
Abstract
Benzalkonium chloride (BAC) is widely used as a disinfectant and preservative. This study investigated the effect on antimicrobial susceptibility and the cellular changes that occurred after exposure of Klebsiella pneumoniae clinical isolates to sublethal concentrations of BAC. Minimum inhibitory concentration and minimum bactericidal concentration of BAC were determined for the collected 50 K. pneumoniae clinical isolates by broth microdilution method, and the tested isolates were adapted to increasing sublethal concentrations of BAC. The effect of adaptation on MICs of the tested 16 antimicrobial agents, the cell ultrastructure, efflux, and membrane depolarization of the tested isolates were examined. Interestingly, most K. pneumoniae isolates that adapted to BAC showed increased antimicrobial resistance, various morphological and structural changes, increased membrane depolarization, and enhanced efflux activity. The findings of this study suggest that the extensive use of BAC at sublethal concentrations could contribute to the emergence of antibiotic resistance in K. pneumoniae clinical isolates that might complicate the therapy of infections caused by this pathogen. In conclusion, the hazard associated with the prolonged exposure to sublethal concentrations of BAC represents a public health risk and therefore it should be a focus in both hospital and community sanitation practices.
Introduction
T
Bacteria are continuously challenged by a gradient of biocide concentration from a subinhibitory level up to an inhibitory level in various settings including domestic, health care, and industrial environments. 5 Quaternary ammonium compounds like benzalkonium chloride (BAC) are common biocides that are widely used as disinfectants in food-processing and medical environments and as preservatives in pharmaceuticals and cosmetics. 6 There is an increasing concern that the use of biocides, such as BAC in these environments may contribute to the development of microorganisms with increased resistance to antimicrobials. 7 Antimicrobial resistance is a global public health threat that involves all major microbial pathogens and antimicrobial drugs. The factors involved in the increase of antimicrobial resistance should be studied worldwide. 8 A principal aim of this study is to determine the effect of exposure of K. pneumoniae isolates to sublethal concentrations of BAC on antimicrobial susceptibility, bacterial ultrastructure, membrane depolarization, and efflux system.
Materials and Methods
Bacterial isolates
A total of 50 K. pneumoniae isolates were collected from various departments of Tanta University Hospital from January to July 2017. Isolates were obtained from urine (11 isolates; 22%), wound (13 isolates; 26%), blood (7 isolates; 14%), and sputum (19 isolates; 38%). The clinical isolates were examined microscopically and were identified using standard biochemical tests according to the protocols suggested by MacFaddin 9 and included the oxidase test; glucose and lactose fermentation, gas and hydrogen sulfide production in triple sugar iron agar; motility and indole production in indole motility medium; citrate and ornithine decarboxylation; urease production; and methyl red tests. Klebsiella pneumoniae ATCC 13883 was used as reference strain.
Chemicals
Chemicals utilized in this study were bis-(1,3-dibutylbarbituric acid) trimethineoxonol [DiBAC4(3)] (Invitrogen), potassium phosphate dibasic anhydrous and potassium dihydrogen phosphate (Merck), and ethidium bromide (EtBr) (Himedia). All other chemicals utilized in this study were of analytical grade, purchased from Sigma-Aldrich.
Determination of minimum inhibitory concentration and minimum bactericidal concentration of BAC
BAC minimum inhibitory concentration (MIC) for each tested bacterial isolates was determined by broth microdilution method in Mueller–Hinton broth (MHB) (Oxoid) using microtitration plates according to Clinical and Laboratory Standards Institute (CLSI) guidelines. 10 Each plate contained a growth control well (positive control) and uninoculated well (negative control). MIC is the lowest concentration that completely inhibits growth of bacteria and it was indicated by no turbidity in the wells compared to the positive and negative control. After MIC determination in MHB, the wells showing growth inhibition were subcultured on Mueller–Hinton agar (MHA) (Oxoid) for determination of minimum bactericidal concentration (MBC). The MBC was defined as the lowest bactericidal concentration of BAC required to kill bacteria after incubation at 37°C for 24 hours. All determinations were carried out in independent triplicates. As described previously, 6 isolates showing MIC >12 μg/mL were considered BAC tolerant isolates.
Adaptation to BAC
Adaptive responses of bacterial isolates were investigated by exposing the tested isolates daily to an increasing sublethal concentrations of BAC according to the method described before 11 with a starting concentration of 0.5× MBC of BAC. When growth was observed, a 10-fold diluted culture was transferred to fresh MHB supplemented with a higher concentration of BAC. This step was continued until no growth was observed after incubation for 24 hours at 37°C. Then the suspension from the last tube that showed bacterial growth were spread with a loop on MHA and incubated for 24 hours at 37°C for growth conformation and storage. All isolates were investigated in triplicate.
Determination of antimicrobial MICs
K. pneumoniae isolates were screened for antimicrobial susceptibility, before and after adaptation to BAC, to a panel of 16 antimicrobial agents on MHA by agar dilution method as described by the CLSI. 10 The following antimicrobials were tested: ampicillin, ampicillin/sulbactam, cefazoline, cefaclor, cefotaxime, cefepime, erythromycin, azithromycin, amikacin, gentamycin, tetracycline, chloramphenicol, ciprofloxacin, lomefloxacin, sulfamethoxazole/trimethprime, and imipenem.
Electron microscopic examination
The tested isolates before and after adaptation had been subjected to electron microscopic examination. The procedures were carried out according to McDowell and Trump 12 in the electron microscope unit, Mansoura University, Egypt using scanning electron microscope (SEM) (Akashi Seisakusho, Japan) and transmission electron microscope (TEM) (Jeol-1200 ECII, Japan).
Membrane depolarization assay
Membrane depolarization was measured in the tested isolates before and after exposure to sublethal concentrations of BAC using the molecular probe DiBAC4(3). 13 This fluorescent agent exhibits enhanced fluorescence and a red spectral shift as it can enter depolarized cells and binds to intracellular proteins. The tested isolates before and after adaptation were harvested by centrifugation at 12,000 rpm and resuspended in phosphate buffered saline. Then, the cells were stained with 5 μg/mL DiBAC4(3) and cell staining was analyzed using FACS verse flow cytometer (BD Biosciences, USA).
Evaluation of efflux activity before and after adaptation
The assessment of efflux of EtBr as a model substrate was conducted in the tested isolates before and after adaptation to BAC using the semi-automated fluorometric cartwheel method as described by Suresh et al. 14 Briefly, tryptic soy agar containing EtBr concentrations ranging from 0 to 2.5 mg/L were prepared and protected from light. The plates were then divided into sectors by radial lines. Cultures were then swabbed on EtBr agar plates starting from the plate center toward its edges and incubated at 37°C for 18 hours in the dark. The cultures were placed on an 1800 UV-Vis trans-illuminator (SCHIMADZU, Japan) and photographed. The minimum concentration of EtBr that produced fluorescence of the bacterial isolates was recorded, taking the reference strain as negative control.
Quantitative Real Time-PCR
The expression of four efflux genes (acrB, mdfA, norE, and yihV) representing the four major families of multidrug-resistant efflux pumps in Gram-negative bacteria were detected by Quantitative Real Time (RT)-PCR (qRT-PCR) and gapA gene was used as housing gene as described by Huguet et al. 15 using Rotor-Gene Q 5plex instrument (Qiagen, Germany). Total RNA was extracted from the tested isolates according to the manufacturer's instructions using the Purelink® RNA Mini Kit (Thermo SCIENTIFIC, USA). The RNA concentrations and the purity were assessed by Nanodrop ND1000 UV-Vis spectrophotometer (Thermo SCIENTIFIC). Then, RNA was retrotranscribed into complementary DNA (cDNA) using Power cDNA Synthesis Kit (first-strand cDNA synthesis) (iNtRON Biotechnology, Korea). Amplification of the selected genes was performed using specific oligonucleotides (Table 1) using Power SYBR® Green master mix (Thermo SCIENTIFIC). RNA samples amplified without reverse transcription were used as negative control to detect possible DNA contamination. Relative gene expression was determined using the 2−ΔΔCt method. 16 Isolates not exposed to biocides were used as calibrators or control samples (expression set to 1). An increase of twofold or greater compared to that of the control samples was considered as indicative of overexpression. 17 All experiments were performed in triplicate and the results were expressed as mean ± standard deviation values.
Oligonucleotides Used in Quantitative Real Time-PCR
Statistical analysis
Independent repeating of experiments (minimally three times) was performed to achieve reproducibility. Expression of data was in the form of mean and standard deviation. One-way analysis of variance was used to detect the significant differences between the groups (p < 0.05) using IBM SPSS (17.0, IBM, USA).
Results
In this study, it was found that 12% of the isolates were tolerant to BAC before adaptation. However, the percentage of tolerant isolates has increased to 26% after adaptation. Antimicrobial susceptibility testing of the tested K. pneumoniae isolates before and after adaptation revealed that a significant increase in resistance to the tested antimicrobials after adaptation had occurred (p < 0.05) except amikacin and chloramphenicol as shown in Table 2.
Prevalence of Antimicrobial Resistance in Klebsiellae pneumoniae Isolates Before and After Adaptation to Benzalkonium Chloride
BAC, benzalkonium chloride; MIC, minimum inhibitory concentration.
Determination of bacterial morphology and ultrastructure before and after adaptation to BAC
In an attempt to elucidate the reason why bacterial adaptation to BAC decreases its susceptibility to BAC and other antimicrobials, morphological and ultra-structural changes in K. pneumoniae occurring after adaptation to BAC were examined by SEM and TEM.
The SEM analysis (Fig. 1) revealed that after adaptation to BAC, considerable morphological modifications were exhibited that ranged from deformed cells, with individual bumps, grooves, ridges, and cavities, to overall cell surface wrinkling. Cell wall disruption was observed in the form of cracks, holes, or even cell lysis. Also, clusters of some lysed and fused cells were observed. Moreover, the cells appeared thinner and longer than the nonadapted cells. TEM examination of cells adapted to BAC showed electron dense regions, the cytoplasm was disorganized and spacing between inner and outer membrane occurred as shown in Figure 2 where a representative isolate K35 (showing a significant increase in MICs of BAC and other tested antimicrobials with p-value <0.05 after adaptation to BAC) was studied.

Scanning electron microscope image of representative Klebsiellae pneumoniae isolate before
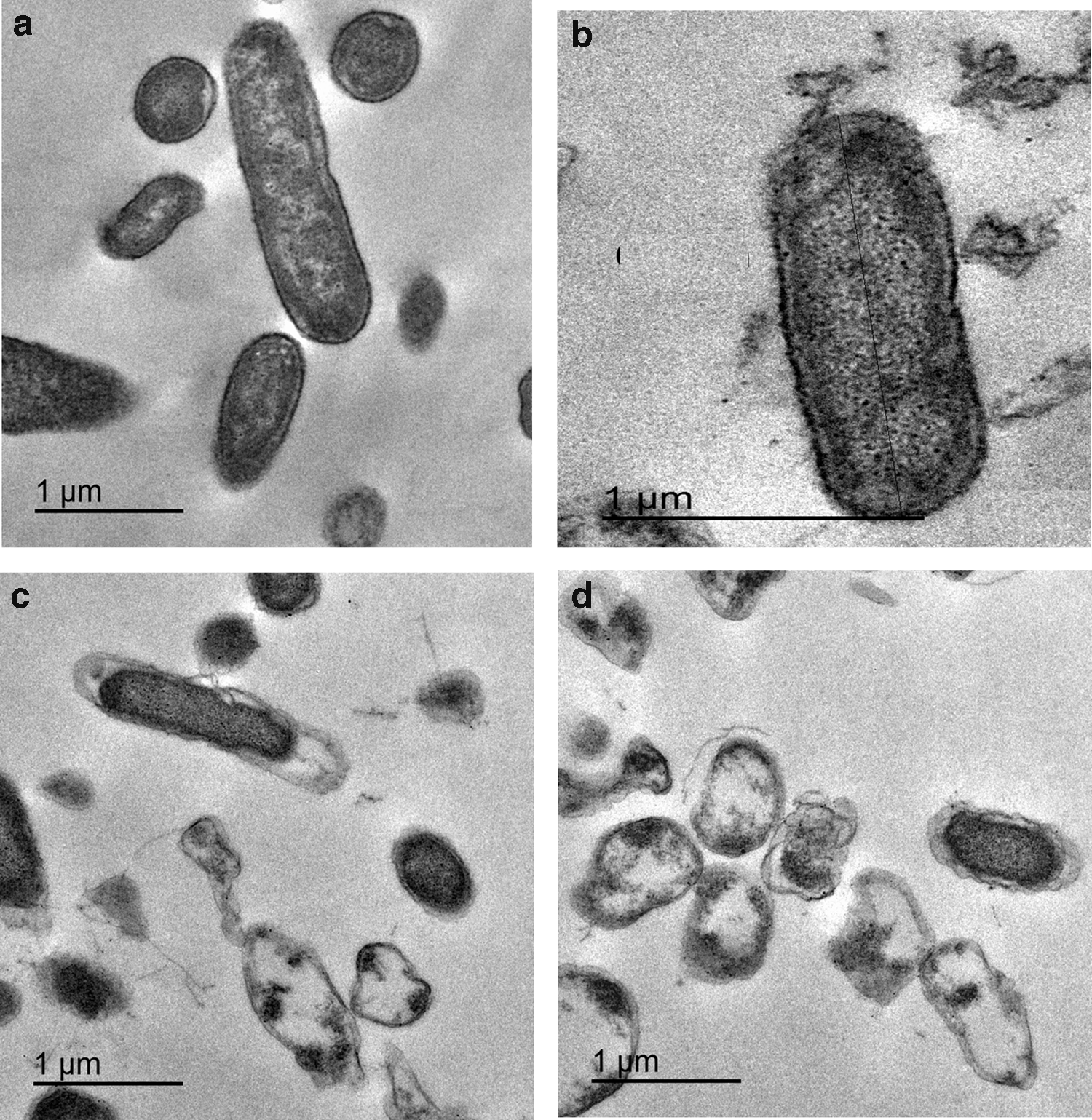
Transmission electron microscope image of representative Klebsiellae pneumoniae isolate before
Membrane depolarization
In our experiment, DiBAC4(3) entering into the cell indicated membrane depolarization by increasing fluorescence. 13 The change in membrane potential using DiBAC4(3) was investigated using flow cytometry. It was observed that 33 (66%) out of 50 isolates showed a significant increase in fluorescent gap after adaptation to BAC (p < 0.05). A representative example (isolate K35) is shown in Figure 3, where we observed that fluorescent gap has increased from 54.6% before adaptation to 96.4% after adaptation to BAC. The average increase in fluorescent gap in the tested isolates was 61% after adaptation to BAC.

Representative example of effect of BAC adaptation on membrane depolarization of Klebsiellae pneumoniae isolate before adaptation [dot plot
Detection of efflux
The EtBr-agar cartwheel method used for detection of efflux activity as EtBr traverses the bacterial cell wall of Gram-negative bacteria via porin channels and once inside, it can be concentrated to a point where it fluoresces when excited by UV light. Efflux pumps of bacteria recognize this substrate and are able to extrude it to the medium. 18 The minimum concentration of EtBr that produces fluorescence of the bacterial mass was recorded. In the current study, K. pneumoniae isolates showed increase in efflux activity after adaptation to BAC as shown in Table 3. Clinical isolates lacking efflux activity were found to fluoresce at EtBr concentration of 0.5 mg/L. Interestingly, 11 out of 20 of such isolates showed efflux activity after adaption to BAC.
Determination of Efflux Activity at Varying Concentrations of Ethidium Bromide as Flourochrome
No. of bacterial isolates.
EtBr, ethidium bromide.
Quantitative RT-PCR
With the aim of better understanding of the influence of adaptation to BAC on efflux systems, the expression of four efflux pump genes in ten K. pneumonia isolates (showing the highest increase in fluorometric efflux with EtBr in the cartwheel method after adaptation) before and after repeated exposure to sublethal concentrations of BAC was tested. We examined the gene expression of the adapted cells relative to the corresponding non adapted cells as shown in Table 4.
Relative Gene Expression (Mean ± Standard Deviation) for the Tested Klebsiellae pneumoniae Isolates After Adaptation to Benzalkonium Chloride
Bolded values point to changes in gene expression >2-fold (relative gene expression calculated using 2−ΔΔCT method).
Primer dimers and other artifacts were evaluated by melting curve analysis. To confirm that specific amplification had occurred, the melting curves of each amplicon were assessed.
Discussion
The major pathways involved in development of resistance when bacteria are subjected to subinhibitory antibiotic concentrations have been reviewed recently 19 and it is known that bacteria follow similar pathways while adapting to sublethal concentrations of biocides. 20 The objective of this work was to understand the effect of adaptation of K. pneumoniae to BAC. To achieve this aim we investigated the change occurred in antimicrobial susceptibility, ultrastructure, membrane depolarization, and efflux pump gene expression after exposure of K. pneumoniae isolates to increasing sublethal concentrations of BAC. In this study, 14% of the tested isolates acquired tolerance to BAC after exposure to the increasing sublethal concentrations. This finding is in accordance with other research articles8,21,22 where adaptation of bacterial isolates to different biocides has been found to increase biocide tolerance. Moreover, we have found that there was a significant increase in antimicrobial resistance in K. pneumoniae isolates after adaptation to BAC. This finding is in agreement with reports of other authors.7,8,23–25 There are several mechanisms used by microorganisms to escape the toxicity of antimicrobial agents, especially those by the adapted strains. Generally, the resistance against BAC is attributed to changes in fatty acid profile of cell membrane, although it does not seem to be the sole mechanism as the alterations are reported during early stages of adaptation itself. 26 Multiple drug resistance in K. pneumoniae has often been attributed to efflux pump activity that confers adaptive and cross-resistance against antimicrobials. Moreover, correlation between resistances against antimicrobials specify a common nonspecific mechanism such as change in bacterial morphology, increase in membrane depolarization and efflux pump activity that might equip microorganisms with resistances against broad spectrum antimicrobials and lead to selection of antibiotic-resistant strains. 27
As one major way in which microorganisms cope with their environment is to alter their morphology and various morphological adaptations have been reported and reviewed. 28 In our study, morphological changes of the adapted cells, resulting in changes in shape, size, and roughness of the cell surfaces, were obvious and this agrees with the research conducted by several researchers.8,18,29
Although other mechanisms may be involved,22,30 increased membrane depolarization appears to play a role in increased resistance in adapted cells. Membrane potential is the difference in electric potential between the interior and exterior of a biological cell. It is important that the cell provide power to operate a variety of molecular devices such as ion pumps embedded in the membrane and to transmit signals between various parts of a cell. 31 In this study, we determined that membrane depolarization increased after adaptation to BAC that could participate in the observed increase in antimicrobial resistance in adapted cells.
To study the effect of adaptation on efflux pumps in K. pneumoniae isolates, their activity using fluorometric analysis of the accumulation and efflux of EtBr was detected. It is known that EtBr, similar to BAC and other substances such as antibiotics, is a substrate for the efflux pumps.32,33 It is interesting that the percentage of isolates that showed fluorometric efflux with EtBr as a substrate after adaptation increased from 60% to 82%. This was further studied using qRT-PCR to detect the influence of adaptation on the expression of efflux pump genes. Our results showed that repeated exposure to sublethal doses of BAC increased the expression of acrB, mdfA, norE, and yihV efflux pump genes, and these results are consistent with Martins et al. 18 and Maseda et al. 34
In conclusion, we have showed that repeated exposure to sublethal concentrations of BAC may contribute not only to the increased tolerance to BAC but also to the promotion of cross-resistance to a range of antimicrobial agents in K. pneumoniae. This can occur either by change in bacterial morphology, increase in membrane depolarization, increase in efflux activity, or all of these factors together. The identification of the possible molecular mechanisms of biocide adaptation in K. pneumoniae should help to improve appropriate control measures to prevent further spread of these opportunistic pathogenic bacteria in both hospitals and community with frequent use of BAC. The data of this study strongly recommend that underlying hazards associated with prolonged exposure to low concentrations of a single biocide like BAC should be a focus in hospital and community sanitation. Thus, much more efforts are urgently required to minimize the use of biocides, and to develop biological systems to effectively degrade such agents from the environment.
Footnotes
Disclosure Statement
No competing financial interests exist.