
Abstract
The Klebsiella pneumoniae carbapenemase (KPC)-producing K. pneumoniae has become a serious problem because the species is wide ranging and there are few treatment options. Fosfomycin has attracted renewed interest in combination therapy for infections caused by KPC-producing K. pneumoniae isolates. Because of the increasing use of fosfomycin, resistant isolates have been continually reported in carbapenem-resistant K. pneumoniae (CRKP). At present, multiple mechanisms can result in fosfomycin resistance. However, there is limited knowledge with respect to plasmid-mediated fosfomycin resistance gene (fosA3) determinants in KPC-producing K. pneumoniae isolates. In this study, a total of 101 CRKP strains were collected from four hospitals in Zhejiang province from January 2013 to August 2014; 28.7% (29/101) of CRKP isolates were resistant to fosfomycin. Gene fosA3 was detected in 29 fosfomycin-resistant KPC-producing K. pneumoniae isolates, whereas genes fosA, fosB, fosB2, fosC, fosC2, and fosX were all negative among the resistant isolates. In addition, among 29 fosfomycin-resistant KPC-producing K. pneumoniae isolates, pulsed-field gel electrophoresis (PFGE) analysis revealed five pulsotypes. S1-PFGE and Southern blot showed that the fosA3 gene was located on an approximately 140-kb plasmid in all isolates. Eight of the 29 isolates (27.6%) tested could successfully transfer their fosfomycin-resistant phenotype to Escherichia coli strain J53. All fosA3-positive isolates were determined to have an identical genetic background, IS26-tetR-cadC-orf1-fosA3-IS26, which is the same as that of the fosA3-positive plasmid pFOS18 in China. The primary resistance mechanism to fosfomycin was caused by a plasmid-mediated fosA3. Furthermore, it is noteworthy that the plasmid genetically carrying a combination of the fosA3 and blaKPC-2 genes could accelerate the spread of antibiotic resistance. Effective and persistent monitoring and surveillance will be vital to prevent further dissemination of these resistance genes.
Introduction
C
Fosfomycin, a naturally occurring antimicrobial agent, had a broad spectrum of antibacterial activity against Gram-positive and Gram-negative bacteria and enters cells by active transport through the L-a-glycerophosphate and the hexose-6-phosphate uptake systems. 8 Fosfomycin is able to bind irreversibly to UDP-N-acetylglucosamine enolpyruvyl transferase (MurA) to inhibit the synthesis of peptidoglycan. 9
However, with the increased application of fosfomycin, resistant isolates have been continually reported. Multiple mechanisms can result in fosfomycin resistance, such as antibiotic-modifying enzymes, target site modification, or decreased permeability. 10 Currently, known mechanisms of fosfomycin resistance in K. pneumoniae are primarily involved in fosfomycin-inactivating glutathione S-transferase enzymes. 11 Fosfomycin-modifying enzymes can be disseminated by conjugative plasmids or mobile element, and have resulted in serious challenges in clinics. However, there is limited knowledge of plasmid-mediated fosfomycin-modifying enzymes in KPC-producing K. pneumoniae isolates.
In this study, we aimed to investigate fosfomycin susceptibility and plasmid-mediated resistance determinants in clinical KPC-producing K. pneumoniae isolates.
Materials and Methods
Bacterial strains
During the period from January 2013 to June 2014, a total of 101 nonrepetitive KPC-producing K. pneumoniae were collected and identified from four different hospitals in Zhe Jiang province in China. The sources of the isolates were urine (n = 23), sputum (n = 53), abscess (n = 9), blood (n = 11), pus (n = 3), and other patient samples (n = 2).
Initially, bacterial identification was performed using a VITEK®2 system (bioMérieux, Lyons, France). Strain Escherichia coli J53 was used as a recipient in the conjugation experiments, and E. coli ATCC 25922 and K. pneumoniae ATCC 700603 were used as quality control strains for antimicrobial susceptibility testing.
Antimicrobial susceptibility testing
Minimum inhibitory concentrations (MICs) of all isolates were determined using agar dilution method according to the approved standard of the Clinical and Laboratory Standards Institute 2017 guidelines, 12 including the following antimicrobials: ceftazidime, cefepime, amikacin, imipenem, ciprofloxacin, norfloxacin, and cefoperazone/sulbactam. Tigecycline and colistin were determined using broth microdilution method. In addition, the MIC of fosfomycin was determined by the agar dilution method using supplementation with 25 mg/L glucose-6-phosphate, and the susceptibility results were analyzed according to the CLSI guidelines, 12 except tigecycline and colistin, for which the European Committee on Antimicrobial Susceptibility Testing breakpoints were used (www.eucast.org/clinical_breakpoints).
Polymerase chain reaction screening
The KPC-producing K. pneumoniae isolates were confirmed to amplify the blaKPC-2 gene. 13 All fosfomycin-resistant isolates were screened for the plasmid-encoded fosfomycinase genes (fosA, fosA3, fosB, fosB2, fosC, fosC2, and fosX). Primers for the aforementioned resistance genes were used according to previous studies.14–16 The positive products of polymerase chain reaction (PCR) amplification were sequenced using an ABI 3730 sequencer (Applied Biosystems, Foster City, CA). Nucleotide sequences were compared using BLAST (http://blast.ncbi.nlm.nih.gov/Blast.cgi).
Bacterial genotyping
Twenty-nine fosA3-producing isolates were subjected to pulsed-field gel electrophoresis (PFGE) analysis for detecting the clonal relatedness of the isolates. Briefly, genomic DNA was digested by the XbaI (TaKaRa, Japan), the producing DNA fragments were further separated by electrophoresis in 1% agarose III (Sangon, Shanghai, China) in 0.5 × TBE buffer with a CHEF Mapper XA apparatus (Bio-Rad, Hercules, CA) for 22 hr at 14°C and 6 V/cm, with alternating pulses at a 120° angle in a 5–35 sec pulse time gradient. The Salmonella enterica serotype Braenderup H9812 was used as the size marker.
Multilocus sequence typing (MLST) was also used for molecular typing. Bacterial genomic DNA was extracted from these isolates. Seven housekeeping genes of K. pneumoniae (gapa, infb, mdh, pgi, phoe, rpob, and tonB) were amplified by PCR, and the products were sequenced to analyze the sequence type according to protocols available at the MLST Pasteur website (http://bigsdb.pasteur.fr/perl/bigsdb/bigsdb.pl?db=pubmlst_klebsiella_seqdef_public).
Conjugation experiments
Conjugation experiments were carried out by broth mating to test the transmissibility with sodium azide-resistant E. coli J53 as the recipient strain. Transconjugants were selected on Mueller-Hinton agar plates containing supplemented with 100 μg/mL sodium azide and 64 μg/mL fosfomycin. PCR analysis and antimicrobial susceptibility testing were performed to confirm the resistance gene transfer. Antimicrobial susceptibility testing for the transconjugants were compared with E. coli J53 and their respective donors to further confirm the transferable resistance genes.
Southern blotting
To confirm the plasmid location of fosA3 and blaKPC-2 genes, genomic DNA digested with S1 nuclease (TaKaRa, Japan) was electrophoresed on a CHEF-mapper XA PFGE system (Bio-Rad) for 22 hr at 14°C with run conditions of 6 V/cm and pulse times from 2.16 to 63.8 sec. The DNA fragments were transferred to a positive-charged nylon membrane (Millipore) and then hybridized with a digoxigenin-labeled fosA3 and blaKPC-2-specific probes. An NBT/BCIP color detection kit (Roche, Germany) was then used to detect the fragments. The Salmonella enterica serotype Braenderup H9812 was used as the size marker.
Analysis of the genetic environment flanking the fosA3 gene
Primers of genetic environment flanking the fosA3 gene were used according to the previous study. 17 The PCR amplification products were sequenced and blasted with sequences deposited in the BLAST database.
Results
Fosfomycin susceptibility testing
Of the 101 KPC-producing K. pneumonia isolates tested, the MICs for fosfomycin ranged from 2 mg/L to greater than or equal to 1,024 mg/L (Supplementary Fig. S1. Twenty-nine (28.7%) isolates were resistant to fosfomycin. The MIC50 and MIC90 values of the strains were 64 mg/L and greater than or equal to 1,024 mg/L, respectively. All resistant isolates had high MICs of fosfomycin (greater than or equal to 1,024 mg/L) (Supplementary Fig. S1). In addition, the fosfomycin resistance rate of KPC producers in four hospitals was different, varying from 17.8% to 65.0%. The fosA3-positive rate in different hospitals was also different, varying from 27.3% to 60.0%. Furthermore, these strains present a multidrug-resistant phenotype (Table 1), including ceftazidime, cefepime, imipenem, nitrofurantoin, amikacin, cefoperazone/sulbactam, and ciprofloxacin, with the exception of colistin- and tigecycline-classified extensively drug resistant (XDR) bacteria according to the recently proposed international classification scheme. 18
AMK, amikacin; CAZ, ceftazidime; CIP, ciprofloxacin; CPS2/1, cefoperazone/sulbactam 2:1; CST, colistin; FEP, cefepime; FOS, fosfomycin; IPM, imipenem; MIC, minimum inhibitory concentrations; NIT, nitrofurantoin; TGC, tigecycline.
Fosfomycin resistance gene screening
Gene fosA3 was detected in these 29 (28.7%) fosfomycin-resistant KPC-producing K. pneumonia isolates, whereas the fosA, fosB, fosB2, fosC, fosC2, and fosX genes were all negative among the resistant isolates. Twenty-nine randomly selected fosfomycin-sensitive K. pneumoniae isolates did not contain fos genes. Notably, isolates carrying the fosA3 gene showed high-level fosfomycin MIC values in KPC-producing K. pneumoniae.
Clonal relatedness
According to the PFGE patterns, phylogenetic clonal analysis showed that the 29 fosA3-positive KPC-producing isolates were divided into five clonal groups (groups I–V) and seven singletons (S) using a similarity index (SAB) of 0.8 (Fig. 1). Clonal group I primarily comprised seven isolates that were detected in three hospitals. Clonal groups II, III, IV, V, and S contained 4, 2, 6, 3, and 7 isolates, respectively. In addition, all these fosA3-positive KPC-producing isolates belonged to ST11 clones.
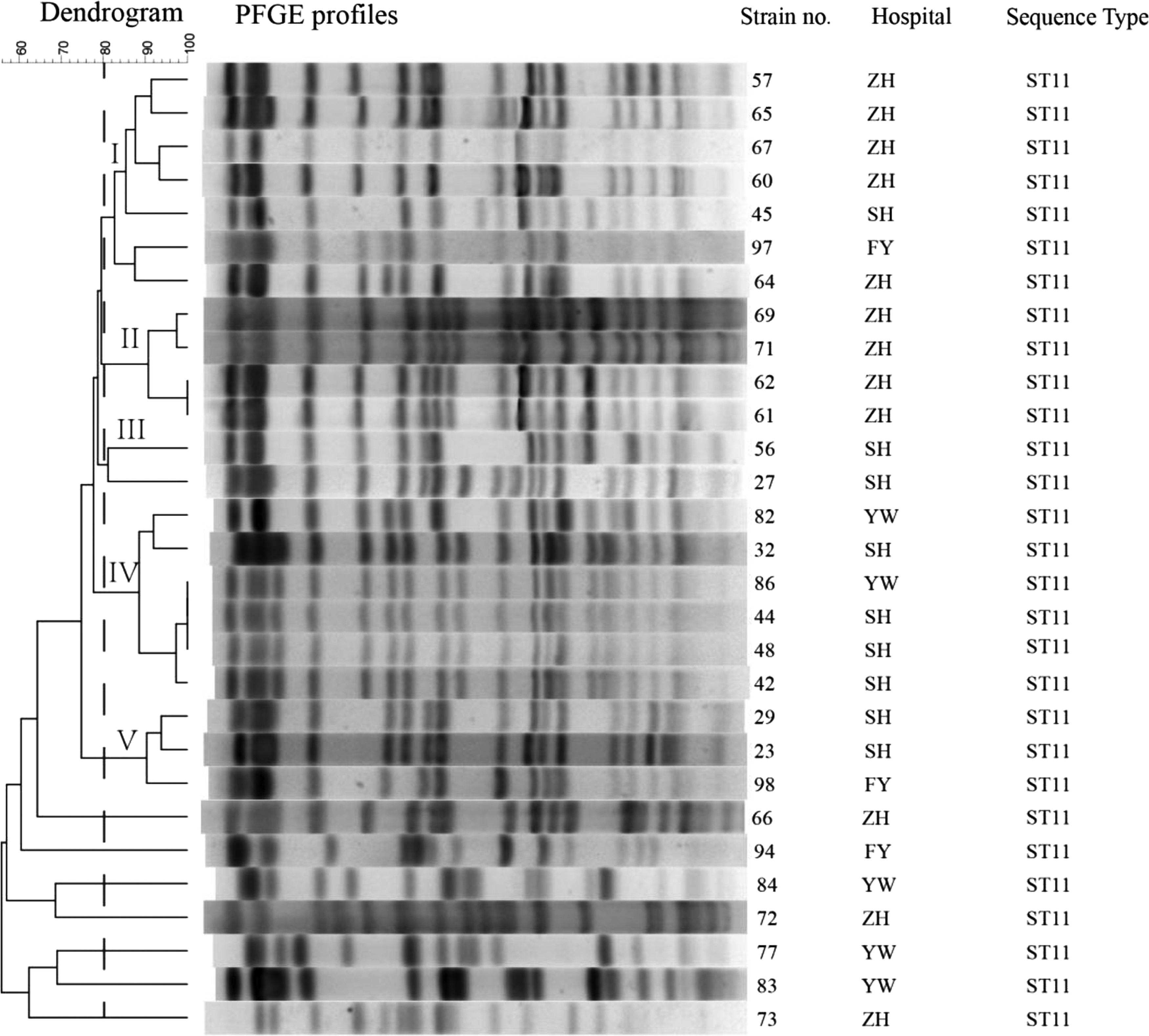
The dendrogram is based on the similarity of PFGE patterns from 29 fosA3-positive clinical CRKP isolates. The right illustrates results from isolate name, hospitals, and sequence type. The left illustrates results from clonal groups (groups I–V) and seven singletons. CRKP, carbapenem-resistant Klebsiella pneumoniae; PFGE, pulsed-field gel electrophoresis.
Location of the fosA3 gene
S1-PFGE followed by Southern blot analysis showed that the fosA3 gene was located on plasmids of the same size (∼140 Kb) (Fig. 2). The filter-mating experiments were carried out to confirm the transferability of these fosA3 plasmids. Of the 29 fosA3-positive strains, 8 (27.6%) fosA3 genes were successfully transferred to the recipient E. coli J53 strain in the filter-mating tests (Table 1). The fosfomycin MICs of transconjugants were significantly elevated compared with those of the recipient E. coli J53 strain. In addition, the probe for blaKPC-2 showed the same hybridization signal as the fosA3 gene (Fig. 2), indicating that two resistance genes could locate in a single plasmid.
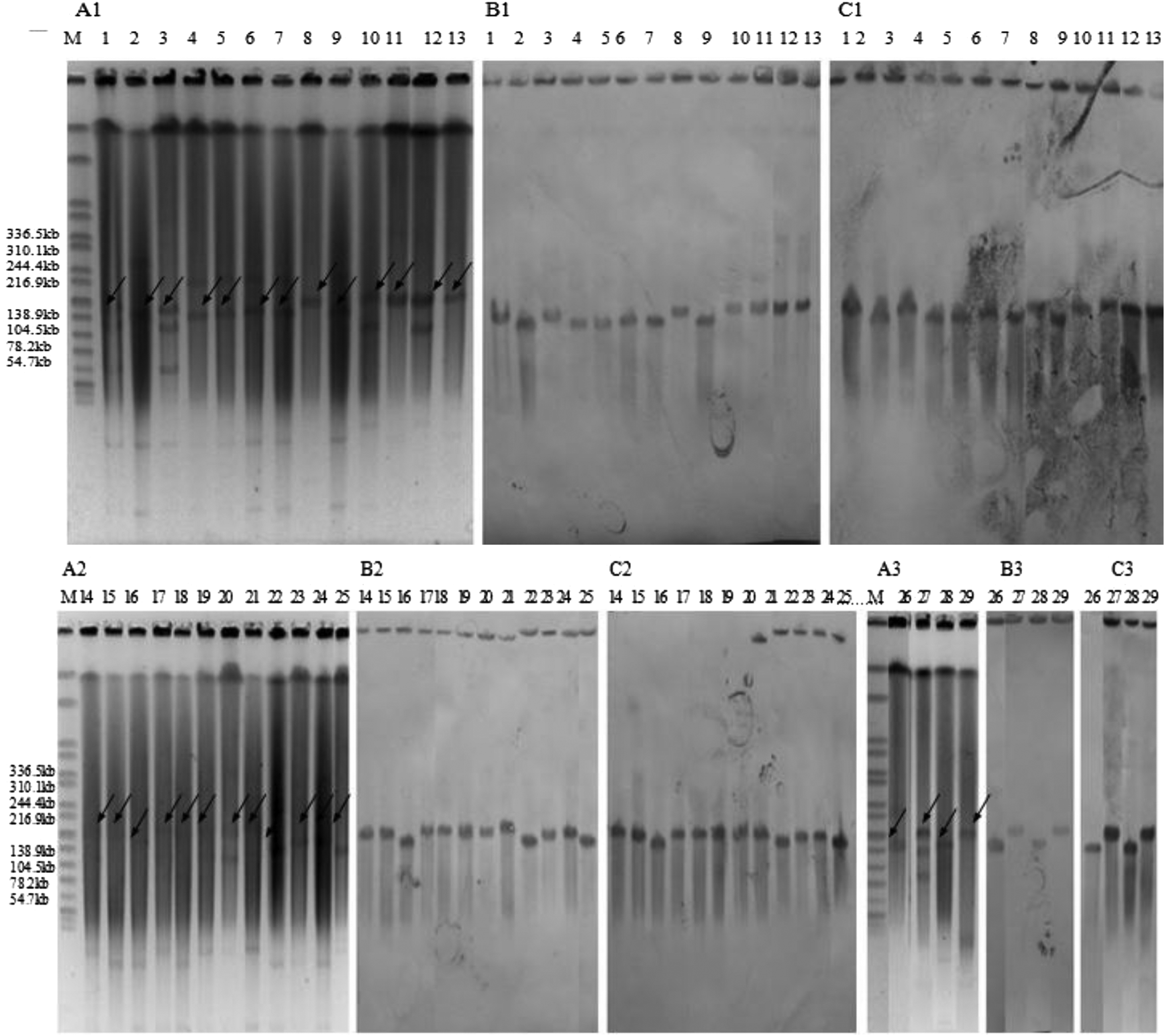
S1-digested plasmid DNA and Southern blot hybridization of blakpc-2- and fosA3-positive isolates. Bands with arrows pointing to them showed positive signals in Southern blot hybridization with the blakpc-2 and fosA3 probes. S1-PFGE was shown in
Genetic surroundings of fosA3
All fosA3-positive isolates were determined to have an identical genetic background, IS26-tetR-cadC-orf1-fosA3-IS26, which is the same as that of fosA3-positive plasmid pFOS18 in China 7 (GenBank Accession No. KJ653815).
Discussion
Most studies have shown favorable susceptibility of K. pneumoniae isolates to fosfomycin, such as those conducted in Thailand (90.7%; 39/43) 19 and Spain (97%; 134/138), 20 and K. pneumoniae isolates containing extended-spectrum beta-lactamase (ESBL) and metallo-β-lactamase in Greece (100%; 30/30). 21 Thus, fosfomycin has been suggested as a possible auxiliary drug to treat KPC-producing Klebsiella pneumoniae infections. Pontikis et al. reported that intravenous treatment of nosocomial infections caused by carbapenem-resistant K. pneumoniae has suggested that fosfomycin might have a role in the treatment of CRKP. 4 In addition, all patients had a good bacteriological and clinical outcome with this treatment. 4 However, recent studies have revealed that the lower susceptibility rate of fosfomycin occurs in ESBL-producing and KPC-producing K. pneumoniae isolates.7,22 Liu et al. 22 and Jiang et al. 7 reported that 42.4% and 60.8% of ESBL-producing and KPC-producing K. pneumoniae isolates were resistant to fosfomycin, respectively. These inconsistent results indicate that the sensitivity of fosfomycin to K. pneumoniae isolates, especially to multidrug-resistant K. pneumoniae, requires further evaluation. This study demonstrated that 28.7% (29/101) of KPC-producing K. pneumoniae isolates were resistant to fosfomycin (MIC >256 mg/L). This result is different from reports by Liu et al. 22 and Jiang et al., 7 but similar to a study by Lu et al., 17 who reported that 30 (27.8%) isolates were not susceptible to fosfomycin among 108 ESBL-producing K. pneumoniae isolates tested. The different fosfomycin susceptibilities reported in these studies might be the result of different MIC breakpoints chosen in the respective studies (EUCAST: ≤32 mg/L, susceptible; ≥64 mg/L, resistant) and (CLSI: ≤128 mg/L, susceptible; ≥256 mg/L, resistant). In addition, fosfomycin susceptibility testing methodology suggested by CLSI and EUCAST for specific bacteria is also different. Thus, it is essential to consider this factor in the determination and unification of clinical breakpoints of fosfomycin.
MLST and PFGE experiments were performed to analyze the clonal relatedness of fosA3-positive KPC-producing isolates. According to the MLST results, ST11 was the only clone in our study that is consistent with the report that ST11 was the primary sequence type in Mainland China and involved in KPC. 23 Notably, ST11 K. pneumoniae has shown to be a good colonizer to capture the resistance genes. 24 In our study, these ST11 type fosfomycin resistance KPC-producing isolates owned fosA3 gene and classified as XDR bacteria, indicating that ST11 type K. pneumoniae might result in so-called “superbug” isolates.
The PFGE typing results further showed that fosfomycin resistance isolates identified in this study might be predominantly caused by clonal dissemination. Some clones were present in different hospitals, resulting in the wide dissemination of fosfomycin resistance. For example, strains 86 and 44, which were isolated from different hospitals, have an identical PFGE profile (Fig. 1). Furthermore, the fosA3 gene was detected in all fosfomycin-resistant isolates and was located in a plasmid. Compared with chromosomal-mediated fosfomycin resistance in K. pneumoniae, 11 the plasmid-mediated fosfomycin resistance gene (fosA3) was able to produce higher MICs for fosfomycin. Furthermore, the fosfomycinase and blaKPC genes are usually identified in the same isolates in China. Our findings reveal that the fosA3 gene (28.7%, 29/101) was co-disseminated with blaKPC-2. In this study, Southern blot analysis showed that fosA3 and blaKPC-2 were both located in the same sized plasmid. Recently, fosA3 and blaKPC-2 genes located on different transposon systems in a single plasmid have been reported in China. Notably, PCR screening in this study determined that the fosA3-positive and KPC-producing clinical isolates carried the IS26-fosA3-IS26-like structure, indicating that this mobile element plays an important role in the transfer resistance of fosfomycin. In addition, based on antimicrobial susceptibility of fosA3-producing transconjugants, aminoglycosides resistance gene may be co-disseminated with blaKPC-2 and fosA3. Thus, PCR screening was performed to detect aminoglycoside resistance gene. In this study, gene rmtB was detected in all aminoglycoside resistance transconjugants by PCR screening (Supplementary Fig. S2). Overall, these resistance genes could spread together globally through plasmid further transfer, which resulting in KPC-producing K. pneumoniae isolates difficult-to-treat pathogen due to limited treatment options.
Conclusions
In this study, 28.7% (29/101) of KPC-producing K. pneumoniae isolates were resistant to fosfomycin. The only resistance mechanism to fosfomycin was caused by a plasmid-mediated fosA3. Furthermore, it is noteworthy that plasmid carries a combination of the fosA3, rmtB, and blaKPC-2 genes, which could accelerate the spread of antibiotic resistance. Effective and persistent monitoring and surveillance will be vital to prevent further dissemination of these resistance genes.
Funding
This study was supported by the Natural Science Young Foundation of Zhejiang Province, China (LQ17H190006), the Medical and Health Research Project of Zhejiang Province, China (2017KY224), and General scientific research project of Zhejiang provincial education department (Y201534683). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.