
Abstract
Increasing rate of silent intestinal carriers with extended-spectrum betalactamase (ESBL)-producing Klebsiella pneumonia (ESBL-KP) has given rise to a serious healthcare problem in clinical settings. Various epidemiological studies are being conducted to determine clonal relatedness among carriers. In this study, we investigated the intestinal carriage of ESBL-KP and clonal relatedness among ESBL-KP isolated from fecal carriage in Iran for the first time. A total of 120 rectal swabs (RSs) were collected including 61 from inpatients of intensive care unit and 59 from outpatients. ESBL-KP screening was performed using MacConkey agar supplemented with cefotaxime. PCR was done for detection of ESBL, carbapenemase, and virulence factor genes. Conjugation experiments and PCR-based replicon typing were performed. Clonal relatedness was investigated by multilocus sequence typing (MLST) and multiple locus variable number tandem repeat analysis (MLVA). Out of a total of 120 RSs, 18.3% (22/120) ESBL-KP were isolated. The rate of blaCTXM-15 was 81%. ompk35 was the most prevalent virulence gene detected in 86.3% of the isolates. In conjugation experiments, three out of five tested isolates had conjugative plasmids. The most prevalent plasmid types belonged to IncL/M, IncA/C, and Inc FII. The MLST analysis showed that the main sequence types (STs) identified among ESBL-KP isolates were ST147, ST15, and ST16. The isolates were characterized into 4 miniclusters and 11 singletons using MLVA. High heterogeneity among ESBL-KP isolates indicated that this bacterium could be colonized in different sites and easily transferred. Screening of carriers in hospitals and community could help in controlling of infection in the healthcare and community settings.
Introduction
Uncontrolled prescription of antibiotics leads to emergence of antimicrobial resistance in bacterial strains. Colonization of extended-spectrum betalactamase-producing Enterobacteriaceae (ESBL-PE) in gastrointestinal tract often has minimum clinical symptoms. Therefore, carriers are not aware of this problem. 1 Klebsiella pneumoniae as member of Enterobacteriaceae family is one of the most important pathogens causing nosocomial and community-acquired infections. 2 Highly antibiotic resistant K. pneumoniae have been increasing worldwide. 3 Moreover, increasing rate of silent intestinal carriers with ESBL-PE, specifically ESBL-producing K. pneumoniae (ESBL-KP), has given rise to a serious healthcare problem in clinical settings in recent years.4,5 Colonization with ESBL-KP in gastrointestinal tract and hands of healthcare personnel can be considered a horrible repertoire of resistant bacteria that rapidly outbreaks among patients in hospitals and also in community. Since the plasmid coding ESBL may also carry other genes including aminoglycosides, treatment is hardly effective. 6 blaCTX-M is one of the most important and prevalent ESBL genes first reported in strains of Escherichia coli isolated from patients in Germany in 1989. 7 The rate of CTX-M-15 betalactamase has recently increased all over the world. 8 Plasmids belonged to different incompatibility (Inc) groups including IncFII and IncL/M that are responsible for harboring blaCTX-M gene.9,10
Various epidemiological studies are being conducted to determine clonal relatedness among carriers. Molecular typing techniques include multilocus sequence typing (MLST) and multiple locus variable number tandem repeat analysis (MLVA), which have been used to show diversity among K. pneumoniae isolates. 11 In contrast, detection of virulence factors could be helpful in description of ESBL-KP. The major pathogenicity of K. pneumoniae is due to various virulence factors, including capsular serotype, hypermucoviscosity (magA) lipopolysaccharide, iron acquisition systems (entB), porins (ompK35 ompK36), adhesions (fimH, fimA, mrkD, mrkA, and kpn), and biofilm formation.12,13
Previous studies mostly discussed about ESBL-PE in clinical isolates. The aim of this study was to investigate the intestinal carriage of ESBL-KP and clonal relatedness among ESBL-KP isolated from fecal carriage.
Materials and Methods
Ethical statement and bacterial isolates
This cross-sectional study was carried out from June to October 2016 in a public hospital in Iran. A total of 120 rectal swabs (RSs) had been collected from patients of intensive care unit (ICU) (61) and outpatients (59). Informed consents were obtained from all participants (IR.PII.REC.1395.44). RSs were cultured as previously described. 14 In brief, the RSs were inoculated in Tryptic soy broth and then delivered to the laboratory within 2 hours. All swabs were incubated in 37°C overnight. Next day, 200 μL of bacterial suspensions was cultured on MacConkey agar supplemented with cefotaxime (CTX, 1 mg/L) and incubated at 37°C for 24 hours. K. pneumoniae ATCC 700603 was used as a positive control. Grown colonies were subcultured and identification of K. pneumoniae was confirmed by using biochemical tests. 15
Antimicrobial susceptibility test and minimal inhibitory concentration determination
The susceptibilities of CTX-resistant K. pneumoniae isolates against 11 antibiotics ceftazidime (CAZ: 30 μg), CAZ/clavulanic acid (CAZ/CLA: 30/10 μg), CTX (30 μg), CTX/CLA (30/10 μg), cefepime (CPM: 30 μg), amikacin (AK: 30 μg), gentamicin (GM: 30 μg), ciprofloxacin (CIP: 5 μg), levofloxcine (LVX: 5 μg), ertapenem (ETP: 10 μg), and imipenem (IMP: 10 μg) (MastGroup Ltd., Merseyside, United Kingdom) were tested by agar disk-diffusion method according to clinical and laboratory standards institute (CLSI) guidelines. 16 E. coli ATCC25922 was used as a control for disk diffusion method. We used the double-disk synergy test (DDST) for the phenotypic detection of ESBL producers according to the CLSI recommendation. K. pneumoniae ATCC 700603 and E. coli ATCC25922 were used as positive and negative controls in the DDST method, respectively. Minimal inhibitory concentrations (MICs) of CTX, IMP, and colistin were determined by the broth microdilution method and gradient test strips, respectively (Liofilchem, Roseto degli Abruzzi, Italy). MIC of CTX was performed in all 22 ESBL-KP isolates; however, MIC of IMP and colistin was carried out only in ETP-resistant isolates. E. coli ATCC 25922 was used as the control strain in susceptibility testing.
Biofilm formation using wrinkled colony development
Biofilm formation was evaluated as previously described. 17 We categorized wrinkled colonies into four morphology types (A, B, C, and D). The classification was done according to the background of the colonies in the presence of light, the ground state of the bacteria involved in the molecular matrix, and the thickness of the surrounding colonies.
Molecular detection of ESBL and carbapenemase genes and virulence factors
Genomic DNA was extracted by the DNA extraction kit (Bioneer Company, Korea). PCR assays were carried out using primers specific for ESBL and carbapenemase genes, including blaTEM, blaSHV, blaCTX-M15, blaVEB, blaPER, blaNDM-1, blaOXA-48, blaKPC, blaVIM, blaIMP, and blaGES 18 and virulence factors including adhesions (fimH, mrkD, mrkA, fimA, and ECP), FimH-like-adhesion (kpn), mucoviscosity-associated gene A (magA), enterobactin (entB), outer membrane protein (ompk35 and ompk36) and capsule serotype K1 and K2 as previously described.12,13,19
PCR-based replicon typing and conjugation experiments
The plasmid incompatibility group was determined by PCR-based replicon typing. 20 The conjugation experiments were carried out using the filter mating for only five selected isolates. An E. coli K12 [F− lac + Nal (r)] was considered as the recipient strain as previously described. 21 The transconjugants were selected on MacConkey agar plates containing 32 μg/mL nalidixic acid and 1 μg/mL CAZ. Plasmid DNA was extracted using the Gene All Exprep Plasmid (GeneAll Biotechnology, Seoul, Korea). The confirmation of putative transconjugants was performed by detection of blaCTX-M15 gene using PCR.
Multilocus sequence typing
A total of 22 ESBL-KP isolates were analyzed by MLST. Allele typing of seven housekeeping genes (gapA, infB, mdh, pgi, phoE, rpoB, and tonB) was done to access Pasture's MLST scheme. The list of primers used for MLST is provided on the Pasteur Institute website. The allelic profiles were compared in the Pasteur Institute MLST and whole genome MLST database to recognize the sequence type (ST) of each strain.
Multiple locus variable number tandem repeat analysis
MLVA of 22 ESBL-KP isolates was performed by PCR amplification using six variable number tandem repeat (VNTR) loci (A, D, E, H, J, and K). The PCR conditions and the list of primers were described previously. 22 Clustering of MLVA types was analyzed by GelCompar II software version 4.0 (AppliedMaths, Sint-Matens-latem, Belgium). Clustering was determined by the UPGMA with 80% similarity cutoff. Moreover, the diversity indices of the VNTR loci were measured by Simpson's Diversity Index.
Statistical analyses
Statistical analyses were performed by Fisher's exact test with SPSS v25 software to compare variables of different groups. In all experiments, p-values <0.05 were considered as statistically significant. Difference in the prevalence of ESBL among both groups (ICU patients vs. outpatients) was tested by Z test. Results were considered significant at 0.05 levels.
Results
Detection of ESBL-KP in RSs
From a total of 120 RSs collected from outpatients (59) and ICU patients (61), 10.1% (6/59) of outpatients and 26.2% (16/61) of inpatients of ICU were colonized with ESBL-KP isolates. The prevalence of ESBL was significantly higher among ICU patients than among outpatients (26.2% vs. 10.1%, p = 0.0225).
Antimicrobial susceptibility pattern
DDST showed that all the CTX-resistant K. pneumoniae isolates had ESBL positive phenotype. Disk diffusion method revealed that the resistance rates of ESBL-KP to CAZ and CPM were 90% (20/22) and 77% (17/22), respectively. Resistance to other antimicrobial agents was as follows: ETP, 41% (9/22); IMP, 41% (9/22); AK, 36% (8/22); GM, 41% (9/22); CIP, 50% (11/22); and LVX, 50% (11/22). The MICs of CTX, IMP, and colistin agents are given in Table 1. Although all collected isolates of ICU patients were resistant to cephalosporins, fluoroquinolones, aminoglycoside, and carbapenems, the isolates collected from outpatients were found to be resistant just to cephalosporine antibiotics (Table 1).
Clinical, Phenotypic, and Genotypic Characteristics of the 22 Extended-Spectrum Betalactamase-Producing Klebsiella pneumoniae Isolated from Intestinal Carriage in Iran
AK, amikacin; CAZ, ceftazidime; CIP, ciprofloxacin; CPM, cefepime; CTX, cefotaxime; E-ICU, emergency intensive care unit; ETP, ertapenem; G-ICU, general intensive care unit; GM, gentamicin; ICU, intensive care unit; IMP, imipenem; LVX, levofloxacine; MICs, minimal inhibitory concentrations; ND, not determined; OP, outpatient; UT, untypable.
Wrinkled colony morphology typing
The wrinkled colony morphology of isolates was categorized into four types (A, B, C, and D). The rates of types A, B, C, and D were 27% (6/22), 9% (2/22), 13% (3/22), and 50% (11/22) of isolates, respectively.
Detection of ESBL, carbapenemase, and virulence genes among ESBL-KP
All 22 ESBL-KP isolates harbored at least 1 ESBL gene as shown in Table 1. The blaCTX-M15, blaSHV, and blaTEM genes were present in 81% (18/22), 68% (15/22), and 63% (14/22) of isolates, respectively. The blaNDM-1, blaOXA-48, blaVIM, and blaGES genes were detected in 40% (9/22), 18% (4/22), 18% (4/22), and 4% (1/22) of isolates, respectively. None of the isolates had blaKPC, blaIMP, blaVEB, and blaPER genes. ompk35 was the most prevalent virulence gene found in 86.3% (19/22) of isolates. The presence rates of other virulence genes were as follows: ompk 36 77% (17/22), entB 72% (16/22), mrkD 72% (16/22), mrkA 72% (16/22), fimH 68% (15/22), kpn 68% (15/22), fimA 36% (8/22), ECP 36% (8/22), and K2 9% (2/22), respectively. None of the isolates had K1 and mag genes. Moreover, two isolates from outpatients had no virulence factors tested in this study (Table 1). However, our findings show that the distribution of virulence factor has no relatedness in outpatients and ICU patients.
Conjugation experiments and replicon typing
In conjugation experiments, the plasmids carrying blaCTX-M15 were successfully transferred to E. coli K12 in three of five tested isolates. PCR amplification confirmed presence of the blaCTX-M15 gene in the transconjugants. The E. coli K12 transconjugant of CTX-M15 displayed resistance to CTX (MICs >4 mg/L), but maintained susceptibility to ETP, IMP, GM, AK, and CIP. Plasmid analysis revealed that the most frequent types detected among plasmids belonged to IncL/M, IncA/C, and Inc FII incompatibility groups.
MLST analysis
The MLST analysis showed that 22 isolates were assigned to 16 different STs (Fig. 1). The number of allele differences between these STs was displayed in a minimum spanning tree (Fig. 1). The main STs identified among ESBL-KP isolates were ST147 (n = 3), ST15 (n = 3), and ST16 (n = 2). Majority of them were being associated with blaCTX-M15, blaSHV, and blaTEM as well as blaNDM-1 (Table 1). All these main STs belonged to ESBL-KP from ICU patients isolated at different periods of time, which may demonstrate the long-term sustainability of these strains in the hospital environment.

Genetic relationship of ESBL-KP. Minimum-spanning tree (MST) illustrating STs of ESBL-KP in fecal carriage isolates. STs belonging to outpatient, general intensive care unit, and emergency intensive care unit are marked in dark gray (smoked), gray and black circles, respectively. The number above the line represents the number of allele differences between these STs.
MLVA
Analysis of six VNTR loci profiles showed that the number of alleles ranged from four for VNTR_K to eight for VNTR_J (Table 2). The lowest diversity index was seen in VNTR_K (0.591) and the highest diversity index was found for VNTR_J (0.793). Based on a cutoff of 80% similarity, the isolates were characterized into 4 miniclusters and 11 singletons (Fig. 2). Some of ESBL-KP isolates isolated from outpatients and ICU patients had exactly the same MLVA profiles (K67 and K5, K6, and K64), which could indicate a widespread dissemination of these strains in the community and hospital (Fig. 2).
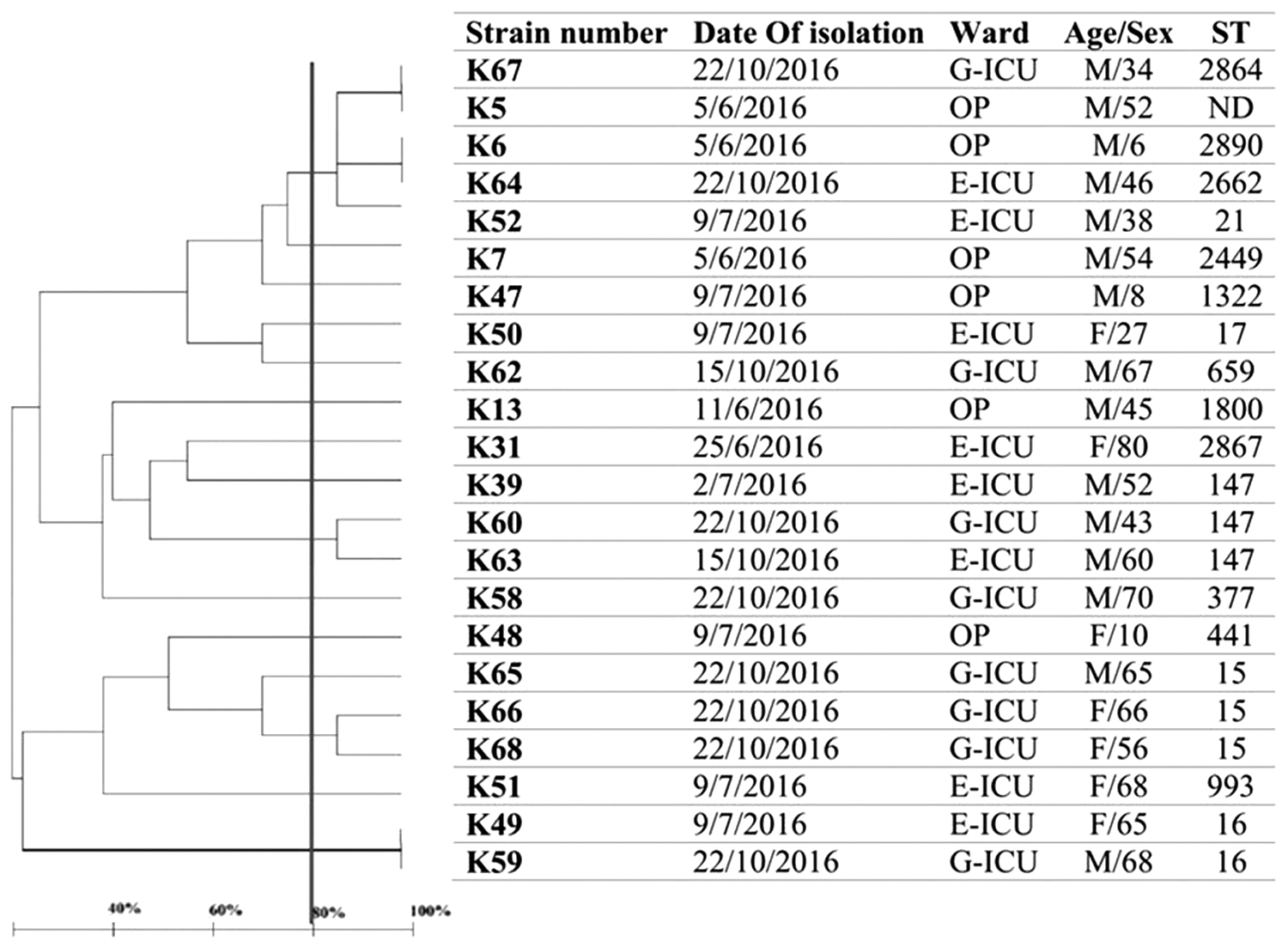
Dendrogram based on MLVA of 22 ESBL-KP in fecal carriage isolates and their STs determined through MLST. E-ICU, emergency intensive care unit; F, female; G-ICU, general intensive care unit; ICU, intensive care unit; M, male; MLST, multilocus sequence typing; MLVA, multiple locus variable number tandem repeat analysis; ND, not determined; OP, outpatient.
Diversity Indices and Confidence Interval at Each Multiple Locus Variable Number Tandem Repeat Analysis Locus
ESBL, extended-spectrum betalactamase; K, number of different repeats present at this locus; VNTR, variable number tandem repeat.
Discussion
K. pneumoniae belongs to the group of “ESKAPE” that is an important causative agent of both hospital- and community-acquired infections.23,24 Therefore, dissemination of resistant K. pneumoniae from hospital to the community is a major public health crisis. 25 Previous and prolonged hospitalizations in clinical settings and ICU as well as using some medical devices, for instance catheters, have been identified as risk factors in inpatient carriers.6,26 In outpatients, particularly in family members and healthcare workers, some risk factors including hand contact, sharing food, or even embracing 27 and frequent or inadequate use of antimicrobial agents in patients could lead to increase in the number of carriers in the population. 28 It may be suggested to screen patients at their admission since preventing the dissemination of ESBL-producing isolates to healthcare settings is notably important. 29
Previous studies mostly discussed about ESBL-PE in clinical isolates. However, investigation about fecal carriage particularly in both inpatients of ICU and outpatients are being less discussed. In Iran, several studies showed high rates of ESBL isolates in clinical settings,6,30,31 but there is no report about fecal carriage of ESBL-PE isolates especially in outpatients. We aimed to investigate the isolation of ESBL-KP among fecal carriage, which is the first study in Iran. Our ESBL-KP isolates were obtained from inpatients (6/22) and hospitalized ICU patients (16/22). The patients were characterized as noninfected inpatients and ICU patients, aged 6–80 years old, with no diarrhea or other gastrointestinal symptoms. From a total of 120 individuals, 22 (18.3%) were ESBL-KP positive, of which 10.1% (6/59) and 26.2% (16/61) were from outpatients and ICU patients, respectively. In terms of hospitalized ICU patients, our findings are similar to reports from other countries, including ESBL-KP positive isolates in India 28.3% (17/60) 32 and Tunisia 21% (5/38). 33
The profile distributions of virulence factors are given in Table 1. Within 22 ESBL-KP isolates, ompk35 and ompk36 were the most prevalent virulence genes that were detected in 86.3% (19/22) and 77% (17/22) of isolates, respectively. Many studies discussed that deletion of these outer membrane proteins may reduce the virulence and potential of pathogenicity of Klebsiella. 12 OmpK35 plays a major role in K. pneumoniae virulence and resistance to phagocytosis. 12 Since our data showed that there was significant relatedness between the presence of ompk35 and isolates collected from ICU (p < 0.05), it could be concluded that probably ESBL-KP isolates isolated from ICU patients had more pathogenicity than those isolated from outpatients. The assessment of fecal carriage rate of serotype K1 K. pneumoniae is investigated in several studies34–36 due to the fact that K1 capsule serotype is a decisive virulence factor in liver abscess. mag also plays a significant role in the pathogenesis of liver abscesses and is restricted to serotype K1. 13 Our data showed that none of our isolates had serotype K1 and mag. This could result in reducing the probability of causing liver abscess. In contrast, most of our isolates had high distribution of virulence factors along with resistance genes (Table 1). This would be so impressive because dissemination of these virulent and resistant strains to other wards could reduce the ability of elimination of them and increase the rate of carriage mode. As given in Table 1, no relationship was seen in the distribution of virulence factors and resistance genes between outpatients and ICU patients. We had two strains, K5 and K6, with no virulence factors. These isolates also had low level of resistance to CTX (MICs of 8 and 16 μg/mL, respectively) and had no carbapenemase genes. This may suggest that some outpatients colonized with less virulent or resistant ESBL-KP, but the rate can be increased if the dissemination remains uncontrolled among different environments.
In terms of biofilm formation, type D was the most prevalent detected in 50% (11/22) of isolates. Two major fimbriae known as type 1 and type 3 fimbriae are associated with biofilm formation. FimH is one of the components of type 1 fimbria. Type 3 fimbria is composed of major subunit MrkA and the adhesive unit, MrkD. 20 Our data showed that there was significant correlation between presence of these three genes, which are involved in biofilm formation, and type D of biofilms (p < 0.05).
Molecular typing techniques are precise methods for epidemiological study of bacterial infections. Taken together, our data of typing methods including MLVA and MLST showed high heterogeneity among ESBL-KP isolates. In our study, the related strains had different STs and only eight strains had the same STs (ST147, ST15, and ST16); ST147 as a successful international clone is important in the dissemination of multidrug and even extensive drug-resistance isolates globally. 36 ST15 is also widely associated with K. pneumoniae-producing CTX-M-15 in different parts of the world.37–39 Despite high heterogeneity among our isolates, analyses of phenotypic and genotyping data revealed remarkable points. MLVA dendrogram showed identical strains without any other common characteristics, including K5 (outpatient) and K67 (ICU patient). Some isolates with the same profiles also had the same distribution of ESBL genes and mostly the same types of plasmids, although they were isolated from different environments (Table 1). For instance, K64 and K6 were isolated from a 45-year-old man hospitalized at emergency-ICU (E-ICU) and a 6-year-old outpatient child, respectively. This indicated that ESBL-KP with the same profiles could be colonized in different age groups and different places and, therefore, could be disseminated widely in both community and hospital settings.
This study also revealed that the isolates with the same STs were isolated mostly from different wards in different times (Fig. 2). This may suggest that strains with the same STs could be disseminated widely, threatening the ability to control infection in healthcare settings. Furthermore, these isolates had a high distribution of ESBL and carbapenemase genes and plasmid types responsible for harboring blaCTX-M15 (FII and L/M). One of our isolates with ST147 collected from E-ICU was resistant to all tested antibiotics except colistin with high MIC for CTX and IMP; moreover, it harbored blaCTX-M15, blaSHV, blaTEM, blaNDM-1, and blaVIM. These data showed that not only in clinical samples37,40 but also in fecal carriage, ST15 and ST147 are important STs in ESBL and carbapenem-resistant isolates.
In contrast, comparison between MLVA and MLST data of ICU patients indicated that the isolates with the similar MLVA profiles had the same STs (Fig. 2). So that K60 and K63 (ST147) clustered in one MLVA group, K66 and K68 (ST15) clustered in another one, and K59 and K49 (ST16) had exactly the same MLVA type. The strains clustered in one group also had similar distribution of resistance genes and virulence factors. Therefore, it can be said that similar strains were disseminated between different wards, leading to increase in the rate of fecal carriage.
We found out that most plasmid types belonged to IncL/M, Inc FII, and IncA/C incompatibility groups. Different types of plasmid including IncFII, IncA/C, IncL/M, or untypable are able to carry blaNDM gene.41,42 Interestingly, IncFII and IncL/M plasmids have also been associated with dissemination of CTX-M-15 [9, 10]. In this study, three isolates (K65, K66, and K68), having conjugative plasmids, contained ESBL (blaTEM, blaSHV, blaCTX-M15) and carbapenemase (blaNDM-1 and blaOXA-48) genes simultaneously. Of these three isolates, two had IncL/M plasmid type and one was untypable. These isolates also had the same sequence type (ST15). The ability of conjugation and transmission of IncA/C, IncF, and IncL/M plasmids especially in isolates with ST15 may be responsible for dissemination of K. pneumoniae-producing CTX-M-15, particularly in fecal carriage. This may result in dissemination of ESBL and carbapenemase genes in different parts of hospital and community settings.
The rate of coresistance of cephalosporins and fluoroquinolones within isolates was 50% (11/22) in our study (Table 1). Recent studies showed that previous use of fluoroquinolone could be a risk factor for the acquisition of ESBL-producing isolates. ESBL genes and qnr are located in the same plasmids. This suggested that CIP resistance is associated with the presence of CTX-M beta-lactamase. 43 Our data showed that there was significant relatedness between the rate of resistance to CIP and ICU isolates (p < 0.05). Moreover, our data showed that the rate of resistance to carbapenems is also high. Therefore, it seems that finding other antimicrobial resistance agents including resistance to fluoroquinolones and carbapenems among ESBL-KP carriers is an important point.
In this study, we were not able to discuss the prevalence of fecal carriage since the sampling was done only in one public hospital. High heterogeneity among ESBL-KP isolates both in outpatient and inpatient individuals indicated that this bacterium can colonize in different sites and can easily transfer. In contrast, gut is an important ecosystem with high number of different bacterial genes, and colonization of ESBL-PE, for example, ESBL-KP, could result in later intestinal infections. 44 Titelman et al.'s findings revealed that fecal carriage of extended-spectrum β-lactamase (ESBL)-producing Enterobacteriaceae often persists up to 1 year after infection. 45 This prolonged duration of colonization can increase the risk of dissemination. Therefore, colonization of ESBL-KP in gut and hands of patients, family members, and healthcare workers or even in equipment of ICUs and invasive therapy tools (e.g., catheter) could be a silent reservoir and be responsible for future epidemic infections. Screening of carriers in hospitals, strict personal protective equipment guidelines for hospital personnel, and accurate training of family members can prevent the spread of resistance genes in bacteria and may help control infection in the healthcare settings.
Footnotes
Acknowledgments
The authors thank the personnel in the bacteriology department of Pasteur Institute of Iran, Dr. Azadeh Safarchi from the Pasteur Institute of Iran, and Loghman Hospital for their help. This work was funded by a research grant from Pasteur Institute of Iran (project no: B-9216).
Ethical Statement
This project was done based on ethical guidelines as previously approved by the Pasteur Institute of Iran (project no: 180 IR.PII.REC.1395.44).
Disclosure Statement
No competing financial interests exist.