATCC 19585
FT-IR spectroscopy was used to characterize the chemical components of WT-, CET-, and CIP-induced Salmonella Typhimurium ATCC 19585, CIP-resistant Salmonella Typhimurium ATCC 19585, Salmonella Typhimurium CCARM 8009, and Salmonella Typhimurium KCCM 4025 (Fig. 4). The entire spectral range of infrared region was 4,000 to 500 cm−1, which was divided into lipid region (3,000–2,800 cm−1), protein region (1,700–1,500 cm−1), phospholipid–nucleic acid region (1,500–1,200 cm−1), carbohydrate region (1,200–900 cm−1), and fingerprint region (900–700 cm−1). The highest absorbance intensities at 3,300 cm−1 were observed for CET-induced Salmonella Typhimurium ATCC 19585 and CIP-induced Salmonella Typhimurium KCCM 4025 (Fig. 4A, D). Various amide-related bands were observed for all strains, including amide I region (α-helical structures) at 1,650 cm−1, amide II region (β-pleated sheet structures) at 1,540 cm−1, and amide III region at 1,235 cm−1.
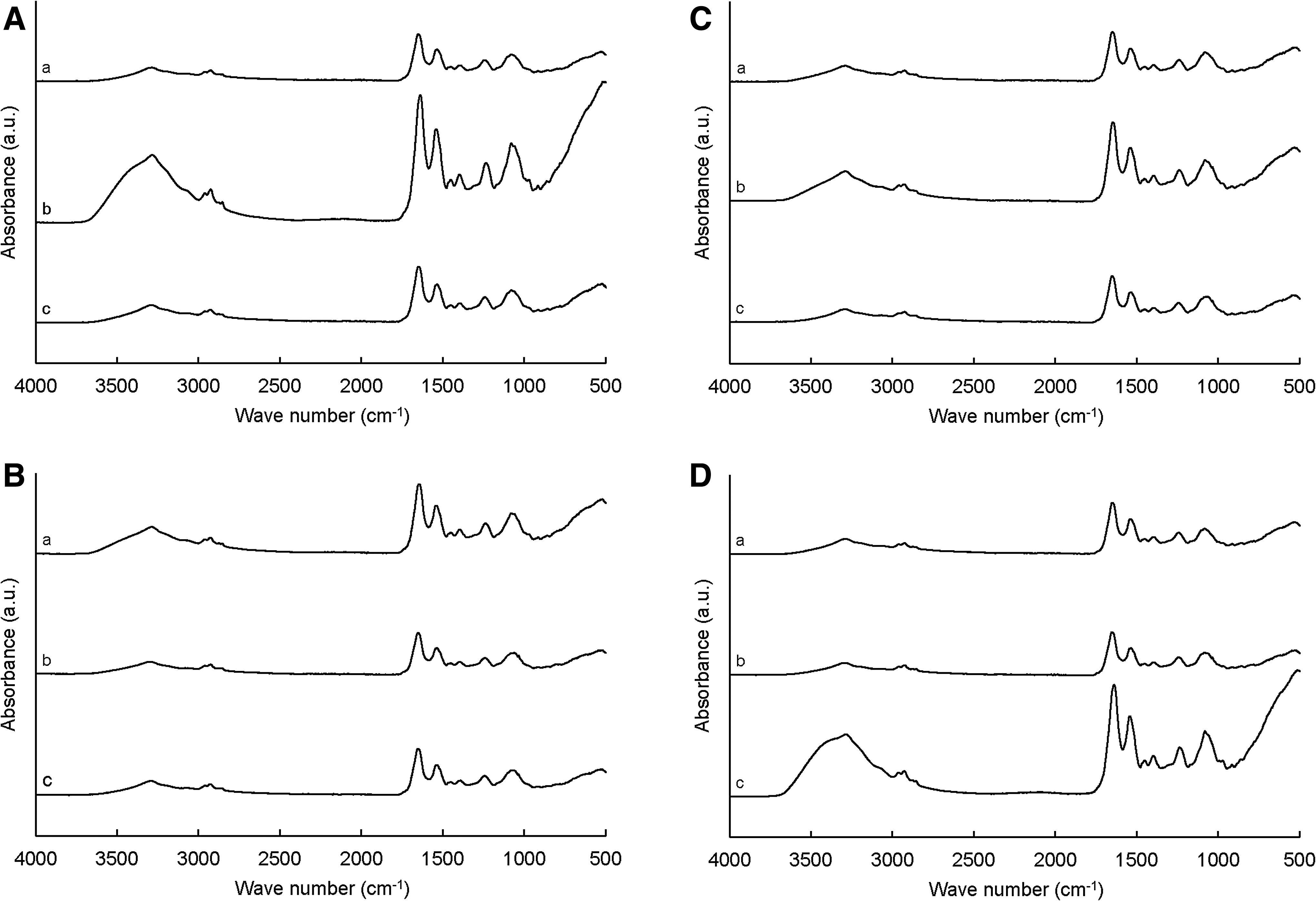
FT-IR spectra of Salmonella Typhimurium ATCC 19585
Variability in adaptive resistance genotype to environmental stresses
The relative expression levels of efflux pump-related genes (acrA, acrB, and TolC), porin-related gene (ompC), virulence-related gene (stn), adhesion-related gene (fimA), and stress-induced alternative sigma factor (rpoS) were observed in WT-, CET-, and CIP-induced Salmonella Typhimurium ATCC 19585, CIP-resistant Salmonella Typhimurium ATCC 19585, Salmonella Typhimurium CCARM 8009, and Salmonella Typhimurium KCCM 40253 exposed to pH 4.5, 4% NaCl, and heat at 42°C (Fig. 5). The transcriptional responses of all strains tested in this study varied in acidic, osmotic, and heat stresses. The relative expression levels of acrA, acrB, and tolC genes varied with the antibiotic resistance and stress exposure. The acrA, acrB, and tolC genes were upregulated in most strains tested, whereas they downregulated in CET-induced Salmonella Typhimurium CCARM 8009 (Fig. 5H). The downregulation of ompC was observed at the CET-induced Salmonella Typhimurium ATCC 19585 exposed to pH 4.5, 4% NaCl, and heat at 42°C (Fig. 5B). The highest expression of ompC was observed at CIP-induced Salmonella Typhimurium KCCM 40253 exposed to pH 4.5, 4% NaCl, and heat at 42°C (Fig. 5L), followed by CIP-induced CIP-resistant Salmonella Typhimurium ATCC 19585 (Fig. 5F) and Salmonella Typhimurium CCARM 8009 (Fig. 5I). The rpoS gene in all strains tested was overexpressed when exposed to pH 4.5, 4% NaCl, and heat at 42°C (Fig. 5). The relative gene expression level of stn was increased more than sevenfold and fivefold, respectively, in the CET-induced CIP-resistant Salmonella Typhimurium ATCC 19585 and Salmonella Typhimurium KCCM 40253 exposed to 42°C, whereas that was decreased more than sevenfold in the CET-induced Salmonella Typhimurium ATCC 19585 exposed to pH 4.5 and 4% NaCl.
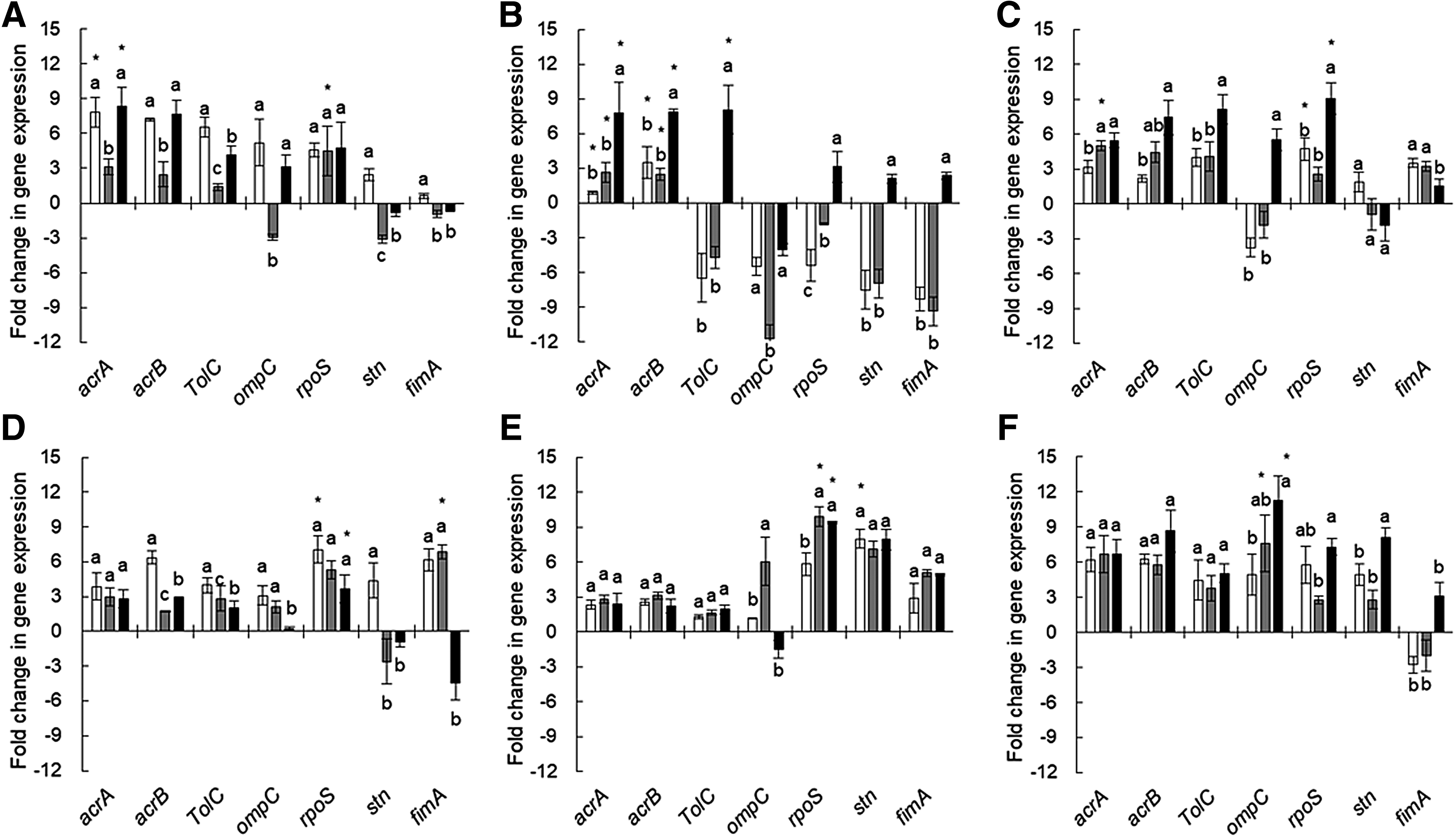
Relative gene expression in WT-
Discussion
This study describes the variability in resistance phenotype and genotype of antibiotic-resistant Salmonella Typhimurium exposed to environmental stresses, including acidic, osmotic, and heat stresses. The emergence of MDR bacteria has been of great concern because of the limitations of chemotherapy resulting in treatment failure. Frequent exposure to stresses (antibiotics, acid, heat, and NaCl) influences the acquisition and spread of multiple antibiotic resistance in nature. To the best of our knowledge, this study is the first to describe the cross-protective response of CET- and CIP-induced MDR Salmonella Typhimurium to environmental stresses. CET and CIP were used to induce antibiotic resistance, representing the third generation of cephalosporin and the second generation of fluoroquinolone, respectively.
The antibiotic resistance in all strains was increased after CET and CIP induction with the exception of kanamycin, meropenem, and streptomycin (Table 2). The increase in antibiotic resistance indicates that the serial exposure to antibiotic concentrations can induce multidrug resistance in bacteria. 17 As shown in the CIP-induced Salmonella Typhimurium CCARM 8009 (Table 2), the change in susceptibility to a broad range of nonquinolone antibiotics may be attributed to the gyr A mutation in Salmonella Typhimurium. 18 The induction rate of antibiotic resistance may depend on the inherited antibiotic resistance profiles. The development of multiple antibiotic resistance to β-lactams, fluoroquinolones, tetracycline, and macrolides is because of the antibiotic selection pressure, leading to pleiotropic resistance. 19 This is in good agreement with the previous report that the susceptibility of Gram-negative bacteria to aminoglycosides was increased after the induction of antibiotic resistance. 20 The decrease in affinity of fluoroquinolones results from the alterations in DNA gyrase and topoisomerase IV, leading to the induction of antibiotic resistance to CIP.21,22 The increased resistance of β-lactams and fluoroquinolones is associated with the efflux pump activity and outer membrane permeability. 23 The bacterial resistance to β-lactams can be mediated by not only β-lactamase production but also by other mechanisms such as low-affinity penicillin-binding proteins and membrane permeability.24,25 The induction of β-lactamases can be stimulated by the exposure to CET in this study.
Under sublethal pH, NaCl, and temperature, the CET- and CIP-induced strains showed similar or better survival than the WT strains. This result suggests that antibiotic-resistant bacteria have the ability to adapt to environmental stresses.26,27 FT-IR spectroscopy was applied to differentiate between antibiotic-sensitive and antibiotic-resistant strains in terms of the changes in chemical components (Fig. 4). The dominant absorption at 3,300 cm−1 observed for CET-induced Salmonella Typhimurium ATCC 19585 and CIP-induced Salmonella Typhimurium KCCM 4025 represents O-H stretching and N-H stretching modes, which are attributed to proteins.28–30 The absorption dominance at the region from 2,930 to 2,920 cm−1 corresponds to lipid aliphatic chains, asymmetric C-H stretching of CH3 in fatty acids. 29 Amide I was observed in the spectra of all strains except CET-induced Salmonella Typhimurium ATCC 19585 and CIP-induced Salmonella Typhimurium KCCM 4025 that are dominated by amide II and amide III. The carbohydrate region at 1,080 cm−1 was observed in the spectra of WT-, CET-, CIP-induced Salmonella Typhimurium ATCC 19585, WT CIP-resistant Salmonella Typhimurium ATCC 19585, WT Salmonella Typhimurium CCARM 8009, WT-, CET-, and CIP-induced Salmonella Typhimurium KCCM 4025. The bacterial surface cell components, including external appendages (pili and flagella), glycocalyx (capsule and slime layer), membrane proteins, and wall components (lipopolysaccharides), are altered in antibiotic-resistant bacteria. 31 FT-IR spectroscopy could detect the changes in bacterial cell components. Thus, the FT-IR spectral patterns can discriminate antibiotic-resistant bacteria according to the changes in cellular components. 32
The AcrAB–TolC efflux pump overexpressed in this study (Fig. 5) is associated with the broad substrate specificity to β-lactams, fluoroquinolones, macrolides, aminoglycosides, and tetracycline, which plays an important role in the development of multidrug resistance and pathogenicity in bacteria.33–35 The decreased expression of ompC results in the change in the membrane permeability, leading to the cross-protective responses to environmental stresses. 36 The development of multidrug resistance in bacteria can be attributed to the decreased permeability of porin channels, which can block the influx of various types of substrates such as β-lactams, chloramphenicol, fluoroquinolones, and tetracycline. 37 The rpoS gene in all strains was overexpressed under the exposure to sublethal conditions (pH 4.5, 4% NaCl, and heat at 42°C) (Fig. 5). The rpoS encoding σs RNA-polymerase subunit mediates the regulation of cross-protective responses to high osmolarity, heat, and acidic stresses.38,39 The environmental stresses, including antibiotic, starvation, acid, osmolality, and heat, trigger stress shock responses to regulate the expression of chaperones and proteases.40,41 The cross-protection against other stresses can be induced because of the presence of common regulatory mechanisms. 42 The relative gene expression level of stn in Salmonella Typhimurium strains varied in the environmental stress conditions (Fig. 5). This suggests that the expression of virulence factors in bacteria can be coordinately regulated depending on the types of cells and stresses.43,44 The highest expression level of fimA was observed in WT CIP-resistant Salmonella Typhimurium ATCC 19585 exposed to pH 4.5 and 4% NaCl, showing more than sixfold increase (Fig. 5). The fimA is directly related to the bacterial attachment and invasion, which were considerably increased under stressful conditions. 45
In conclusion, this study highlights the variability in adaptive responses of antibiotic-induced Salmonella Typhimurium to pHs (4.5–9.0), NaCl concentrations (0% to 5%), and temperature (42°C). The CET- and CIP-induced Salmonella Typhimurium strains could induce cross-protection against acidic, osmotic, and heat stresses depending on the antibiotic resistance status. The activation of cross-protective responses to stresses is linked to the development of antibiotic resistance in bacteria, leading to the changes in outer membrane permeability and efflux pump activity. The antibiotic resistance in Salmonella Typhimurium can be discriminated based on the physicochemical characteristics. The results obtained from this study might not be sufficient to conclude the clear relationship between antibiotic susceptibility and environmental stresses. However, this study pointed out that the ability to adapt to stresses was highly related to the altered antibiotic resistance in Salmonella Typhimurium. Therefore, this study would provide valuable insights into the induction of cross-protective responses of MDR Salmonella when exposed to different environmental stresses.
Footnotes
Acknowledgment
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2016R1D1A3B01008304).
Disclosure Statement
All authors declare that they have no conflicts of interest or any other competing financial interests to disclose.