
Abstract
Aspergillus clavatus is a common environmental species known to cause occupational allergic disease in grain handlers. We have recently observed azole-resistant isolates of this fungus as a cause of onychomycosis. To further characterize the cause of resistance, the genes encoding 14 α-sterol demethylase enzyme (cyp51A and cyp51B) were characterized and analyzed in 9 ITC-susceptible isolates and 6 isolates with high minimum inhibitory concentrations (MICs) of clinical (nail and sputum) and environmental A. clavatus strains. We found that six isolates with itraconazole MIC >16 mg/L demonstrated nonsynonymous mutations, including V51I, L378P, E483K, and E506G, and synonymous mutations, including F53F, A186A, Q276Q, and H359H. Moreover, P486S was detected in five strains with ITR MIC >16 mg/L. One mutation, F324S, was detected in an isolate with posaconazole MIC >16 mg/L. The effect of E483K and P486S mutations of CYP51A on azole resistance was further investigated using homology modeling and molecular dynamics. We found that E483K and P486S mutations were located near the ligand access channel of CYP51A that could partly lead to narrowing the entry of the ligand access channels. Therefore, we concluded that E483K and P486S mutations may potentially contribute to the limited access of inhibitors to the binding pocket and therefore confer resistance to azole agents.
Introduction
Aspergillus infections remain as significant causes of mortality in different immunocompromised hosts, particularly neutropenic patients with hematopoietic stem cell transplantation, solid organ transplantation, and those receiving chemotherapy.1,2 Although numerous species of Aspergillus have been implicated in different clinical features of aspergillosis, Aspergillus fumigatus, Aspergillus flavus, Aspergillus niger, Aspergillus terreus, and Aspergillus nidulans are the predominant pathogens reported worldwide.3,4 Aspergillus clavatus, a cosmopolitan fungus usually isolated from soil, dung, and grains, has been reported to be responsible for malt worker's lung, 5 tremorgenic neurotoxicosis, 6 endocarditis, 7 otomycosis, 8 and onychomycosis. 9 The production of patulin by A. clavatus, a mycotoxin with gastrointestinal, immunological, and neurological outcomes, has raised concerns about contaminated foods and fruit-based products.10,11 In general, Aspergillus species are susceptible to triazole compounds for the treatment and prophylaxis of aspergillosis, such as voriconazole, itraconazole, and posaconazole; however, resistance leading to clinical failure has been shown to be increasing.1,12,13 The most important mechanism of Aspergillus azole resistance is associated with mutation in the azole target gene cyp51A, which encodes lanosterol demethylase.14,15 To our knowledge, in vitro antifungal susceptibility data are relatively limited for A. clavatus and other members of section Clavati. 9 Also, despite global reports on the resistance of A. fumigatus and A. flavus isolates to antifungal agents specifically azoles,1,13–19 no report describing the mechanism of intrinsic or acquired resistance in this species has been published to date. Only recently, high minimum inhibitory concentrations (MICs) of A. clavatus isolated from onychomycosis to voriconazole and other lanosterol demethylase inhibitors (i.e., itraconazole, posaconazole, econazole, and miconazole) have been documented. 9 The current study shows that variations in the cyp51A and cyp51B genes of A. clavatus are associated with resistance and investigates the potential mechanism of resistance in azole-resistant A. clavatus isolates by modeling the 3D structure of these proteins and assessing the effects of mutations on the structure and possible drug access.
Materials and Methods
A total of 15 clinical, environmental, and standard isolates of A. clavatus were included on the basis of their susceptibilities to itraconazole, voriconazole, and posaconazole. The A. clavatus strains used in the study included (i) two strains, JCM 10080 and CBS 514.65 (environmental strains), used to characterize the cyp51A and cyp51B genes, as well as to act as control strains for antifungal susceptibility testing; (ii) four further environmental (air) isolates; and (iii) nine clinical A. clavatus strains isolated from patients with onychomycosis (nail) and asthma (sputum) (from the culture collection of the Invasive Fungi Research Center [IFRC]; Table 1). The strains were stored at −80°C in glycerol broth and cultured on potato dextrose agar and Sabouraud's dextrose agar 20 supplemented with chloramphenicol for 7 days at 35°C to ensure purity and good sporulation.
Antifungal Susceptibility Profile, Nucleotide and Amino Acid Substitutions in the cyp51A and cyp51B Genes from Aspergillus clavatus Clinical and Environmental Isolates
AMB, amphotericin B; BUT, butenafine; CAS, caspofungin; ITC, itraconazole; LAN, lanoconazole; LUL, luliconazole; MIC, minimum inhibitory concentration; POS, posaconazole; TER, terbinafine; TOL, tolnaftate; VRC, voriconazole.
In vitro antifungal susceptibility testing
In vitro antifungal susceptibility testing was performed using the previously described microdilution guidelines in the Clinical and Laboratory Standards Institute (CLSI) M38-A2 document. 20 Conidia from fresh colonies were scraped and suspended in sterile distilled water containing 0.05% Tween 20. The densities of the conidial supernatants were adjusted to turbidity equivalent to 80–82% transmittance at 530 nm, and after dilution in RPMI 1640 medium (1:50), the final inoculum in 96-well microtitration plates was between 2 × 10 5 and 5 × 10 5 CFU/mL. Antifungal agent concentration ranged from 0.016 to 16 μg/mL for itraconazole (Janssen, Beerse, Belgium), voriconazole (Pfizer, Sandwich, United Kingdom), amphotericin B (Bristol-Myers-Squib, Woerden, The Netherlands), tolnaftate (Sigma-Aldrich, Steinheim, Germany), and posaconazole (Merck Sharp & Dohme BV, Haarlem, The Netherlands); 0.008–8 μg/mL for caspofungin (Merck Sharp & Dohme BV); 0.004–4 mg/L for butenafine and terbinafine (Sigma-Aldrich); and 0.001–1 for luliconazole and lanoconazole (Nihon Nohyaku Co, Osaka, Japan). Drugs stock solutions were prepared in dimethyl sulfoxide. For quality control, two standard strains of Candida recommended by CLSI, Candida parapsilosis (ATCC 22019) and Candida krusei (ATCC 6258), were used. After inoculation, the microtiter trays were incubated at 35°C for 48 hr. Visual readings were used for determination of MIC, as the lowest concentration of antifungal agent that completely inhibited growth, while the minimum effective concentration (MEC) was read as the lowest concentration of drug producing compact and rounded forms compared with complete growth in the well of the control.
PCR amplification and analysis of cyp51A and cyp51B gene sequences
The strains were grown and incubated at 25°C for 7 days on Czapek agar and malt extract agar. 10 Fungal identity was reconfirmed by sequencing of the β-tubulin gene (Accession Nos. MG322299–322309) using two primers (Bt2a: 5′-GGTAACCAAATCGGTGCTGCTTTC-3′) and reverse (Bt2b: 5-ACCCTCAGTGTAGTGACCCTTGGC-3′) as described previously. 21 Total DNA of A. clavatus strains was extracted from a hyphal plug taken from a fresh colony cultured on media using previously described procedures.21,22 The coding sequences of the cyp51A and cyp51B genes were amplified for the isolates with low and high triazole MIC values using specific primer sets: Acla-A1 (5′-CTGTATGGTTTGGGATTATAGG-3′) and Acla-A2 (5′-CCTCTGACACTTAGCTTAGG-3′) for the cyp51A gene and Acla-B1 (5′-CATGGTGCCTCTGTCTTATAG-3′) and Acla-B2 (5′-TCAGAAGGTCTCAAGGGTG-3′) for the cyp51B gene. PCRs were carried out in 25 μL containing 2 μL genomic DNA, 1 μL of 25 pmol of each primer, 400 μM deoxynucleotide triphosphates, 2.5 U of Taq DNA polymerase, 2.5 mM MgCl2, and water. The following cycling steps were used for amplification of cyp51A gene: initial denaturation at 95°C for 3 min, 35 cycles of 94°C for 30 sec, 59°C for 45 sec, and 72°C for 2 min, followed by a final extension step at 72°C for 7 min; and for cyp51B gene: 95°C for 5 min and 30 cycles of 95°C for 30 sec, 61°C for 1 min, and 72°C for 2 min, finalized by an extension at 72°C for 7 min. All PCR products were sequenced from both directions using an automated DNA Sequencer (ABI PRISM™ ABI-3730 Genetic Analyzer; PE Applied Biosystem) to assess potential mutations associated with azole resistance. The sequences were assembled and edited with CLC Genomics Workbench software package (CLC Bio, Qiagen, Aarhus, Denmark), aligned with reference strains and finally compared with each other.
Homology modeling
The amino acid sequences of CYP51A from the reference 92 and 763 isolates were used for construction of conformational models of CYP51A. Protein modeling simulation and analysis were performed using ICM-pro (Molsoft). The homology modeling procedure in ICM-pro consists of the following steps: the primary protein sequences of CYP51A from the 92 and 763 isolates were imported to the software, and Protein Data Bank (PDB) searched for a similar and homologous structure. Afterward, the PDB template (crystal structure of CYP51B from A. fumigatus PDB code: 4UYM) was selected, and the query sequence was aligned against the template primary amino acid sequence. The conformational structure of the template was optimized, including deletion of water molecules, optimization of His, Pro, Asn, Gln, and Cys, optimization of hydrogens, and assignment of ideal geometry of residues. Subsequently, automated protein modeling was performed using the internal mechanics force field. This predicts side-chain torsion angles by simultaneous global optimization of energy for all nonidentical residues. 23 Protein side chains and loops were modeled by internal coordinate definition of the molecular object combined with computationally efficient biased probability Monte Carlo optimization. 24 Finally, the modeled structure was checked using the protein health option inside ICM and the side chains and loops were energy minimized and refined. The models were also evaluated using the SAVE server (https://services.mbi.ucla.edu/SAVES). The tools used for evaluation include PROCHECK, WHAT_CHECK, ERRAT, VERIFY_3D, PROVE, and Ramachandran plot.
Molecular docking
To analyze the predicted substrate and drug binding pocket of CYP51A and probe interaction of a test ligand with the amino acids in this region of the protein, itraconazole was docked to the modeled CYP51A structure using the ICM-pro docking option. To validate docking parameters, crystal structure of the energy-minimized CYP51B from A. fumigatus was redocked with voriconazole. To dock itraconazole to the modeled structure, the model was optimized by adding hydrogen atoms, defining the protonation states of residues and minimizing the structure to remove steric clashes. Itraconazole was also checked for bond types and formal charges using Merck molecular force field implemented in ICM software. The ICM score was used to rank the docking poses.
Molecular dynamics simulation
The molecular dynamics simulation was performed in an explicit solvation model on both forms of CYP51 (wild type and mutant) to understand the exact mechanism of drug resistance in the mutant form. The simulation was begun by solvation of the proteins in a cubic box with more than 3,000 TIP3P water molecules. Then, Na+ and Cl− counter ions were added to the box up to a 50 mM concentration to neutralize the system. Subsequently, systems were minimized by conjugate energy minimization until the energy gradient was 0.001. Thereafter, the systems were gradually heated from 0 to 300K in 5 ns steps and then equilibrated for another 10 ns to stabilize them. Finally, the production steps were carried out at the isothermal/isobaric ensemble (NPT) for 50 ns. The temperature and pressure were held constant at 300K and 1.01 bars by means of Langevin dynamics and Langevin piston method. Nonbonded interaction cutoff was also set to 14 Å, and the spherical cutoff method was used to treat long-range electrostatic interactions. The trajectories were saved every 2 ps and analyzed for changes in root-mean-square deviation (RMSD), root-mean-square fluctuation (RMSF), and radius of gyration (Rg) value. All the simulation steps were conducted on BIOVIA Discovery Studio 2017.
Results
In vitro susceptibility testing
The 15 strains of A. clavatus exhibited different susceptibilities to the tested antifungal agents (Table 1). Two new imidazoles, lanoconazole and luliconazole, demonstrated better activity against all A. clavatus isolates, in comparison with the other tested agents. MIC values of luliconazole and lanoconazole ranged from 0.001 to 0.125 mg/L and from 0.002 to 0.0625 mg/L, respectively, compared with MEC range 0.016–2 mg/L for caspofungin and MIC ranges 0.032 to >16 for amphotericin B, 0.25–2 mg/L for voriconazole, 0.125 to >16 mg/L for posaconazole, 0.125 to >16 mg/L for tolnaftate, 0.125 to >4 mg/L for butenafine, and 2 to >4 mg/L for terbinafine. The itraconazole and voriconazole MIC values for standard strain, CBS 514.65, were 1 mg/L, while these values for strain JCM 10080 were 1 and 0.5 mg/L. Because azole breakpoints have not been established yet for A. clavatus strains, MIC >16 mg/L has been considered high MICs for itraconazole. Three clinical isolates, 91 (Accession No. MG322306), 92 (Accession No. MG322307), and 94 (Accession No. MG322308), and one environmental isolate, 59 (Accession No. MG322299), with ITC MIC value of ≤0.5 mg/L were defined as ITC susceptible. Six strains, 877 (Accession No. KP851942), 87 (Accession No. 322303), 1193 (Accession No. 322301), 95 (Accession No. 322303), 763 (Accession No. MG970259), and 96 (Accession No. 322309) exhibited ITC MICs >16 mg/L, which were considered strains with high MICs for ITC. Among all clinical isolates, only one patient previously received azole agent. Moreover, regarding the proposed MIC breakpoints for resistance to posaconazole in A. fumigatus strains (MIC >0.25 mg/L), 25 14 A. clavatus isolates had approximately high MICs to posaconazole with MIC values ≥1 mg/L (Table 1). The isolate 91 (Accession No. 322306) demonstrated completely high MIC values to posaconazole with MIC value of >16 mg/L. Furthermore, the majority of isolates demonstrated VOR MIC value of ≤1 mg/L except for the isolate 1194 (Accession No. 322308) (MIC value 2 mg/L) (Table 1).
Characterization of cyp51A and cyp51B genes
The coding sequences of cyp51A and cyp51B genes of all A. clavatus strains were successfully amplified and the sequences were submitted to the GenBank and registered as accession numbers (MF987528–MF987557). The protein coding region of cyp51A gene is composed of 1,626 bp with a single intron of 81 bp, whereas cyp51B gene consists of 1,747 bp and 3 introns with 63, 56, and 53 bp. The predicted cDNA (1,545 bp) of cyp51A gene encodes a 515-amino acid protein. The cyp51B gene (1,575 bp) codes for a 525-amino acid polypeptide. A comparison between the amino acid sequences of CYP51A and CYP51B of A. clavatus with the other sequences in UniProt protein sequence database showed 82.7% and 89.7% identity with Aspergillus lentulus and A. fumigatus, respectively. The characteristics of all isolates, the MIC ranges, and the types of mutations are presented in Table 1. Alignment of the nucleotide sequences of cyp51A and cyp51B genes from four ITC-susceptible isolates with that of the six drug-high MIC isolates, as well as isolates with MIC = 1 mg/L, showed six missense and three silent mutations in cyp51A and one silent mutation in cyp51B (Table 1). In general, the isolates with itraconazole (ITR) MIC >16 mg/L had more mutations (four clinical and two environmental) than isolates with ITR MIC ≤1 mg/L. Among all mutations detected in the current study, four have already been described (F324S, F53F, E427K, and E506G) in A. fumigatus isolates,16,26,27 whereas six other mutations consisting of V51I, A186A, Q276Q, H359H, L378P, and P486S were original. As shown in Table 1, all isolates with ITR MIC >16 mg/L demonstrated nonsynonymous mutations, including V51I, L378P, E483K, and E506G, and synonymous mutations, including F53F, A186A, Q276Q, and H359H. Furthermore, the P486S change was detected in five strains with ITR MIC >16 mg/L and strain 1189 (Accession No. 322305) with ITR MIC 1 mg/L (Table 1). Remarkably, one specific missense mutation, T971C, translating to F324S, was detected in an isolate with posaconazole MIC >16 mg/L (Accession No. 322306) (Table 1).
Homology modeling and in silico analysis
In the current study, six nonsynonymous mutations were identified in the sequence of cyp51A of different A. clavatus isolates. In this study, for construction of conformational models for cyp51A, we used the amino acid sequences of isolates 92 and 763 as representatives for sensitive and resistant isolates. The X-ray crystallography structure of CYP51B (PDB code: 4UYM) from A. fumigatus was the closest structure for construction of a conformational model of CYP51A for A. clavatus. The sequence identity between the amino acid sequence of CYP51B of A. fumigatus and CYP51A of A. clavatus was 64% and 63% for the 92 and 763 isolates, respectively. The accuracy of the constructed models was verified with SAVE server, the results revealed good geometrical values, also ERRAT analysis showed the overall quality factor of 95.49% and 97.36% for 92 and 763 isolates, respectively. The Ramachandran plots for the models also demonstrated that most of the amino acids were in the favored and allowed regions (Fig. 1). RMSD values for Cα between the template structure (CYP51B) and the modeled CYP51A of isolated 92 and 763 were 0.67 Å and 0.73 Å, respectively; moreover, the RMSD value for Cα between the two models was 0.51 Å. The amino acid comparison between CYP51B of A. fumigatus and CYP51A of A. fumigatus also revealed that those amino acids required for proper structure and function of CYP51A and CYP51B are well conserved (Fig. 2). The conformational structure of CYP51A in A. clavatus shared the common p450-fold seen in other eukaryotes, containing a heme group coordinated with Try114, Try128, lys 140, Arg 376, and His 458. The iron atom in the center of heme is stabilized with four nitrogens within the heme group and also coordinates with Cys460 of CYP51A. The structure of CYP51A also contains two ligand access channels designated channel 1 and channel 2 in Fig. 3. In this study, six mutations were identified and mapped on the structure of modeled CYP51A. E483K and P486S were located near the entry of channel 1, potentially contributing to itraconazole resistance by sterically obstructing drug access. The effects of E483K and P486S substitutions on itraconazole binding to CYP51A were investigated using molecular dynamics analysis and molecular docking studies. The data derived from analysis of the MD trajectory are shown in Figs. 4 and 5. Analysis of the RMSD plot indicated that the native form shows more flexibility in comparison with the mutant form by ∼0.5 Å (Fig. 4). This might be connected to the replaced residues in the mutant form of the protein. Evaluation of Rg of both forms is in accordance with the RMSD plot. Moreover, the data also indicated that the native form of the Cyp51 is more flexible than the mutant form (Fig. 5A). Initial Rg value of the native form began around 22 Å. This value was held relatively constant during the first 36 ns of the simulation and then elevated to near 29 Å until the end of simulation. In the other site, Rg for the mutant form was initiated from 29 Å and remained almost constant until the end of simulation. In accordance with the previous data, the RMSF plot reveals some interesting results, postulating the effect of mutations on the antifungal drug resistance behavior of the mutant Cyp51A protein (Fig. 5B). The overall behavior of the RMSF plot for both forms was similar, but some residues of the native protein fluctuated considerably in comparison with the mutant form. These residues that include T14, I15, P180, V181, F183, S455, and S456 are located at the gate of the active site channel and may act as gate keepers (Fig. 5B). Therefore, their fluctuations might have a significant impact on any substrate, inhibitor, or ligand that needs to reach the active site residues in the channel's depth. This speculation was further investigated by the molecular docking study that confirmed fluctuation is essential for opening the gate of the channel (Fig. 6).

Ramachandran plots for constructed model of isolate 92

Multiple alignment of amino acid sequences of CYP51B of Aspergillus fumigatus and CYP51A of 15 isolates of Aspergillus clavatus. Alpha-helix and beta-strand are shown on the sequence. Those residues participating in the ligand binding pocket are shown with boxes. The position of iron-coordinated cysteine is noted on top of the box. The positions of six nonsynonymous mutations are shown with stars.

Conformational structural of superimposed CYP51B of A. fumigatus (PDB code: 4uym) and CYP51A model of isolate 92 and 763. The position of E483K and P486S mutations has been mapped on the structure with lines. The position of channel 1 and 2 has also been shown relative to the mutation sites. PDB, Protein Data Bank.

Combined backbone RMSD and Rg plot of CYP51A of isolates 92 and 763. RMSD and Rg values were plotted on the left and right axis, respectively. As described within the text, the flexibility of the backbone is relatively similar for both forms (isolates 92 is a little more flexible). The Rg curves show that these proteins had a considerable difference in their gyration radius (more than 7 Å) in the first three-quarters of the simulation time. However, in the last quarter of simulation, both proteins adopted similar Rg values. Rg, radius of gyration; RMSD, root-mean-square deviation.

The RMSF plot of CYP51A of isolates 92 and 763 during the simulation time
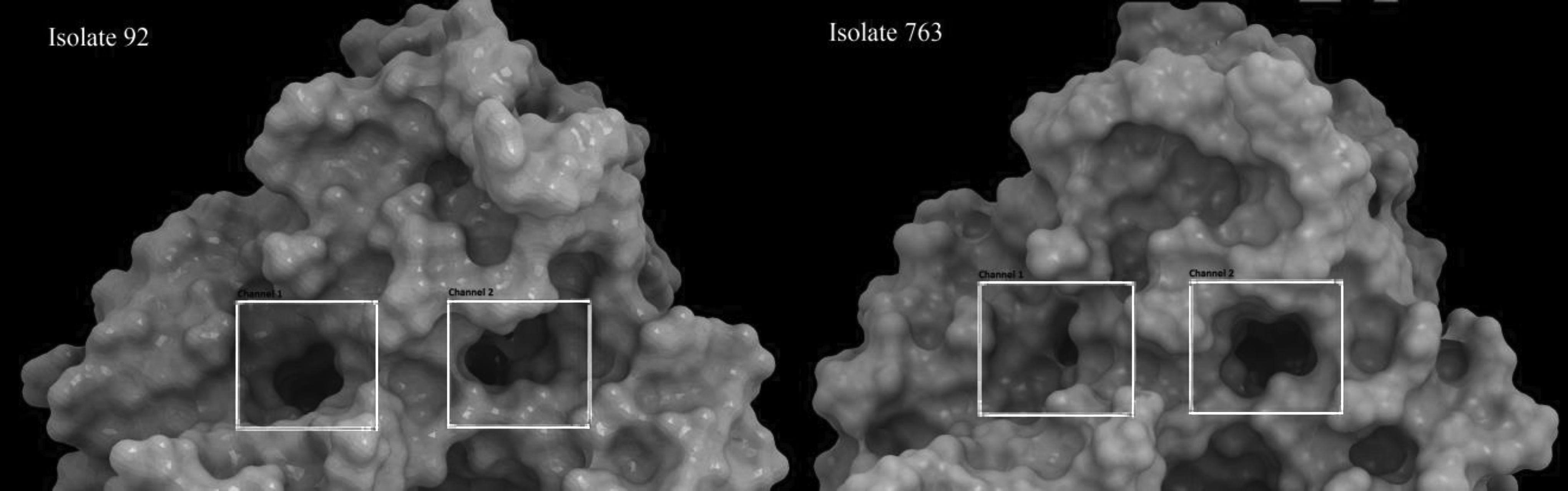
Surface structure of CYP51A of isolates 92 and 763. Surface structure for both proteins was generated after 50 ns MD simulation and the change in the gate of the access channel has been shown with squares. There is a marked decrease in the diameter of the gate of channel 1 in isolate 763 (isolate harboring E483K and P486S mutations).
Discussion
In the current study, the cyp51A and cyp51B genes of A. clavatus coding for 14 α-sterol demethylases A and B were characterized to evaluate associations of variants in these genes with resistance to antifungal drugs. Despite the description of a spectrum of resistance mechanisms to azole drugs in Aspergillus species, azole resistance is commonly associated with mutations in the cyp51A gene.2,28 Congruently, in this study, we found all mutations associated with high MICs to azoles were restricted to CYP51Ap, suggesting that there is a high correlation between these mutations and resistance to itraconazole. The TR34/L98H substitution of the cyp51A gene was first found in the Netherlands 29 and is now the most commonly reported mutation globally. However, Paul et al. 30 demonstrated that Y319H substitution in cyp51C gene was related to azole resistance in A. flavus species. To our knowledge, this is the first report of the three nonsynonymous mutations V51I, L378P, and P486S in the genus Aspergillus worldwide, while E427K, F324S, and E506G substitutions were involved in the resistance of A. fumigatus to itraconazole in the previous studies.16,26,27 The newly detected mutations in A. clavatus strains, including A186A, Q276Q, and H359H, were not responsible for amino acid substitutions.
Interestingly, F219S mutation, which was detected in this study in A. clavatus isolates with high MICs for posaconazole, was previously 26 described for A. fumigatus strains with ITR MIC value >16 mg/L, as well as POS, VOR, and ISA MIC value >2 mg/L.25,30 Lescar et al. demonstrated that Y121F of AfCYP51A decreased susceptibility to only voriconazole, not to itraconazole and posaconazole. 25 Tyrosine 121 is a conserved amino acid in the sequence of AcCYP51A that is essential for coordination with the heme structure. The results of molecular dynamics showed that Y121F leads to a change in voriconazole/heme interaction in the binding pocket. 25 Similarly, the effect of Y121F/T289A mutations has been investigated in another study using in vitro and in silico experiments. 31 It has been stated that Y121F mutation alone, and in combination with T289A, was associated with voriconazole resistance. This phenomenon was supported by homology modeling and molecular docking experiments as Y121 is not only involved in the stabilization of heme structure and overall structure of CYP51A but may also participate in hydrophobic interactions with azoles. 31 Moreover, analysis of the structure of CYP51A and CYP51B of A. fumigatus showed that they contain two ligand access channels, one between helix A′, FG loop, and β4 hairpin 32 and the other between helix K, F, and helix J. 33 In this study, the same access channels were identified for CYP51A of A. clavatus and we assumed that amino acid change surrounding these channels could potentially contribute to limited access of inhibitors to the binding pocket, leading to drug-high MICs. It was reported that L98H in CYP51A of A. fumigatus was linked to azole resistance not because it is located at the ligand binding pocket or ligand access channels, but because it was located on a loop that forms an arch-like structure near the access channel. The leucine to histidine substitution increases the flexibility of the loop, thereby narrowing the diameter of the entry of the ligand access channels. 13 Similarly, molecular dynamics studies have shown that the L98H mutation could partly affect the diameter of ligand access channel of CYP51A, potentially leading to azole resistance. This effect has been documented using site-directed mutagenesis studies on CYP51A. The results showed that introduction of L98H mutation alone and in combination with the 34 bp TR in CYP51A leads to increased azole MIC compared with the wild-type CYP51A. 34 Similarly, in this study, E483K and P486S substitutions located close to access channel are suggested to behave the same way as L98H substitution of CYP51A of A. fumigatus. In contrast, similar to several previous reports,35–39 antifungal activity of new imidazoles, luliconazole and lanoconazole, against all A. clavatus isolates was surprisingly superior to amphotericin B, triazoles, caspofungin, and allylamines, suggesting promising candidates for the treatment of different forms of cutaneous aspergillosis caused by this species.
Our data clearly indicate that amino acid substitutions in CYP51A, V51I, L378P, E483K, and E506G were highly correlated with high MIC values (ITR MIC >16 mg/L), however, further investigations in the future, to explore the other mechanisms, provide new insights of drug resistance or high MICs in A. clavatus strains. The results obtained from molecular dynamics studies performed on the constructed models of CYP51A of susceptible isolates and isolates with high MICs of A. clavatus indicate that although the mutations do not locate directly in the access channel nor to the active site of the CYP51A, the distortions of the ligand access channel made by E483K and P486S mutations could potentially limit access of the azole compounds to the protein in isolates with high MICs.
Footnotes
Acknowledgments
This study was supported by a grant (No. 2786) from the School of Medicine, Mazandaran University of Medical Sciences (Sari, Iran), which we gratefully acknowledge. We thank Seyed Mojtaba Seyed Mousavi for his expert advice.
Ethical Approval
This research was approved by the Ethics Committee of the Mazandaran University of Medical Sciences with the ethical code IR.MAZUMS.REC.95-2786.
Disclosure Statement
No competing financial interests exist.