
Abstract
Trueperella pyogenes is a common inhabitant of mucosal surfaces in animals and causes a variety of infections, including endometritis, mastitis, and liver abscessation, in dairy cows. Many antimicrobial agents are used for treatment of infections caused by T. pyogenes; however, antibiotic resistance has recently become a serious problem. The objective of this study was to characterize the effect of the efflux pump-encoding multidrug and toxic compound extrusion (MATE) gene on antibiotic resistance in T. pyogenes isolates from cows with signs of endometritis. As a compound from plants, luteolin showed antimicrobial activities in Escherichia coli and Staphylococcus aureus; therefore, we also investigated whether luteolin can eliminate antibiotic resistance. We constructed a MATE deletion mutant in BM-H06-3 to identify the function of MATE in antibiotic resistance. MATE mRNA expression was measured to identify the mechanism of luteolin in gentamicin resistance elimination effect in T. pyogenes. The T. pyogenes isolate BM-H06-3 became susceptible to gentamicin, amikacin, streptomycin, erythromycin, and roxithromycin after MATE deletion. No synergistic effect between luteolin and gentamicin was observed in eight isolates, which were randomly selected from 34 T. pyogenes isolates, but the isolates became susceptible to gentamicin after luteolin treatment at a subinhibitory concentration (1/4 minimum inhibitory concentration [MIC]) for 36 hr. Furthermore, luteolin can decrease MATE mRNA expression after luteolin treatment at a subinhibitory concentration (1/4 MIC). We found that the MATE gene was involved in antibiotic resistance and that luteolin induces a resistance elimination effect in T. pyogenes. Therefore, luteolin may be a potential agent to inhibit efflux pumps in multidrug-resistant T. pyogenes.
Introduction
T
Several strategies are involved in bacterial drug resistance: (i) producing enzymes to inactivate antibiotics, (ii) altering targets to reduce antibiotic affinity, and (iii) reducing drug accumulation by decreasing permeability or increasing efflux. 8 Some bacteria can confer antibiotic resistance by intrinsic features acquired by mutation or by the acquisition of exogenous genes. 9 Multidrug and toxic compound extrusion (MATE), which is a multidrug efflux pump, was found in Vibrio parahaemolyticus (NorM protein), the human liver, the mouse kidney, and plants. 8 Bacterial MATE transporters function as xenobiotic efflux pumps and play a crucial rule in the development of multidrug resistance against structurally diverse drugs, including antibiotic and DNA-damaging agents. 10
The MATE gene, which encodes an efflux pump, has been detected in the T. pyogenes DNA genome (GenBank No: CP012649.1). However, whether drug efflux pumps are involved in tetracycline and macrolide resistance in T. pyogenes has still not been elucidated.
Natural products from plants have recently attracted increasing interest in drug discovery. 11 Several plant-derived compounds, such as reserpine, have activity in inhibiting efflux pumps and are known as efflux pump inhibitors. 12 Flavonoids showed antibacterial activity against gram-positive bacteria. 13 Luteolin, a classic flavonoid compound from plants, showed antimicrobial activities by membrane-binding and ATPase-inhibiting activities, and the researchers also observed that luteolin induced the disruption of the cytoplasmic membrane of methicillin-resistant Staphylococcus aureus (MRSA). 14 Similarly, luteolin was reported to have antibacterial activity against MRSA clinical isolates. 15 These results suggest that luteolin can be a potential antimicrobial agent.
Many natural products, including luteolin, showed synergism with antibiotics, and a possible mechanism may be that natural products can inhibit the bacterial efflux pumps in MRSA.16,17 However, whether luteolin can inhibit efflux pumps remains to be elucidated. Therefore, we identified the function of the MATE transporter by the construction of a T. pyogenes mutant and further investigated the synergistic effect of luteolin and gentamicin against multidrug-resistant T. pyogenes. In addition, the mRNA level of the MATE gene was measured in this study.
Materials and Methods
Bacterial strains and growth conditions
The 34 T. pyogenes isolates were obtained from dairy cows with endometritis, 6 identified by colony morphology, Gram staining, and API kits (bioMérieux, France), and stored at −80°C in brain heart infusion (BHI) broth (AoBox Biotechnology, Beijing, China) containing 20% glycerol until further testing. One T. pyogenes isolate, namely BM-H06-3, was used to construct the MATE gene mutant. T. pyogenes ATCC 19411 (purchased from American Type Culture Collection, Manassas, VA) was selected as a reference strain.
T. pyogenes strains were cultured on BHI agar plates containing 5% rabbit blood at 37°C for 48 hr or in BHI broth containing 8% heat-inactivated fetal bovine serum (FBS; Tianjin Haoyang Biological Manufacture, Tianjin, China) and 0.11% v/v Tween 80 (Sigma, Beijing, China).
DNA extraction
The template DNA was prepared as previously described. 18 Briefly, a single colony was picked from a fresh bacterial culture on BHI agar (AoBox) plates, which contained 5% rabbit blood, and inoculated into 3 mL of sterile BHI broth at 37°C for 24 hr in an incubator. The DNA extraction was carried out by using a bacterial DNA extraction kit (Takara, Dalian, China) according to the manufacturer's protocol. All DNA preparations were stored at −20°C until use.
Screening for the MATE gene
Polymerase chain reaction (PCR) was used for the evaluation of the presence of the MATE gene. The MATE gene was amplified by a PCR system (PTC-100® thermal cycler, MA), and PCR amplifications were carried out in a 25 μL reaction mixture containing 0.1 μM each primer, 40 mM dNTPs, 2.5 μL of 10 × PCR buffer, 2.5 U of exTaq DNA polymerase (Takara), and 50 ng of template DNA. The thermal cycling condition was identical for all primers: a 3 min denaturation at 94°C followed by 30 cycles of 30 sec at 94°C, 45 sec at 64°C, 1 min at 72°C, and a 5 min final extension at 72°C. The amplified DNA products were confirmed by 1.0% (w/v) agarose gel electrophoresis and ethidium bromide staining, and the bands were visualized with an ImageMaster® VDS System (Amersham Pharmacia Biotech).
The amplified DNA products were sequenced by Sangon Biotech (Shanghai, China). The sequence comparison was performed using MEGA 7 v6.1, 19 and the phylogenetic tree was constructed with MEGA 6 v6.1 by using the maximum likelihood method. The primers used to amplify the MATE gene (size: 1,311 bp) are as follows: forward: ATGGTCAGCTTCCAGTCAG (5′ to 3′) and reverse: TCAAACTCGTGGTTGCCG (5′ to 3′).
Construction of BM-H06-3ΔMATE
A MATE-inactivated T. pyogenes mutant was constructed in the BM-H06-3 isolate (BM-H06-3ΔMATE) according to a previously described method.20,21 The reason we chose BM-H06-3 for the construction of the MATE-inactivated mutant is that this strain showed multidrug resistance, including resistance to aminoglycosides, tetracyclines, macrolides, and beta-lactams.
The MATE locus was identified on the chromosomal region of T. pyogenes strain BM-H06-3, and primers were designed to amplify the upstream region (MATEUF and MATEUR) and the downstream region (MATEDF and MATEDR) of the chromosome flanking the MATE locus. Kanamycin was used for mutant selection since BM-H06-3 was susceptible to kanamycin, so the kanamycin resistance gene cassette and its promoter were amplified by primers (KanRF and KanRD) from the Epicentre Tn5 transposon (Epicentre, Madison, WI). Restriction enzyme recognition sites (shown in Table 1) were incorporated in the primers to facilitate the cloning of the amplification products, and the fragments was amplified by PCR with Phusion DNA polymerase (Takara) under the following conditions: 98°C for 3 min; 35 cycles at 98°C for 10 sec, 56°C for 1 min, and 72°C for 30 sec; and a single cycle at 72°C for 10 min.
Underlined and bold sequences are restriction sites.
The PCR products were digested with appropriate restriction endonucleases (Promega, Madison, WI) according to the manufacturer's instructions. Both pBluescript SK+ and the upstream fragment of the MATE gene were digested with the endonucleases SalI/HindIII, and the upstream fragment was then cloned into pBluescript SK+. Similarly, the downstream fragment of the MATE gene was cloned into pBluescript SK+ after SalI/HindIII digestion. There was a HindIII restriction site in the constructed plasmid, and the kanamycin resistance cassette was inserted to yield the plasmid pBSMATE_Kan after digestion with HindIII. The MATE gene sequence is interrupted by the kanamycin resistance cassette in the plasmid pBSMATE_Kan, which provides a selection marker for the transformants.
The plasmid pBluescript SK+ acts as a suicide vector since it is based on ColE replication, which cannot occur within T. pyogenes, as reported previously. 21 Competent BM-H06-3 cells were prepared and transformed with pBSMATE_Kan by electroporation, as described previously. 22 Transformants were selected by growth on BHI agar with 10% FBS supplemented with 30 μg/mL kanamycin; PCR was subsequently used to confirm the loss of the MATE gene in T. pyogenes BM-H06-3.
Antimicrobial susceptibility test for BM-H06-3 and BM-H06-3ΔMATE
Susceptibility to antimicrobial agents was determined by the broth microdilution method as described by the Clinical Laboratory Standards Institute guidelines. 23 The concentration of bacteria was adjusted to ∼5 × 10 5 cfu/mL in Mueller–Hinton broth [MH(B), AoBox] containing 8% FBS, and trays were maintained in a normal atmosphere incubator for 24 hr at 37°C. T. pyogenes ATCC 19411 was used as a reference strain. Each experiment was carried out three times.
The antimicrobial agents tested in this study were as follows: penicillin, ampicillin, cefalexin, cefazolin, ceftiofur, gentamicin, amikacin, streptomycin, tetracycline, oxytetracycline, erythromycin, and roxithromycin. The antimicrobial agents were purchased from the China Institute of Veterinary Drugs Control.
Inhibition and checkerboard assays
According to our previous result, eight isolates were selected according to their resistance phenotype to investigate the interaction between luteolin and gentamicin; for example, HC03-1, HC03-7, HC-H03-3, and HC-H14-1 were resistant to no more than three antimicrobial agents, also resistant to gentamicin with minimum inhibitory concentrations (MICs) ≥32 μg/mL; whereas HC-H02-2, BM04-2, QMC01-3, and WBX02-1 were resistant to at least four antimicrobial agents, and these isolates were resistant to gentamicin with MICs ranging from 4 to 32 μg/mL.
MH(B) containing 8% FBS was selected as the medium for susceptibility testing. The concentration of bacteria was adjusted to ∼5 × 10 5 cfu/mL. Luteolin (Code No. SL8300) was purchased from Solarbio Life Science & Technology (Beijing, China). Luteolin was dissolved in dimethyl sulfoxide, and the pH was adjusted to 8.0 by using 0.5 mol/L NaOH. The susceptibility of the 34 T. pyogenes isolates to luteolin and gentamicin was carried out as previously described. 23 Trays were maintained in a normal atmosphere incubator for 24 hr at 37°C, and T. pyogenes ATCC 19411 was used as a reference strain. Each tray was performed at least three times.
A checkerboard method was used to study interactions of the luteolin and gentamicin combinations. 24 In each well of a 96-well microtiter plate, a total of 100 μL of antimicrobial solutions containing both luteolin and gentamicin was mixed with 100 μL of T. pyogenes culture (∼5 × 10 5 cfu/mL). The volume of each antimicrobial solution was adjusted with T. pyogenes culture to obtain a final overall antimicrobial concentration corresponding to 2 MIC, 1 MIC, 1/2 MIC, 1/4 MIC, and 1/8 MIC. Each experiment was repeated three times.
After determining the MIC using the above protocol, the fractional inhibitory concentration (FIC) index of a luteolin and gentamicin combination was calculated, and the FIC was calculated as follows: FIC = FICL+FICG, where FICL is the MIC of luteolin in combination/the MIC of luteolin alone and FICG is the MIC of gentamicin in combination/the MIC of gentamicin alone. Effects were considered “synergistic” if the FIC was ≤1.00 for a luteolin and gentamicin mixture, and “No interaction” if the FIC was ≥1.00.
Effect of luteolin at subinhibitory concentrations on resistant strains
Effects of luteolin at subinhibitory concentrations on resistant T. pyogenes isolates were examined. Five isolates were randomly selected from nine multidrug-resistant strains, and the effects of luteolin at 1/2 MIC, 1/4 MIC, 1/8 MIC, and 1/16 MIC were investigated on resistant strains, and then, the MATE gene expression was measured at 1/4 MIC.
Briefly, overnight cultures of T. pyogenes were adjusted to a final concentration of 5 × 10 5 cfu/mL, and luteolin was added to obtain final concentrations of 1/2 MIC, 1/4 MIC, 1/8 MIC, and 1/16 MIC; the inoculated cultures were then incubated at 37°C. At 18, 30, 36, 42, 48, 54, and 60 hr, 100 μL of the inoculated cultures was inoculated into 5 mL MH(B) containing 8% FBS for overnight culture at 37°C. The overnight culture was diluted to a final concentration of 5 × 10 5 cfu/mL, and the MIC of gentamicin against the diluted overnight culture was then measured. The MIC changes were determined after treatment with luteolin. Each experiment was repeated three times.
Real-time fluorescent quantitative PCR
T. pyogenes isolates that were treated by luteolin (1/4 MIC) were inoculated onto BHI agar plates, and the plates were kept in an incubator at 37°C for 48 hr; a single colony was then inoculated in BHI broth containing 8% FBS at 37°C. The isolates were prepared for RNA isolation after growth to exponential phase (OD600 = 0.5), and each isolate was measured from three independent cultures. Total RNA was extracted from strains by TRIzol reagent (Invitrogen, Carlsbad, CA) according to the manufacturer's instructions. The yield of total RNA was quantified at A260, and the RNA integrity was checked on a 1% (w/v) agarose gel (Sigma, China) stained with ethidium bromide. DNA contamination was removed by treatment with 2 U of RNase-free DNase I (Tiangen, Beijing, China) for 15 min at room temperature followed by heat inactivation for 10 min at 65°C in the presence of 2.5 mM EDTA (Sigma, China).
The cDNA was synthesized in a 20 μL reaction mixture containing 1 μg of RNA, 4 μL of 5 × reverse transcriptase buffer, 20 mM dNTPs, 20 U of RNase inhibitor, 50 pM random primer, and 10 U of avian myeloblastosis virus reverse transcriptase (Takara). A control PCR reaction without reverse transcriptase was performed to determine potential DNA contamination.
The mRNA expression levels of the MATE gene in isolates after treatment with luteolin (1/4 MIC) were measured, and the mRNA expression levels in isolates, which were cultured for five generations after treatment with luteolin (1/4 MIC), were also measured, as previously described. 18 Standard curves for real-time fluorescent quantitative PCR (RT-PCR) were constructed by serially diluting the plasmids as templates for the MATE gene and 16S rRNA. The mRNA expression of 16S rRNA was shown to be stable when treated by luteolin in T. pyogenes (data was not published). cDNAs of the target bacteria were used as templates for sample detection.
RT-PCR were performed using the SYBR Green PCR core reagent kit (ShineGene, Shanghai, China) according to the manufacturer's instructions. The reaction mixture (25 μL) consisted of 1.0 μL of template, 0.5 μL (10 mM) of each primer, 12.5 μL of 2 × Hot Start Fluo-PCR mixture, and 10.5 μL of ddH2O. DNA amplification and quantification were performed using a DNA Engine® Chromo 4™ Real-Time PCR detector (Peltier Thermal Cycler, Bio-Rad, Hercules, CA). Two negative controls without the target gene were subjected to the same treatment. The reaction conditions were as follows: a 5 min initial denaturation at 94°C; 30 cycles of denaturation at 94°C for 30 sec, annealing at 64°C for 30 sec, extension at 72°C for 1 min; melt curve reading for 2 sec every 0.2°C from 65°C to 95°C; and a final extension for 10 min at 72°C.
Data were analyzed using Opticon analysis software version 3.0 (Bio-Rad). The Ct values and copy numbers were calculated automatically. The primers used for RT-PCR are listed in Table 2. Each sample was analyzed in triplicate.
MATE, multidrug and toxic compound extrusion.
Statistical analyses
The mRNA expression levels of the MATE gene were compared (ANOVA) among groups. These data were analyzed using repeated-measures ANOVA followed by Fisher's protected least significant difference test. For all analyses, p < 0.05 was considered significant, and values are presented as the mean ± standard error of the mean.
Results
MATE gene screening and DNA sequence analysis
Most isolates were positive for the MATE gene, which had a prevalence of 94.12% (32/34) among all isolates. According to the BLASTN analysis, in the MATE gene-positive isolates, the identity to the MATE gene sequence in the genomic DNA of an isolate (GenBank No: CP012649.1) ranged from 96.34% to 99.47%. The maximum likelihood tree revealed that the isolates belonged to different clusters and that there was no relationship between the clusters and the antibiotic resistance phenotypes (shown in Fig. 1).
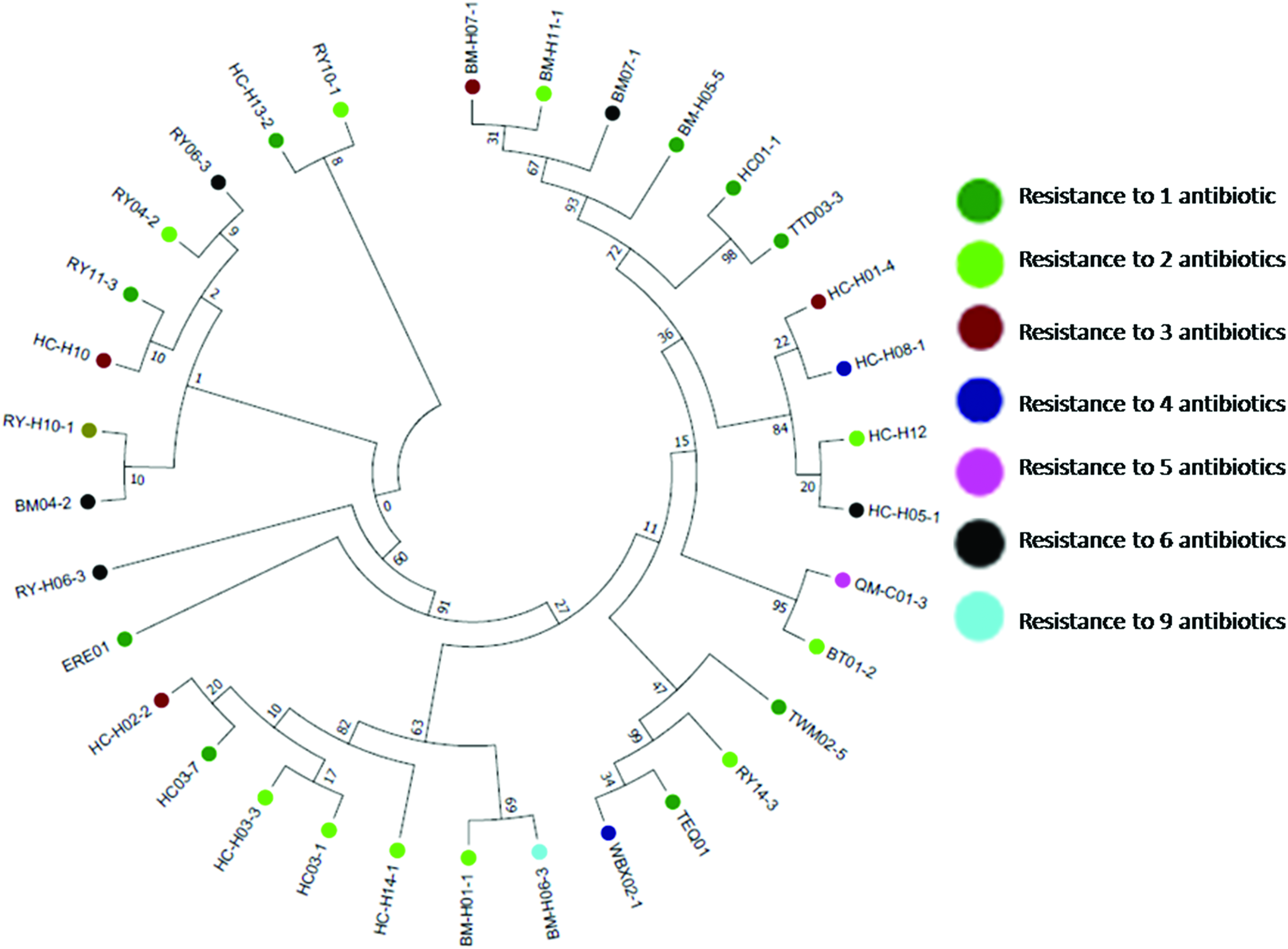
Phylogenetic tree constructed using maximum likelihood method and inferred with the Tamura–Nei model. The resistance phenotype of Trueperella pyogenes isolates are represented in different color shapes. Color images are available online.
The BM-H06-3 mutant became susceptible to antibiotics
To evaluate the effect of the MATE gene on antibiotic resistance in T. pyogenes, MIC measurements were carried out for both the BM-H06-3 and the BM-H06-3 mutant strains. The MICs of the mutant strain decreased for ceftiofur, gentamicin, amikacin, streptomycin, erythromycin, and roxithromycin. For example, the MIC of erythromycin decreased from 8 to 0.25 μg/mL, a 32-fold decrease; similarly, the roxithromycin MIC showed a 32-fold decrease. However, the MIC of amikacin decreased from 8 to 2 μg/mL, a decrease of only fourfold. However, the mutant strain did not show any MIC decrease for other antimicrobial agents, including penicillin, ampicillin, cefalexin, cefazolin, tetracycline, and oxytetracycline (shown in Table 3).
MIC and checkerboard assays for gentamicin and luteolin
The MICs for gentamicin and luteolin against the T. pyogenes strains are shown in Table 4. The MICs of gentamicin and luteolin against eight T. pyogenes isolates ranged from 1 to 32 μg/mL and from 39 to 78 μg/mL, respectively. When gentamicin was combined with luteolin, the FICs of gentamicin plus luteolin were 2 MIC, 1 MIC, 1/2 MIC, 1/4 MIC, or 1/8 MIC against nine strains (HC03-1, HC03-7, HC-H03-3, HC-H14-1, HC-H02-2, BM04-2, QMC01-3, WBX02-1, and ATCC 19411). All the FIC indexes of these combinations against the strains were >1.00, which indicated that there is “No interaction” between gentamicin and luteolin against T. pyogenes strains.
FIC, fractional inhibitory concentration; MIC, minimum inhibitory concentration.
Effect of luteolin on T. pyogenes
The effect of luteolin on gentamicin resistance elimination was evaluated at different concentrations against T. pyogenes. The 1/4 MIC dose showed the best resistance elimination effect (Table 5), and five of the nine isolates mentioned above (HC-H03-3, HC-H02-2, BM04-2, QMC01-3, and WBX02-1) showed decreased resistance to gentamicin. For example, the MIC of gentamicin against HC-H03-3 decreased from 32 to 1 μg/mL after treatment with 20 μg/mL luteolin for 36 hr, whereas the MIC of gentamicin against BM04-2 decreased from 4 to 2 μg/mL. In addition, the resistance elimination effect of luteolin showed a positive correlation with the level of antibiotic resistance in T. pyogenes.
Treatment with luteolin results in reduced MATE mRNA expression
Treatment of the resistant T. pyogenes isolates with luteolin led to a decrease in the expression of the MATE gene (Fig. 2). The expression levels of the MATE gene in the T. pyogenes isolates were significantly decreased (0.025- to 0.25-fold, corresponding to a 4- to 40-fold reduction), except in BM04-2, in which the reduction was ∼1.2-fold. After the isolates were cultured in BHI broth (containing 8% FBS) for five generations, the mRNA levels of the MATE gene in the isolates were ∼0.5–0.98, except for HC-H14-1, with an mRNA level of 0.07. After the isolates were cultured in BHI broth for five generations, the mRNA levels of the MATE gene in the isolates became comparable to those in the corresponding strains that were not treated with luteolin (data not shown).
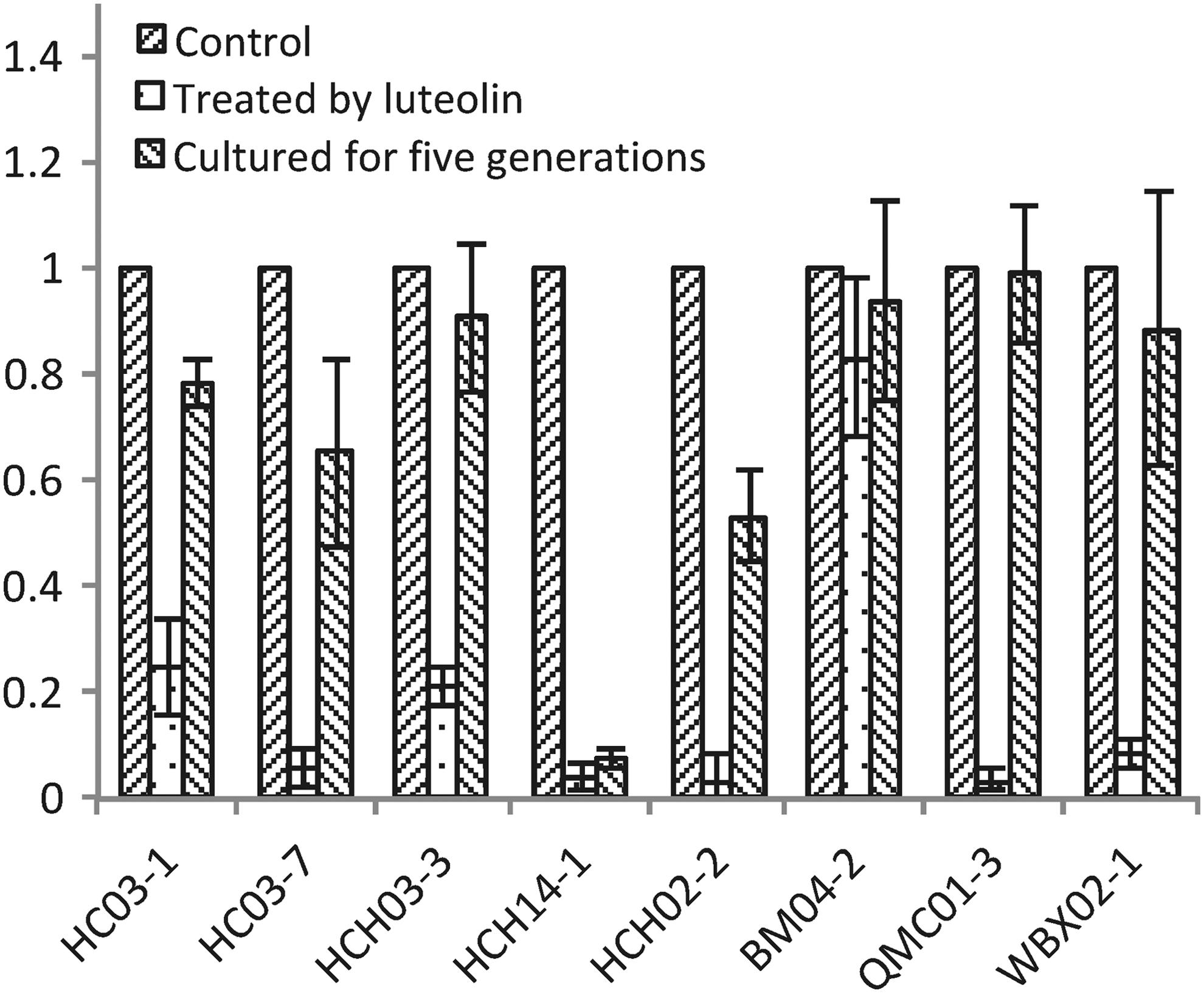
Luteolin at 1/4 minimum inhibitory concentration showed an inhibitory effect on the expression of MATE mRNA in T. pyogenes isolates. MATE, multidrug and toxic compound extrusion.
Discussion
Our data suggest that the MATE gene is responsible for antibiotic resistance in T. pyogenes, including ceftiofur, gentamicin, amikacin, streptomycin, erythromycin, and roxithromycin. Luteolin is a classic flavonoid compound from plants and possesses an antimicrobial activity. But there was no synergistic interaction between luteolin and gentamicin against T. pyogenes. However, T. pyogenes became susceptible to gentamicin after treated by luteolin at a subinhibitory concentration (1/4 MIC), and the mRNA level of the MATE gene increased to a level comparable to that in the resistant isolates after culture in medium without luteolin for five generations.
T. pyogenes commonly causes severe uterine disease in postpartum dairy cattle, with tissue damage and inflammation of the endometrium, 20 and multidrug resistance has repeatedly been reported worldwide.6,25 We studied the effect of the MATE gene, which encodes a multidrug efflux pump, on antibiotic resistance in T. pyogenes. In T. pyogenes, the MATE gene is involved in antibiotics resistance such as ceftiofur, gentamicin, amikacin, streptomycin, erythromycin, and roxithromycin.
According to phylogenetic tree, no relationship was found between the clusters and the antibiotic resistance phenotypes. The increases in susceptibility observed in the MATE gene deletion strain suggest that these antibiotics may be substrates of the efflux pump encoded by the MATE gene in T. pyogenes. Conversely, it is possible that neither of tetracycline or oxytetracycline is transported preferentially by the efflux pump encoded by the MATE gene.
Luteolin showed the antibacterial activity against T. pyogenes (with MICs ranging from 39 to 78 μg/mL), and similar results were observed against other bacteria. For example, luteolin showed favorable antibacterial activity in vitro against E. coli. 26 There is synergism between flavonoids and antibiotics, including methicillin, streptomycin, ofloxacin, levofloxacin, vancomycin, erythromycin, and kanamycin, which can result in an increase in the susceptibility of MRSA to these antibiotics. 27 A possible mechanism contributing to the synergistic effect could be the inhibitory effect of flavonoids on efflux pumps, which leads to the increase in the susceptibility of MRSA. 16 As a flavonoid, luteolin can increase the susceptibility of MRSA to conventional antibiotics. 17 Researchers hypothesize that luteolin may alter the permeability of the outer membrane and inner membrane. 26
Gentamicin needs to irreversibly bind to the 30S subunit of the bacterial ribosome after penetrating the cell wall. Theoretically, there is a synergistic effect between luteolin and gentamicin. However, the results of the checkerboard assays showed that luteolin did not enhance the activity of gentamicin against T. pyogenes. Interestingly, the isolates became susceptible to gentamicin after treatment with luteolin at a subinhibitory concentration. Our results indicated that the mRNA expression levels of MATE in the isolates decreased after treatment with luteolin, which is contributing to the gentamicin resistance elimination.
The clinical use of a combination of luteolin and gentamicin when combating infections caused by multidrug-resistant T. pyogenes is reasonable. We assume that luteolin decreased the expression of the efflux pump, and gentamicin thus accumulated in T. pyogenes; however, the concentration of gentamicin in T. pyogenes needs to be measured to validate our assumption. The mRNA expression levels of the MATE gene among the isolates became comparable with those in the resistant isolates after cultured for five generations; this result indicated that the resistance elimination effect of luteolin is reversible in T. pyogenes and that the mechanism needs to be further identified.
Conclusion
From this study, the following may be concluded: (i) the MATE gene is responsible for the resistance of T. pyogenes to antibiotics, including gentamicin, streptomycin, erythromycin, and roxithromycin; (ii) luteolin has the potential to restore the effectiveness of gentamicin against T. pyogenes; and (iii) this phenomenon may be attributed to the inhibitory effect of luteolin on the expression level of the MATE gene. Therefore, luteolin may offer the potential for development as a possible adjunct to gentamicin in the treatment of multidrug-resistant T. pyogenes infections.
Footnotes
Acknowledgment
This work was supported by the National Natural Science Foundation of China (No. 31572564 and No. 31772795).
Disclosure Statement
No competing financial interests exist.