
Abstract
The increasing number of carbapenem-resistant Acinetobacter baumannii clinical isolates is a major concern, which restricts therapeutic options for treatment of serious infections caused by this emerging pathogen. The aim of this work is to assess the antimicrobial resistance profile and identify the molecular mechanisms involved in carbapenem resistance in A. baumannii isolated from different clinical sources in Mansoura University Hospitals, Egypt. Antimicrobial susceptibility testing has shown that resistance to carbapenem has dramatically increased (98%) with concomitant elevated levels of resistance to quinolones, trimethoprim/sulfamethoxazole, and aminoglycosides. Polymyxin B and colistin are considered the last resort. Random amplified polymorphic DNA (RAPD) typing method revealed great diversity among A. baumannii isolates. Coexistence of diverse intrinsic and acquired carbapenem-hydrolyzing β-lactamases has been detected in the tested isolates: Ambler class A: blaKPC (56%) and blaGES (48%), and Ambler class B: blaNDM (30%), blaSIM (28%), blaVIM (20%), and blaIMP (10%). Most isolates (94%) carried blaOXA-23-like and blaOXA-51-like simultaneously. blaOXA-23-like was preceded by ISAba1 providing a potent promoter activity for its expression. Sequencing analysis revealed that ISAba1 has been also inserted in carbapenem resistance-associated outer membrane protein (OMP) (carO) gene in three isolates, two of which were clonal based on RAPD typing, leading to interruption of its expression as confirmed by SDS-PAGE analysis of OMP fractions. Carbapenem resistance genes are widely distributed among A. baumannii clinical isolates from different clinical sources. Therefore, enhanced infection control measures, effective barriers, and rational use of antimicrobials should be enforced in hospitals for minimizing the widespread resistance to carbapenems and all other antibiotics.
Introduction
A
Resistance to carbapenems is mainly mediated by the production of plasmid- and chromosome-encoded carbapenem-hydrolyzing β-lactamases, 6 and reduced permeability as a result of porin loss or modification. 7 Efflux pump overexpression and penicillin-binding proteins alteration have been found to play a role in developing resistance to carbapenems also. 8 Resistance to carbapenems is most frequently mediated by the enzymatic hydrolysis of the drugs. Carbapenemases belong to three molecular classes: the Ambler class A (including KPC and GES), Ambler class B (including IMP, VIM, SIM, and NDM), and Ambler class D (CHDLs or OXA) β-lactamases. They have emerged and dispersed worldwide. CHDLs or oxacillinases can be intrinsic (OXA-51 like) or acquired (OXA-23 like, OXA-24 like, OXA-58 like, and OXA-143 like). 6 Overexpression of CHDLs is frequently associated with insertion elements situated upstream of those genes conferring strong promoter activity. 9
Carbapenemases have become a major healthcare problem globally due to their association with resistance to other antimicrobial classes, including fluoroquinolones, aminoglycosides, and trimethoprim/sulfamethoxazole, reducing the possibility of treating infections caused by multidrug-resistant strains. 5 This study aims to assess the antimicrobial susceptibility profile and explore the main resistance mechanisms to carbapenems in clinical isolates of A. baumannii collected from Mansoura university hospitals, Egypt.
Materials and Methods
Bacterial isolates
A. baumannii isolates recovered from patient specimens in Mansoura University Hospitals between December 2015 and June 2016 were included in the study. Isolates were identified as A. baumannii according to standard microbiological techniques, including colony morphology, Gram stain, and biochemical reactions, 10 and molecular-based methods.11–14 A pair of primers specific to A. baumannii was used to amplify a single 722 bp fragment of the gltA gene encoding bacterial citrate synthase (Table 1).11,12 Moreover, blaOXA-51-like gene that is intrinsic to A. baumannii species was detected by PCR (Table 1).13,14 Therefore, whole-cell DNA of A. baumannii isolates was prepared by suspending one fresh colony into 100 μL of sterile distilled water and heating at 95°C for 10 min. After centrifugation, the supernatant was stored at −20°C. The thermocycling conditions were as follows: initial denaturation at 95°C for 5 min; followed by 30 cycles of 95°C for 60 sec, variable °C for 30 sec, and 72°C for 60 sec; and final extension at 72°C for 7 min (Table 1). The PCR products were subjected to 1.2% agarose gel electrophoresis and their sizes were confirmed under UV light (300 nm), and scanned in gel documentation and analysis system (Model Gel Doc 1.4, 1189; AccuLab®, USA).
Molecular typing
Molecular typing of A. baumannii clinical isolates was performed by random amplified polymorphic DNA (RAPD) method as described previously. 15 Banding patterns were analyzed using GelJ software version 2.0. 16 Similarity clustering analysis was performed using unweighted pair-group method with arithmetic mean and Dice coefficient. Clinical isolates with a similarity coefficient ≥85% were considered clonal.
Antimicrobial susceptibility testing
The antibiotic susceptibility profile was determined using the disk diffusion method 17 on Mueller-Hinton agar plates. The inhibition zones were interpreted according to the recommendations of Clinical and Laboratory Standard Institute. 18 Imipenem (10 μg) and Meropenem (10 μg) were used. Susceptibility to other classes of antibiotics was simultaneously tested, including Ampicillin-Sulbactam (10 μg/10 μg), piperacillin-tazobactam (100/10 μg), cefoxitin (30 μg), cefotaxime (30 μg), ceftazidime (30 μg), cefepime (30 μg), aztreonam (30 μg), polymyxin B (300 U), colistin (10 μg), gentamicin (10 μg), amikacin (30 μg), trimethoprim/sulfamethoxazole (1.25/23.75 μg), levofloxacin (5 μg), and ciprofloxacin (5 μg). All antibiotic discs were obtained from Oxoid, United Kingdom.
Phenotypic detection of carbapenem-hydrolyzing β-lactamases
Phenotypic detection of carbapenemases (modified Hodge test)
Phenotypic detection of carbapenemases was done using modified Hodge test according to the recommendations of Clinical and Laboratory Standard Institute. 18 Standard suspension 0.5 McFarland of Escherichia coli ATCC 25922 was prepared in broth and diluted 1:10 in broth. Mueller-Hinton agar plates were inoculated and allowed to dry for 3:10 minutes. Meropenem disk was placed on the center of the plate. Using a loop or a swab, 3:5 colonies of test organism were inoculated in a straight line out of the edge of the disk. The plates were incubated at 37°C for 16–20 hrs. Enhanced growth indicates positive carbapenemase production, while no enhanced growth indicates negative carbapenemase production.
Phenotypic detection of metallo-β-lactamases
Screening of metallo-β-lactamase (MBL) production was performed as described previously, utilizing IMP-EDTA synergy test. 19 Two imipenem (IMP, 10 μg) discs were placed on the surface of an inoculated MHA plate 25 mm apart and 2 μL sterile 0.5 M EDTA (Sigma Chemicals, St. Louis, MO) were added on one disc. The inhibition zones of IMP and IMP/EDTA discs were measured and compared after overnight incubation at 37°C. An increase in the inhibition zone around IMP/EDTA ≥7 mm compared to IMP alone is indicative of MBL production.
Molecular characterization of genes encoding carbapenem-hydrolyzing β-lactamases
The prevalence of carbapenem resistance genes among A. baumannii isolates was investigated by PCR using My Taq™ Red Master Mix (Bioline, United Kingdom). The PCRs were conducted on PCR thermal cycler machine (FPROGO2D; Tchne LTD., Oxford Cambridge, United Kingdom).
PCR was performed to detect metallo-β-lactamases (Class B β-lactamases), including blaIMP (imipenemase), 20 blaVIM (Verona integron-encoded metallo-β-lactamase), 21 blaNDM (New Delhi metallo-β-lactamase), 22 and blaSIM (Seoul imipenemase) 23 genes, and Class A β-lactamases, including blaKPC (Klebsiella pneumoniae carbapenemase) 24 and blaGES/IBC (Guiana extended-spectrum/integron-borne cephalosporinase). 25
Multiplex PCR was used to detect five families of class D β-lactamases (oxacillinase-encoding genes, including blaOXA-23-like, blaOXA-24-like, blaOXA-51-like, blaOXA-58-like, and blaOXA-143-like).26,27 The presence of the insertion sequence ISAba1 promoter sequence in the isolates 28 and its association with the blaOXA-23-like gene 9 were examined by PCR. The primer sequences, annealing temperature, and the expected product sizes of each reaction are listed in Table 1.
Molecular characterization of genes encoding outer membrane proteins
Outer membrane proteins carO and oprD were amplified using gene-specific primers as described previously (Table 1). Sequencing was performed for strains showing a different size than expected.
Isolation and characterization of the outer membrane proteins
The outer membrane fractions of different isolates studied were prepared by N-lauryl sarcosinate method. 29 Briefly, the isolates were grown overnight in LB broth, harvested by centrifugation at 7,000 g for 10 min, then the cell pellets were sonicated, and the cell debris was discarded. The supernatants were ultracentrifuged at 100,000 g for 60 min. The pellets were incubated with N-lauroylsarcosine (final conc., 1%) at 37°C for 30 min. The outer membrane fractions were collected by ultracentrifugation at 100,000 g for 60 min. The outer membrane proteins were analyzed by SDS-PAGE with 15% polyacrylamide gels and Coomassie blue staining. 30 Outer membrane fractions were incubated in boiling water bath for 5 min before they were subjected to electrophoresis.
Statistical analysis
Each experiment was repeated thrice. The mean of the three readings was calculated and interpreted according to each experiment specification. The prevalence of resistance to each antimicrobial agent, phenotypic detection of carbapenem hydrolyzing β-lactamases, and prevalence of resistance determinants were recorded as percentage. All data were reported and analyzed using SPSS software (version 20.0; SPSS, Chicago, IL).
Results
Prevalence of A. baumannii among clinical specimens
During the period of the study, 50 isolates were identified as A. baumannii based on conventional methods and amplification of gltA gene encoding species-specific citrate synthase. Isolates were recovered from urine (14 samples, 28%), blood (21 samples, 42%), and sputum (15 samples, 30%). RAPD typing method revealed great diversity among A. baumannii isolates recovered from different clinical sources at 85% cutoff (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/mdr). Isolates U117 and U123 had a similarity coefficient more than 85% and were considered clonal. Isolates SP130 and SP134 also were considered clonal.
Antimicrobial susceptibility of A. baumannii isolates
Evaluation of antibiotic susceptibility pattern indicated that 98% were resistant to imipenem and meropenem. The proportions of resistance to other β-lactam antibiotics were distributed as follows: ampicillin/sulbactam (76%R, 20%I), piperacillin/tazobactam (96%R, 2%I), cefoxitin (100%R), cefotaxime (98%R, 2%I), ceftazidime (98%R), cefepime (100%R), and aztreonam (100%R). Resistance to other classes of antibiotics was distributed as follows: gentamicin (90%R, 2%I), amikacin (86%R, 6%I), trimethoprim/sulfamethoxazole (92%R, 8%I), ciprofloxacin (88%R, 4%I), and levofloxacin (54%R, 16%I). All isolates were sensitive to polymyxin B and colistin.
Phenotypic detection of carbapenem-hydrolyzing β-lactamases
Carbapenemase production was detected in 47 isolates (94%) using modified Hodge test, while MBL production was detected in 49 isolates (98%) based on IMP-EDTA synergy test.
Molecular characterization of carbapenem-hydrolyzing β-lactamase-encoding genes
Multiplex PCR for oxacillinase-encoding genes revealed that 47 (94%) isolates had blaOXA-23-like and blaOXA-51-like, whereas 3 samples (6%) exclusively amplified blaOXA-51-like. The genes blaOXA-24-like, blaOXA-58-like, and blaOXA-143-like were not detected. All the isolates carrying blaOXA-23-like gene also carried ISAbA1 promoter sequence, which was located upstream of the blaOXA-23-like gene (Fig. 1). Sequencing results indicated that ISAbA1 is located 25 bp upstream of blaOXA-23-like gene. The obtained sequence was deposited in DDBJ/GenBank/EMBL, with accession number LC369767.

Detection of carbapenemase-encoding genes and associated genetic elements by PCR. M. Molecular weight marker. 1: blaOXA-51-like (353 bp); 2: blaOXA-23-like (501 bp) and blaOXA-51-like (353 bp); 3: ISAba1 (548 bp); 4: ISAba1+blaOXA-23-like (875 bp); 5: blaKPC (538 bp); 6: blaGES (846 bp); 7: blaIMP (448 bp); 8: blaVIM (780 bp); 9: blaNDM (624 bp); 10: blaSIM (569 bp).
The prevalence of MBL-encoding genes among A. baumannii isolates was determined (Fig. 1). Among the tested genes, blaSIM was the most prevalent gene as it was detected in 15 strains (30%). Thirteen strains (26%), 9 strains (18%), and 3 strains (6%) harbored blaNDM, blaVIM, and blaIMP, respectively. One isolate had both blaIMP and blaNDM, while one isolate carried blaIMP and blaVIM in combination. The tested MBL-encoding genes were not detected in seven isolates (14%) that showed positive phenotypic MBL production. Among the tested Class A β-lactamases, blaKPC gene was detected in 12 strains (24%), while blaGES gene was detected in 8 strains (16%). Sixteen isolates (32%) had both blaKPC and blaGES. Combinations of various natural and acquired carbapenem-hydrolyzing β-lactamases have been detected in the tested A. baumannii isolates (Supplementary Table S1).
Molecular characterization of genes encoding outer membrane proteins
Only one fragment of the expected size (about 700 bp) of carO gene was successfully amplified in all tested strains, except three strains (U10, U117, and U123), where the product size was about 1,700 bp (Fig. 2A). The outer membrane protein (OMP) profile of isolated strains was analyzed by SDS-PAGE. A 29 kDa OMP designated carO (for carbapenem resistance-associated OMP) was detected in all tested strains, except for three strains (U10, U117, and U123), where carO OMP was absent (Fig. 2C). The absence of carO resulted from the disruption of carO gene by distinct insertion element. Sequencing analysis of those strains revealed that carO gene was interrupted by ISAba1 insertion sequence (Fig. 3). The obtained sequence was deposited in DDBJ/GenBank/EMBL, with accession number LC369618. RAPD typing has indicated that strains U117 and U123 had similarity coefficient ≥85% and were considered clonal. Both strains were isolated from urine and carried the same set of carbapenem-hydrolyzing β-lactamases (pattern C, Supplementary Table S1). However, strain U10 did not belong to the same clone and carried a different set for carbapenem-hydrolyzing β-lactamases (pattern P, Supplementary Table S1).
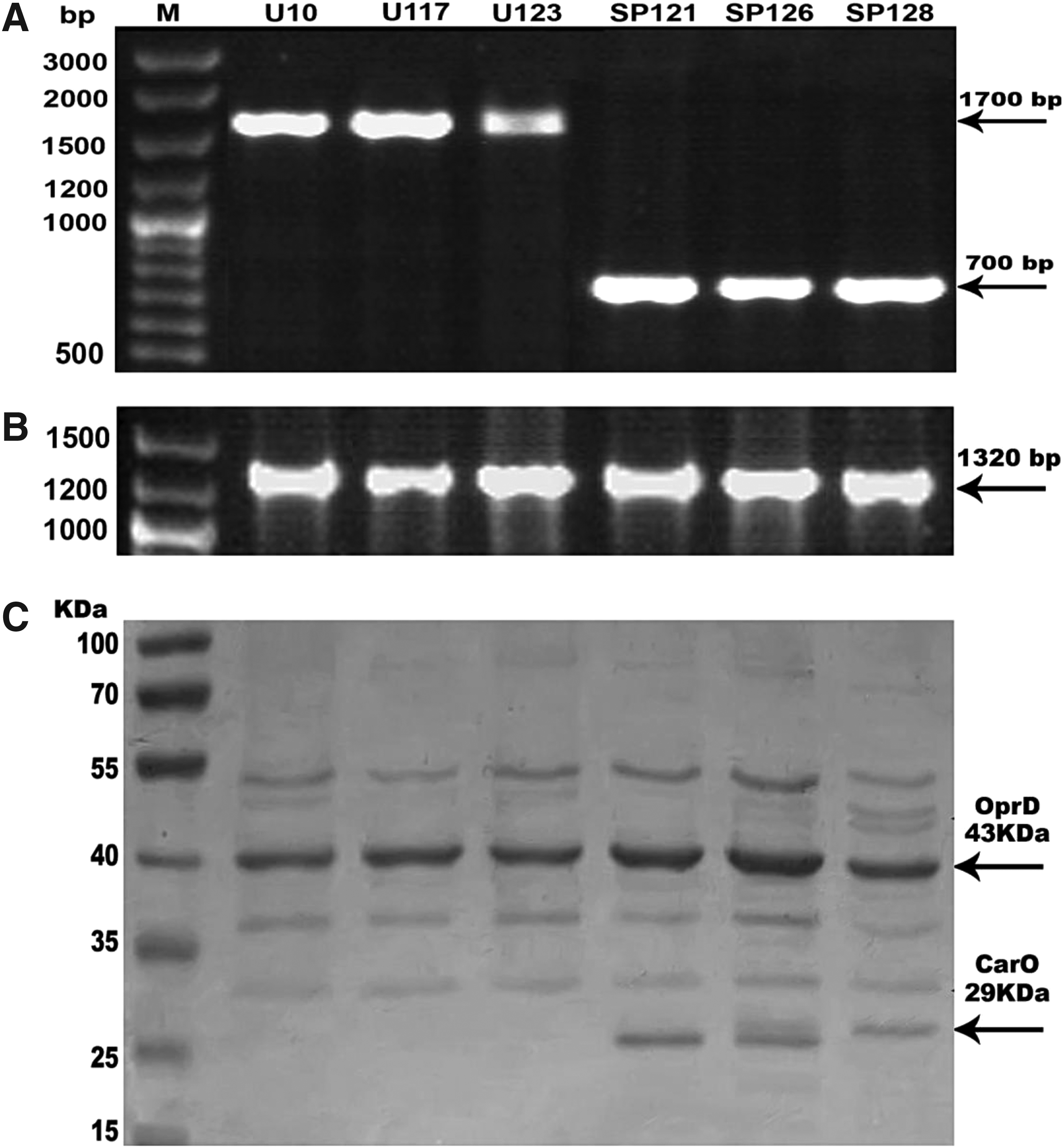
Characterization of outer membrane proteins of Acinetobacter baumannii isolates. Detection of OMP-encoding genes: carO gene

Schematic representation of carO gene in Ab_U123. The nucleotide sequence of amplified PCR product showed that carO gene was disrupted by ISAba1. carO gene and ISAba1 transposase gene (Tpase) are running on opposite strands. Gray boxes represent direct repeat (CGATGATAA), the carO gene region that was duplicated during transposition. The inverted repeats flanking ISAba1 are bold/underlined. The start and termination codons of transposase are designated bold. ISAba1 transposase promoter, minus_35_signal, is indicated in lowercase letters, while minus_10_signal is indicated in lowercase/underlined letters. The obtained complete nucleotide sequence was deposited in DDBJ/GenBank/EMBL, with accession number LC369618.
Only one fragment of the expected size (1,320 bp) of OprD gene was amplified in all tested strains (Fig. 2B). The OMP profile analyzed by SDS-PAGE revealed that oprD, a 43 kDa OMP, was detected in all strains (Fig. 2C).
Discussion
A. baumannii has emerged as a leading cause of healthcare-associated infections with eminent morbidity and mortality. 1 Carbapenems were the drugs of choice for the treatment of infections caused by β-lactam-resistant A. baumannii. 2 However, the rapid emergence of carbapenem-resistant A. baumannii is considered a substantial health problem globally due to limited therapeutic options.2,3 In this study, 98% of the isolates showed resistance to carbapenems, including imipenem and meropenem. Similar prevalence of carbapenem resistance was reported in Turkey 31 and Colombia. 32 Moreover, the percentage of carbapenem resistance has increased to 81% in Greece, 33 and to about 69% in china.34,35
Resistance to carbapenems is primarily mediated by carbapenem-hydrolyzing β-lactamases belonging to different Ambler classes. 6 Oxacillinases (Class D carbapenemases) are the most prevalent class in A. baumannii. 6 OXA-51-like β-lactamases are intrinsic to A. baumannii that usually undergoes low-level expression and confers weak hydrolytic activity to carbapenems,36,37 while OXA-23 resistance phenotype is transferable, and its production by A. baumannii strains is sufficient to confer resistance to carbapenems. 38 Among class A carbapenemases, KPC 39 and GES31,40 were detected in A. baumannii clinical isolates. Metallo-β-lactamases have also been reported in A. baumannii isolates worldwide.41–45
Coexistence of diverse intrinsic and acquired carbapenem-hydrolyzing β-lactamases has been reported in carbapenem-resistant A. baumannii. 6 A previous study has reported that all carbapenem-resistant A. baumannii clinical isolates carried OXA-23 gene, of which 31% of the isolates showed concomitant expression of NDM enzyme. 46 Huang and co-authors have reported that a large number of A. baumannii isolated from a burn center in China possessed more than one resistance gene besides the naturally occurring OXA-51. 47 In their study, OXA-23, VIM, NDM, SIM, and IMP were detected in 65.1%, 53%, 2.9%, 2.3%, and 1.1%, respectively. 47 Another report has indicated that the prevalence of OXA-23 and GES enzymes among A. baumannii clinical isolates recovered from a hospital in Los Angeles, USA, was 57.1% and 95.2%, respectively 25 ; 52.4% of the isolates harbored both OXA-23 and GES enzymes, while none of their isolates harbored KPC, IMP, VIM, SIM, or NDM. 25 The concomitant expression of OXA-23 (ranging from 52.6% to 97.5%) and the naturally occurring OXA-51 has been reported recently; however, other carbapenem-hydrolyzing β-lactamases, including MBLs, were not detected among the tested isolates.32,34,35,48 Coexistence of MBLs has been reported recently in Pseudomonas aeruginosa 49 and A. baumannii45,50,51 clinical isolates. In our study, the tested MBLs were not detected in 14% of the isolates that showed positive phenotypic MBL production. This could be due to the presence of other MBL-encoding genes like blaGIM or blaSPM. A recent study has indicated that IMP, SIM, and GIM were detected in 95.7%, 47.1%, and 42.9% of A. baumannii isolates collected from ICUs in Mansoura University Hospitals, Egypt. 45
Insertion sequences/elements play an important role in the mobilization and expression of oxacillinases in A. baumannii. 6 ISAba1, the most prevalent type, was first described in A. baumannii isolates in 2001.6,9 When ISAba1 is located 25 bp upstream of blaOXA-23 gene, it provides a strong promoter activity and magnifies its expression to a level that confers resistance to carbapenems.52,53 Our results were consistent with recent studies in which ISAba1 was detected upstream of blaOXA-23 gene in all blaOXA-23-positive isolates.32,35 The insertion sequences ISAba3 and ISAba125 are commonly associated with blaOXA-58 gene, 6 while a hybrid promoter consisting of ISAba9/ISAba10 and ISAba1 has also been reported.6,54
Several reports have indicated that OMPs have a role in resistance to carbapenems in A. baumannii.7,8 CarO participate in the influx of carbapenems in A. baumannii as it displays a pore-forming property with no binding site for imipenem, signifying a nonspecific monomeric channel function. 55 Only a single copy of carO gene was found on a single transcriptional unit in A. baumannii genome. 8 The carO gene encodes a polypeptide of 247 amino acid residues with apparent molecular weight of 29 kDa in SDS-PAGE, when OM fractions were preheated at 95°C for 5 min. 56 Limansky et al. have found that imipenem resistance was associated with the loss of CarO in A. baumannii clinical isolates in which no carbapenemase activity was detected. 29 The absence of carO in carbapenem-resistant clinical isolates of A. baumannii is a result of disruption of the gene by different insertion elements as ISAba1, ISAba10, ISAba125, and ISAba825.8,54,55 In our study, loss of carO OMP was associated with disruption of carO gene with ISAba1. A. baumannii also possesses an OprD homologue, which is known to be involved in resistance to carbapenems in P. aeruginosa. 8 However, a later study showed that oprD homologue in A. baumannii is not involved in resistance to carbapenems, but it could play a role in the adaptation of the bacterium to iron- and/or magnesium-depleted environments. 57 All tested isolates in this study harbored intact oprD gene and a 43 kDa OMP was detected in all strains.
RAPD typing method was used previously for analysis of clonality and genetic r of A. baumannii15,58,59 and other Gram-negative isolates. 60 In our study, RAPD typing method revealed great diversity among A. baumannii clinical isolates at 85% cutoff. The diversity of RAPD patterns obtained suggests dissemination of carbapenem-hydrolyzing β-lactamase-encoding genes among genetically unrelated isolates of A. baumannii. This may be attributed to horizontal gene transfer of plasmids carrying resistance determinants.
The antimicrobial susceptibility profiles of A. baumannii isolates evaluated during the period of study indicated high incidence of carbapenem resistance. Moreover, the increasing incidence of co-resistance to fluoroquinolones, trimethoprim/sulfamethoxazole, and aminoglycosides draws attention to how limited treatment options are becoming. Despite their limitations, polymyxin B and colistin could be considered the last resort for the treatment of infections caused by carbapenem-resistant A. baumannii isolates in hospital settings. However, development of resistance to these antimicrobials is a major concern. Overuse/misuse of antimicrobial agents as well as neglecting standard protocols are held responsible for the emergence and spread of multidrug-resistant pathogens in hospital settings. Therefore, implementation of effective measures is necessary to prevent any outbreaks.
Footnotes
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.