
Abstract
As resistance to the β-lactam class of antibiotics has become a worldwide problem, multidrug-resistant (MDR) human (n = 243) and food animal (n = 211) isolates from Lagos, Nigeria were further tested to characterize β-lactamase-encoding genes and plasmid replicons. Four β-lactamase-encoding genes (blaCMY, blaCTX-M, blaOXA, and blaTEM) were detected using PCR-based replicon typing, 13 and 17 different replicons were identified using a subset of MDR E. coli from humans (n = 48) and animals (n = 96), respectively. Replicon types FIB and X2 were detected in equal numbers (2/48; 4.2% each) from human isolates, while type Y (16/96; 16.7%) was the most common type from animals. Only two replicon types, FIB and Y, were detected in both groups; all other types were confined to one group or the other, but not both. Using conjugation, replicon type Y, present in three donors, transferred in all three instances, whereas FIA transferred in 75% (3/4) of the matings. This study showed that β-lactamase genes were prevalent in MDR E. coli from both humans and animals in Nigeria and also contained diverse plasmid replicons. As the replicon-associated genes were mobile, they are likely to continue disseminating among E. coli and facilitating transfer of associated β-lactamase genes in this region.
Introduction
Antibiotics have been used to save millions of lives from infectious diseases. They have also been used in farming and agriculture for therapeutic purposes and also as growth promoters in food-producing animals.1–3 Misuse and overuse of antibiotics and other antimicrobials in humans and animals have contributed to resistance and multidrug resistance (MDR) in bacteria, which has compromised therapy in humans and threatened animal health globally.4–6 These issues are exacerbated in resource-limited regions, such as African countries, where other factors exist, including a lack of antibiotics in some areas for definitive treatment and easily procured antibiotics from nonprofessionals for self-medication in other locales. 7 This is especially true when considering the most commonly used class of antibiotics, the β-lactams.
Production of β-lactamases has been reported as one of the most common mechanisms of resistance to β-lactam antibiotics among Enterobacteriaceae, and has played a significant role in the global spread of resistance, particularly in the community and hospital settings as well as in agriculture and the environment.8–10 β-lactamase-encoding genes include extended-spectrum β-lactamases (ESBLs), AmpC β-lactamases (AmpC), and class D β-lactamases (oxacillinases) and can reside on mobile genetic elements, such as plasmids, which aid in dissemination of β-lactamase genes through bacterial conjugation.11–15 Those plasmids belong to various incompatibility groups (Inc); several plasmid families, that is, IncF, IncI1, IncK, IncN, and IncX4, have been associated with the emergence of the ESBL, CTX-M-15, in Enterobacteriaceae including Escherichia coli.13,14
Although data are available on ESBL prevalence from countries in modernized locations, data are limited for African countries such as Nigeria. The majority of studies in Nigeria have focused on clinical ESBL-producing E. coli, the specific ESBL-encoding genes and the replicons involved in their dissemination7,16–21; very few studies on ESBL-producing E. coli and their characteristics from food animals are available.18,22 However, as these studies show that ESBL-producing E. coli are not only associated with humans, but with different animal and environmental sources, monitoring prevalence in these sources is equally important as they are a reservoir of resistant isolates capable of spreading to humans.
In our previous studies, we demonstrated that E. coli from food-producing animals (chicken, cattle, and swine) and humans (animal handlers working in food production in the market and abattoirs in the State and outpatients attending Lagos hospitals) from Nigeria were resistant to multiple antibiotics commonly used to treat human infections.23,24 As certain plasmid types are associated with MDR, the primary aim of this study was to further analyze MDR E. coli from food animals and humans from the previous studies for the presence of β-lactamase-encoding genes and plasmid replicon types. A subset of isolates were also tested to determine if the replicon-associated genes were able to transfer among E. coli. This information will assist in understanding and monitoring the spread of MDR E. coli-producing β-lactamases and ESBLs and plasmid replicon prevalence among food animal and community isolates in Nigeria important to both human and animal health.
Materials and Methods
Bacterial isolates
E. coli from food animals (n = 211) and humans (n = 243) and their antimicrobial resistance patterns were as previously described.23,24 All bacterial strains were stored at −80°C in Luria-Bertani (LB) broth containing 30% glycerol until use. Isolates included in this study were resistant to two or more of the following antimicrobials: ampicillin (≥32 μg/mL), ampicillin/sulbactam (≥32/16 μg/mL), aztreonam (≥16 μg/mL), cefazolin (≥8 μg/mL), cefepime (≥32 μg/mL), cefoxitin (≥32 μg/mL), cefpodoxime (≥8 μg/mL), ceftazidime (≥16 μg/mL), ceftriaxone (≥4 μg/mL), cefuroxime (≥32 μg/mL), cephalothin (>16 μg/mL), ciprofloxacin (≥4 μg/mL), ertapenem (≥2 μg/mL), gentamicin (≥16 μg/mL), meropenem (≥4 μg/mL), piperacillin/tazobactam (≥128/4 μg/mL), tetracycline (≥16 μg/mL), ticarcillin/clavulanic acid (≥128/2 μg/mL), tobramycin (≥16 μg/mL), trimethoprim/sulfamethoxazole (≥4/76 μg/mL), and tigecycline (≥1 μg/mL).
Resistance gene detection
Whole-cell DNA template was prepared by suspending a single bacterial colony in 100 μL of sterile deionized water; 5 μL of template was used per reaction. The presence of blaCMY, 25 blaCTX-M, 26 blaKPC, 27 blaNDM, 28 blaOXA, 29 blaSHV, 30 and blaTEM 31 was determined as described using PCR and DNA sequencing for confirmation of the gene identity.
PCR-based replicon typing-based replicon typing
A subset of the MDR E. coli from animals (n = 96; resistant to ≥2 antimicrobials) and humans (n = 48; resistant to ≥5 antimicrobials) previously characterized using molecular methods23,24 were subjected to plasmid replicon analysis. MDR E. coli were examined for the presence of plasmid replicons of 25 different types, which are associated with incompatibility groups using the PCR-Based Replicon Typing Kit (Diatheva, Fano, Italy) according to the manufacturer's directions.
Bacterial matings
Isolates designated for use as donors in mating experiments to study transfer of replicon-associated genes were those containing the most frequently detected replicon types among the MDR isolates as well as the two common types (FIB and Y) among the two sources. Isolates selected for matings were previously analyzed using whole-genome sequencing and were blaCTX-M positive as this gene is frequently plasmid mediated. 32 Three MDR E. coli from animals and seven from humans meeting these criteria were selected. Transconjugants were selected using ampicillin as all donors were resistant to this antibiotic.
Conjugations were performed using filter mating as previously described using E. coli J53 AZR (sodium azide resistant) as the recipient strain.33,34 In brief, donor and recipient cells (1:1 dilution; 1–5 × 107 cells/mL) were mixed in brain–heart infusion (BHI) broth, and 100 μL of the mixture was inoculated onto a sterile 0.45 μm pore size nylon filter positioned on the surface of a blood agar plate. The plates were incubated overnight at 37°C without antimicrobials. Cells were removed from the filters by washing in 2 mL of BHI; 100 μL of the mixture was plated onto LB agar plates containing ampicillin (32 μg/mL) plus sodium azide (100 μg/mL). Plates containing only ampicillin or sodium azide served as controls for donors and the recipient.
Transconjugants were phenotypically tested for ESBL production by broth microdilution using the Sensititre™ ESBL panel ESB1F and according to manufacturer's guidelines.35,36 This susceptibility plate was used for ESBL confirmation and contained cephems (ceftazidime, cefazolin, cefepime, cefoxitin, cephalothin, cefpodoxime, cefotaxime, ceftriaxone), carbapenems (imipenem, meropenem), aminoglycoside (gentamicin), penicillin (ampicillin), fluoroquinolone (ciprofloxacin), and β-lactam/β-lactamase inhibitors (piperacillin/tazobactam constant 4, ceftazidime/clavulanic acid, cefotaxime/clavulanic acid). A greater than or equal to eightfold reduction in the minimum inhibitory concentration (MIC) of cefotaxime or ceftazidime combined with clavulanic acid compared with the MIC of the cephalosporin alone was considered a positive phenotype for ESBL production. β-lactam resistance genes and replicon PCR analysis were performed on the transconjugants to verify the presence of β-lactamases and replicon types.
Results
β-Lactamase-encoding genes
Approximately 29% and 15% of E. coli from food animals and humans, respectively, were negative for the seven β-lactamase-encoding genes tested (Table 1). Isolates were negative for blaKPC, blaNDM, or blaSHV; however, gene profiles for isolates containing a β-lactamase-encoding gene were similar for E. coli from both sources. For isolates containing only a single gene, blaCMY was detected most often in E. coli from animals (92/211; 43.3%) and humans (98/243; 40.3%) followed by blaTEM at 9.5% and 8.2%, respectively; blaCTX-M or blaOXA was rarely detected (Table 1). For combinations of two or more genes, only three combinations were observed among E. coli from animals while E. coli from humans were more diverse containing eight different combinations, all possible combinations except blaCTX-M, blaOXA. The two most prevalent single β-lactamase-encoding genes were also the most frequently detected gene combination (blaCMY, blaTEM), which was found in higher numbers in human E. coli (57; 23.5%) than in animal E. coli (30; 14.3%). One isolate from animals and six isolates from humans contained all four genes, blaCMY, blaCTX-M, blaOXA, and blaTEM (Table 1).
β-Lactamase Gene Profiles of Escherichia coli from Animals and Humans
Plasmid replicon profiles
Approximately 14.6% (14/96) of MDR E. coli from animals and 22.9% (11/48) from humans were negative for all 25 replicon types tested. Overall, plasmid replicons from animals were more diverse than those from humans. Eleven single replicon types were identified from animals compared with five from humans. Only two replicon types, FIB and Y, were detected in both groups; all other types were confined to one group or the other, but not both (Table 2). Replicon type Y (16/96; 16.7%) was the most common type detected in animals, while types FIB and X2 were detected in equal numbers (2/48; 4.2% each) in MDR E. coli from humans. The difference in diversity among the single replicon types for each source was also reflected in the multireplicon profiles (composed of ≥2 replicon types). Thirty different replicon combinations were present in E. coli from animals, whereas 15 combinations were found in humans. Combinations ranged from two (21 total patterns) to as many as five replicon types (two patterns) per isolate (Table 2). Replicon profiles composed of two replicons were more frequently detected than profiles containing more than two. However, replicon profiles for MDR E. coli from animals and humans containing two replicon types were more frequently detected than single replicon types for each group (16 vs. 11 for animals and 8 vs. 5 for humans). The most common pattern for animal E. coli was FIB, FII (4/96; 4.2%), while FIA, FII (4/48; 8.3%) was the most common from humans.
Plasmid Replicon Profiles of Escherichia coli from Animals and Humans
For profiles containing three replicon types, the most common was FIA, FIB, FII (9/48; 18.8%) from humans, while the six profiles identified from animals were fairly evenly distributed with just one or two isolates represented in each group. Two replicon profiles composed of five replicon types (FIA, FIB, FIC, FII, I1 and FIB, FII, FIIS, I1, P) were each detected in single isolates from animals only (Table 2). Of the 41 different combinations of replicon types, four types (FIA, FII; FIB, FII; FIB, X2; and FIB, FII, XI) were common between the two sources. Replicon types A/C and T were only detected paired with other replicons (T, FIIS, FII; A/C, FIA, FII, XI and A/C, FIB, I1, T).
Plasmid replicon and β-lactam diversity
Plasmid replicon profiles and associated β-lactamase-encoding genes of MDR E. coli from food animals are shown in Table 3. Four configurations of these characteristics were observed among the isolates, including absence of replicons and β-lactamase-encoding genes, presence of replicons and β-lactamase-encoding genes, β-lactamase genes without a corresponding replicon type, and single and multireplicons without a detected β-lactamase gene. For example, isolate 141 was resistant to five classes of antibiotics, contained two replicon types, and all four detected β-lactamase-encoding genes (Table 3). Isolate 100 was negative for all replicon types tested, but was positive for blaCMY and blaTEM. In contrast, isolates 198 and 246 both contained five replicon types, but only blaCMY and blaTEM, respectively. Two isolates, 285i and 290, which were negative for replicon types as well as β-lactamase genes, exhibited resistance to one β-lactam, ampicillin, although the isolates were resistant to other drug classes. A third isolate, 297, was also negative for replicons and β-lactamase genes, but this was not unexpected as it was not resistant to any β-lactam antibiotics (Table 3).
Plasmid Replicon Profiles and β-Lactam Resistance Genes of Multidrug-Resistant Escherichia coli from Food Animals
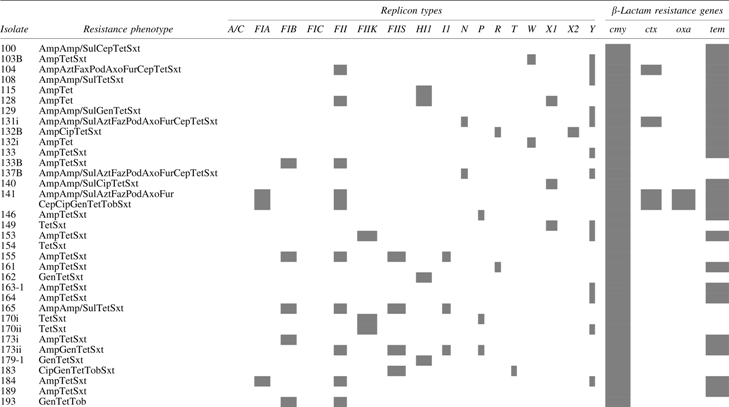


Highlighted cells indicate presence of the replicon type or resistance gene.
Amp, ampicillin; Amp/Sul, ampicillin/sulbactam; Axo, ceftriaxone; Azt, aztreonam; Cep, cephalothin; Cip, ciprofloxacin; Faz, cefazolin; Fox, cefoxitin; Fur, cefuroxime; Gen, gentamicin; P/T4, piperacillin/tazobactam; Pod, cefpodoxime; Sxt, trimethoprim/sulfamethoxazole; Tet, tetracycline; Tim, ticarcillin/clavulanic acid; Tob, tobramycin.
Combinational patterns of replicon types and β-lactamase-encoding genes of MDR E. coli from humans differed from those from animals (Table 4). Isolates were either positive for replicon types and resistance genes or positive for resistance genes without a replicon type present. None of the isolates were negative for both or negative for a replicon type while also containing a β-lactamase-encoding gene. Isolates that were positive for all four resistance genes contained various numbers of replicons as well as replicon types. Three isolates (38.ii.h, 43.ii.h, and 45.h) all contained FIA and FII, while isolate 129.h was positive for I2 and isolate 382.h was positive for FII and HI2. Isolate 253.h was negative for all tested replicon types, but contained blaCMY, blaCTX-M, blaOXA, and blaTEM (Table 4). Further, all six isolates were resistant to multiple β-lactam antibiotics. Replicons were not detected in ten additional isolates (49.h, 236.h, 243i.h, 243ii.h, 246.h, 247.h, 250i.h, 301.i.h, 307i.h, and 307ii.h) in which 1–3 β-lactamase-encoding genes were detected; these isolates also exhibited resistance to various β-lactam antibiotics. The number of antibiotics for which an isolate displayed resistance did not appear to determine the number of plasmid replicons harbored by the isolate.
Plasmid Replicon Profiles and β-Lactam Resistance Genes of Multidrug-Resistant Escherichia coli from Humans


Highlighted cells indicate presence of the replicon type or resistance gene.
Amp, ampicillin; Amp/Sul, ampicillin/sulbactam; Azt, aztreonam; Faz, cefazolin; Fep, cefepime; Fox, cefoxitin; Axo, ceftriaxone; Fur, cefuroxime; Cep, cephalothin; Cip, ciprofloxacin; Pod, cefpodoxime; Taz, ceftazidime; Ert, ertapenem; Gen, gentamicin; Mer, meropenem; Tet, tetracycline; Tig, tigecycline; Tim, ticarcillin/clavulanic acid; Tob, tobramycin; Sxt, trimethoprim/sulfamethoxazole; Fep, cefepime; Ert, ertapenem; Mer, meropenem; Taz, ceftazidime; Tig, tigecycline.
Transfer of plasmid replicons and β-lactamase-encoding genes
The three MDR E. coli donors from animals originated from chickens and contained two replicons each. Replicon types N, Y and FIA, FII transferred from isolates 131i and 141, respectively (Table 5). However, although isolate 104 also contained FII and Y, only type Y was detected in the transconjugants. For the donors from humans, all replicon types transferred from isolates 38.ii.h, 129.h, 201ii.h, and 322i.h. Two of four replicon types transferred from isolate 29B.h, including I2 and X1; replicon types FIA and FIB were not present in the transconjugants from this mating (Table 5). While FIA and FII were identified in transconjugants using donor isolate 299.h, replicon type HI2 failed to move and was not detected. In contrast, HI2 was transferred from isolate 382.h, whereas the second replicon type present in the isolate, FII, was not exchanged.
Transfer of Antimicrobial Resistance, β-Lactam Resistance Genes, and Replicons in Multidrug-Resistant Escherichia coli
Transconjugants were selected using resistance to ampicillin (32 μg/mL). Bolded antimicrobials indicate resistance in the transconjugant not observed in the donor (one-well difference in minimum inhibitory concentration).
Amp, ampicillin; Amp/Sul, ampicillin/sulbactam; Azt, aztreonam; Faz, cefazolin; Pod, cefpodoxime; Axo, ceftriaxone; Fur, cefuroxime; Cep, cephalothin; Cip, ciprofloxacin; Gen, gentamicin; Tet, tetracycline; Tim, ticarcillin/clavulanic acid; Tob, tobramycin; Sxt, trimethoprim/sulfamethoxazole.
Overall, replicon type Y, present in three donors, transferred in all three instances, whereas FIA moved in 75% (3/4) of the matings and FII in 60% (3/5). Transconjugants of isolates 131i and 141 from chickens, and isolates 38.ii.h, 129.h, 299.h, and 382.h from humans, displayed similar resistance gene profiles to their donors (Table 5). Donor isolates from which only a single β-lactamase gene transferred (blaCMY) were 104 from chicken and 29B.h, 204ii.h, and 322i.h from humans (Table 5). ESBL phenotypes (MIC ratio ≥8 for cefotaxime and ceftazidime with and without clavulanic acid) were identified in seven transconjugants, two from chickens (131i and 141) and five from humans (38.ii.h, 129.h, 299.h, 322i.h, and 382.h), which all have ESBL resistance genes transferred from their donors except human isolate 322i.h (Supplementary Table S1). Phenotypic differences between some donors and resulting transconjugants were due to a one-well difference in MIC.
Discussion
To our knowledge, this is the largest study conducted on MDR E. coli-producing β-lactamases in the Nigerian community and one of the few studies on characterization of plasmid replicons in this country.18–20 Combating antimicrobial resistance requires a multifactorial approach, which includes monitoring or surveillance of resistance to antibiotics together with the resistant bacteria, resistance genes, and mobile elements harboring those genes. 37 This presents a challenge for industrialized countries, but is even more difficult in financially challenged countries, such as Nigeria, where capabilities to conduct large studies are lacking. However, smaller, regional studies provide data on resistance from different sources, which can help predict the scope of the problem and the impact on humans, animals, and the environment. In this study, data are provided on prevalence and identification of β-lactamase resistance genes and plasmid replicons present in MDR E. coli from both humans and food-producing animals. Although specific experiments, such as Southern analysis or genome sequencing, to localize the genes to the exact replicon were not performed, multiple β-lactamase genes coexisting with a diversity of plasmids known to facilitate dissemination of resistance determinants were detected, suggesting that the replicons harbored the resistance genes; conjugation experiments verified the mobility of the replicons.
Studies on prevalence of ESBL-producing E. coli and β-lactamase-encoding resistance genes from both clinical and animal samples in Africa have been recently reviewed.7,16,38,39 Previous studies have reported prevalence of E. coli-producing ESBLs in Nigeria ranging from 10.3% to 27.5% in human samples and 0.5% to 20.57% in samples from animals.7,38 However, prevalence of β-lactamase-producing E. coli from our previous studies was much higher with 35% and 46.6% of animal and human samples, respectively, containing resistant isolates.23,24 This could be due to the larger sample sets tested or methodology used in our studies. Furthermore, β-lactamase-encoding genes identified from this study were not the ones predominantly identified in previous studies from Nigeria. Those resistance genes belonged to the blaCTX-M family, specifically blaCTX-M-15, and were by far the most frequently detected gene in E. coli from both animal and clinical samples.21,22,40–43 This gene was a rare occurrence in this study as blaCTX-M was only detected in three isolates from animals.
Of the seven β-lactamase genes tested in this study, blaCMY was identified most often from both sources. This gene family has been reported as one of the most extensively detected in the β-lactamase gene group for E. coli from animals worldwide conferring resistance to penicillin and cephalosporins, although they are not considered ESBLs due to their different substrate utilization and inhibitor profile. 44 This unclear definition of plasmid-borne AmpCs resulting in difficulty in phenotypic characterization may have led to miscalculations in prevalence of AmpC genes. 45 A Clinical and Laboratory Standards Institute-validated phenotypic test to confirm the presence of plasmid-encoded AmpC β-lactamase in patient isolates is also unavailable. 46 In humans, blaCMY-2 is the primary plasmid-borne AmpC gene encountered and has been detected in conjunction with ESBLs in the same bacterial cell.45,47 Coexistence of blaCMY with ESBLs was also observed in this study as blaCMY was identified in E. coli isolates along with blaCTX-M, blaOXA, and blaTEM. However, Enterobacteriaceae containing a combination of blaCMY with any ESBL appears rare in Nigeria as only two reports have been noted for clinical samples.48,49
While ESBLs including blaKPC, blaOXA, and blaSHV have been previously detected in Enterobacteriaceae in clinical isolates in Nigeria, 16 MDR E. coli in this study were negative for blaKPC and blaSHV. The presence of single and multiple β-lactamase genes in these isolates suggests that β-lactam antibiotics may not be a viable treatment option and may lead to ineffective antimicrobial treatment, consequently resulting in increased mortality in clinical infections in this country.50,51
Reports on plasmid replicons in Nigeria are severely limited. Chah et al. 20 conducted a study on 11 E. coli strains and reported eight different plasmid replicon types, B/O, FIB, FIC, I1, K, W, X, and Y, with IncF being the most frequently detected in nine of the isolates. The prevalence of low copy plasmids with multiple replicon-associated genes, such as IncF, aiding dissemination of resistance genes detected among the isolates in this study may be a potential factor for regional dissemination of antibiotic resistance and emergence of relevant resistance determinants. 11 The IncF group were found to be the most common plasmids associated with β-lactamase genes in this study and often carried more than one replicon-associated gene as previously reported. 11 The presence of multiple replicon-associated genes promotes initiation of replication as well as plasmid maintenance, but also limits the host range to the genera of Enterobacteriaceae.52–54 IncF plasmids have been previously linked with blaCTX-M in clinical E. coli in Nigeria, which is a common occurrence worldwide.19,55 This plasmid family, specifically FII replicons, is further characterized by their association with the ESBL-producing E. coli virulent clone ST131. 55 This was also observed in this study as four clinical isolates (38.ii.h, 43.ii.h, 43.iii.h, and 45h), previously identified as ST131, 24 contained FIA and FII as well as two or more ESBL genes. The identification of IncF plasmids among isolates of both human and food-producing animal origin suggests that the plasmids may have a reservoir in the food chain.12,56,57
Other prominent replicon families detected in this study (i.e., HI, X, and Y) have also been previously associated with dissemination of β-lactamase-producing E. coli from both human and animal sources. 11 The IncHI family is a major group of plasmids that can harbor ESBL genes, specifically CTX-M, as well as other antimicrobial resistance genes. 11 Y plasmids have also been associated with blaCTX-M genes, but blaTEM variants have also been localized to these plasmids in clinical E. coli.58,59 This was observed in this study in which Y replicons and blaTEM were colocated in the majority of E. coli from animals and the sole IncY-positive E. coli from humans. Much less studied than the IncHI and IncY families are the narrow host range IncX plasmids. 60 These plasmids are known for their ability to induce biofilm formation and to mobilize to other bacteria at a high rate due in part to their type IV fimbriae allowing for conjugal self-transfer and dissemination of resistance genes.61–64
Transconjugant phenotypes for isolates 131i, 38.ii.h, 129.h, 204ii.h, and 383.h were different from the donor for some antimicrobials due to a change in MIC. For example, the MIC for tobramycin for isolate 38.ii.h was 8 μg/mL for the donor but >8 μg/mL in the transconjugant, resulting in the change from intermediate resistance to full resistance. This could be a result of differences in gene expression between the isolates. Of the ten MDR E. coli selected for conjugation studies, replicon-associated genes transferred from each isolate, but not in a consistent pattern. One pattern that was noticed was for isolates with multiple IncF replicons. For example, FIA and FII were present in the donor isolates 141, 38.ii.h, and 299.h and the resulting transconjugants. Although FIA and FIB were detected in isolate 29B.h, neither were identified in the transconjugants. Taken together, these results suggested that FIA, FII and FIA, FIB replicon types were located on the same plasmid, respectively; however, additional data are needed for confirmation. The overall ability of the replicon types to transfer in this study supported the conclusion that the plasmids contribute to the spread of resistance determinants, promoting horizontal gene transfer among bacteria. These plasmids belong to families that are largely prevalent in commensal and pathogenic bacteria, and usually carry multiple physically linked genetic determinants, which confer resistance to different classes of antibiotics simultaneously. 13 In addition, the presence of multiple replicon types in a single organism allows for greater opportunity for dissemination of plasmids, suggesting that E. coli may serve as plasmid reservoirs.
Conclusion
In conclusion, this study showed that β-lactamase genes were prevalent in MDR E. coli from both humans and animals in Nigeria and also contained diverse plasmid replicons. As the replicon-associated genes were mobile, they are likely to continue disseminating among E. coli and facilitating transfer of associated β-lactamase genes in this region. Constant and indiscriminate use of antimicrobials among humans and ease of global travel also contribute to dissemination of resistance. Increased efforts at antimicrobial surveillance, antibiotic stewardship, and antibiotic use regulation in food animals in Nigeria are urgently needed.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.