
Abstract
The emergence of New Delhi metallo-β-lactamase 1 on carbapenemase-producing bacteria has raised a major worldwide public health concern. This study reports the dissemination of blaNDM-1 in carbapenem-resistant isolates that caused nosocomial infections in a tertiary hospital in Mexico City. Seven Enterobacter cloacae and three Klebsiella pneumoniae nosocomial isolates from the same time period harbored the blaNDM-1 gene. The resistance phenotype and the blaNDM-1 gene were transferred through conjugative plasmids belonging to the incompatibility group IncFIA of 85, 101, and 195 kb in E. cloacae and 95 and 101 kb in K. pneumoniae isolates. Restriction fragment length polymorphism analysis showed that blaNDM-1 was carried in similar plasmids with molecular sizes of 101 and 85 kb, each one in three isolates of E. cloacae and one of 101 kb on two isolates of K. pneumoniae. During a 9-month period, six of the seven isolates of E. cloacae analyzed harbored blaNDM-1 and belonged to clone E1. Similarly, over a 5-month period, two of the three K. pneumoniae isolates that harbored blaNDM-1 belonged to clone K1. These results demonstrate the horizontal transfer of blaNDM-1 between different bacterial species, dissemination of clones with high levels of resistance to carbapenems, and underscore the need for heightened measures to control their further spread.
Introduction
New Delhi metallo-β-lactamase 1 (NDM-1) is one of the most recently discovered class B β-lactamases. Unlike class A, C, and D β-lactamases, NDM-1 has zinc ions in its active site and its greatest clinical significance lies in its ability to hydrolyze all β-lactam antimicrobials, except for monobactams. 1 NDM-1 was first described in a clinical isolate of Klebsiella pneumoniae in a 59-year-old male patient who returned to Sweden from India where he was hospitalized in 2008. This patient was colonized simultaneously with an NDM-1 strain of Escherichia coli in stools. 2
The blaNDM-1 genes are usually found in plasmids (55–500 kb) and are rarely integrated into the chromosome. 3 They have a wide range of hosts and are easily transmitted by conjugation, which favors interspecies dispersion.1,4 Evolution of NDM currently shows the identification of 19 variants of this enzyme in bacteria of food animal origin.5,6 NDM-1 is found predominantly in Enterobacteriaceae and less frequently in Acinetobacter spp., Pseudomonas spp., Stenotrophomonas spp., Aeromonas spp., and Vibrio cholerae. This distribution reflects the association of blaNDM-1 with promiscuous plasmids.1,7
There are several reports from Mexico regarding the presence of NDM-1 in carbapenem-resistant clinical Enterobacteriaceae isolates, each one in different geographical areas of the country and only one in Mexico City.8–13 The objective of this study was to investigate the occurrence of carbapenem-resistant K. pneumoniae and Enterobacter cloacae isolates in a tertiary care hospital in Mexico City and to characterize the genetic nature of carbapenem resistance.
Materials and Methods
Bacterial collection
Single nosocomial infection isolates of K. pneumoniae and E. cloacae were collected between February 2015 and January 2016 at Hospital Regional Gral. Ignacio Zaragoza of the Instituto de Seguridad y Servicios Sociales de los Trabajadores del Estado (ISSSTE) in Mexico City, a 360-bed tertiary referral hospital.
Susceptibility testing
Identification and antimicrobial susceptibility were carried out using the VITEK® 2 system (Healthcare; BIOMÉRIEUX). Colistin susceptibility tests were performed by the broth microdilution method following guidelines of the Clinical and Laboratory Standards Institute (CLSI) and the European Committee on Antimicrobial Susceptibility Testing (EUCAST).14,15 Metallo-β-lactamase (MBL) production in carbapenem-intermediate or -resistant isolates was determined by the disk diffusion test on agar using meropenem and imipenem with or without EDTA. 16
Detection of genetic determinants of antimicrobial resistance
Using the Primer3 program, specific oligonucleotides were designed to determine, by means of PCR, the presence of blaIMP, blaVIM, blaKPC, blaOXA-40-like, and blaNDM-1 genes (Table 1). Amplification of PCR products was carried out using the GoTaq® Green Master Mix Kit (Promega, Madison, WI). The amplification conditions were as follows: one cycle at 95°C for 5 min, 35 cycles at 95°C for 1 min, 58°C for 1 min, and 72°C for 1 min, and a final extension step at 72°C for 15 min. Amplification reactions were carried out using an iCycler (Bio-Rad®, Hercules, CA) thermocycler and the resulting products were resolved by electrophoresis in 1% agarose gels at 120 V using a Power Station 300 chamber (Labnet International, Inc.). The mcr-1 gene was screened by PCR with the primers CLR5-F 5′-CGGTCAGTCCGTTTGTTC-3′ and CLR5-R 5′-CTTGGTCGGTCTGTAGGG-3′ described previously. 17 Amplified fragments were purified and subjected to nucleotide sequencing at the Instituto de Biotecnología, Universidad Nacional Autónoma de México. Nucleotide sequence analysis was performed using the BLAST program of the National Center for Biotechnology Information.a
Primers Designed in This Study to Detect Carbapenem Resistance Genes
bp, base pairs.
Plasmid analysis and conjugation assays
To identify whether the strains were carrying blaNDM-1 in plasmids, the electrophoretic pattern of plasmids was determined using the Eckhardt technique. 18 The bacterial artificial chromosomes of 67, 86, 101, 122, 145, and 195 Kb were used as molecular weight markers. 19 The conjugation assays for horizontal transfer of carbapenem resistance were performed following the method described by Miller 20 using the rifampicin-resistant E. coli strain J53-2 as the recipient strain. Transconjugant strains were selected on Luria-Bertani agar supplemented with rifampicin (200 μg/mL) and imipenem (16 μg/mL) or meropenem (16 μg/mL) and tested for antimicrobial susceptibility. Plasmids of transconjugant strains were purified using the QIAGEN Plasmid Midi Kit (Qiagen, Hilden, Germany), following the manufacturer's specifications, and analyzed by restriction fragment length polymorphism (RFLP) 21 with the restriction enzymes EcoRI and BglII. The percentage of similarity restriction profile was calculated using the Dice coefficient. 22 Transconjugant plasmids with a Dice similarity coefficient >85% were considered as having the same restriction profile.
The incompatibility groups were determined using a PCR-based replicon typing method 23 on transconjugants with a single plasmid encoding blaNDM-1.
Southern hybridization was performed in blaNDM-1-positive isolates and its transconjugants using electrophoretic profiles of Eckhardt plasmids transferred to a nylon membrane and a nonradioactive probe of blaNDM-1 (ECL direct nucleic acid labeling and detection system; GE Healthcare, Piscataway, NJ).
Pulsed-field gel electrophoresis
To evaluate clonality between blaNDM-1-positive strains, genotyping of each of the isolates was carried out using pulsed-field gel electrophoresis (PFGE). 24 Chromosomal DNA of each strain was prepared as previously reported 25 and digested with Xba I restriction endonuclease (New England Biolabs, Beverly, MA). Restriction fragment was resolved in a GenePath System (Bio-Rad®, Hercules, CA). The classification of isolates in clones was based on Tenover criteria. 26 The percentage of similarity profile was calculated using the Dice coefficient. 22 Isolates with a Dice similarity coefficient >85% were considered as belonging to the same clone.
Results
Clinical isolates and antibiotic susceptibility
During the study period, we recovered a total of 27 K. pneumoniae and 12 E. cloacae isolates from nosocomial infections that caused mainly bacteremia, pneumonia, and wound infections. The majority of these isolates were multidrug resistant (Table 2).
Minimum Inhibitory Concentration Data of Antibiotics for Klebsiella pneumoniae and Enterobacter cloacae Isolates Investigated in This Study
IR, intermediate resistant; MIC, minimum inhibitory concentration; N/D, not determined; R, resistant.
Susceptibility results showed that 3/27 (11%) of K. pneumoniae and 7/12 (58%) of E. cloacae isolates were carbapenem resistant. Colistin resistance was observed only 3/42 (7%) K. pneumoniae. Three K. pneumoniae isolates and seven E. cloacae isolates were blaNDM-1 positive, all of them were resistant to most antibiotics, including β-lactams (penicillin, carbapenems, cephalosporins, and monobactams), β-lactam combination agents, aminoglycosides, fluoroquinolones, sulfonamides, and trimethoprim (Table 3). Resistance to tigecycline was observed in 2/3 and 1/7 K. pneumoniae and E. cloacae blaNDM-1-positive isolates, respectively. All blaNDM-1-positive K. pneumoniae isolates were colistin resistant, while all blaNDM-1-positive E. cloacae isolates were susceptible to it. The transconjugants that were obtained from K. pneumoniae expressed resistance to β-lactams and aminoglycosides, although at lower levels than the clinical isolates. As for E. cloacae transconjugants, five of the seven expressed resistance to β-lactams and aminoglycosides, while two were only β-lactam resistant.
Antimicrobial Susceptibility Patterns of Klebsiella pneumoniae and Enterobacter cloacae Positive for blaNDM-1 and Their Transconjugants
AMK, amikacin; AMS, ampicillin–sulbactam; AZT, aztreonam; CRO, ceftriaxone; CFZ, cefazolin; CL, colistin; CIP, ciprofloxacin; Ecl, E. cloacae; FEP, cefepime; GEN, gentamicin; IPM, imipenem; Kpn, K. pneumoniae; MEM, meropenem; ND, not determined; NIT, nitrofurantoin; PTZ, piperacillin–tazobactam; Tc, transconjugant; TGC, tigecycline; SXT, trimethoprim–sulfamethoxazole; TOB, tobramycin.
The majority of blaNDM-1-positive isolates were obtained in patients who were admitted in general surgery and orthopedic wards with four isolates from each ward, followed by the intensive care unit (ICU) with two isolates (Table 4).
Characteristics of blaNDM-1-Producing Klebsiella pneumoniae and Enterobacter cloacae Isolates
Determination of carbapenemase and mcr-1 genes
PCR screening detected the blaNDM-1 gene in all carbapenem-resistant K. pneumoniae and E. cloacae strains, three and seven, respectively. The blaIMP, blaVIM, blaKPC, and blaOXA-40-like genes were not detected by PCR in any of these strains. None of the three blaNDM-1-positive and colistin-resistant K. pneumoniae isolates harbored mcr-1.
Plasmid analysis and conjugative experiments
The plasmid profiles of three K. pneumoniae and seven E. cloacae isolates that were positive for blaNDM-1 harbored between one and three plasmids with sizes of 40, 85, 95, 101, 122, 145, 180, 195, and >195 kb, respectively (Table 4). Plasmids harboring blaNDM-1 from all K. pneumoniae and E. cloacae isolates were successfully transferred to E. coli J53, transconjugants received only one plasmid (Fig. 1A upper, B upper, and Table 4). Acquisition of the blaNDM-1 gene in all transconjugants was detected by PCR and corroborated by nucleotide sequencing. Phenotypic testing of transconjugants exhibited resistance to β-lactams and aminoglycosides (Table 3). Southern hybridization analysis with a blaNDM-1-specific probe confirmed the location of blaNDM-1 in the plasmid transferred by conjugation in all transconjugants (Fig. 1A lower, B lower). The plasmid incompatibility groups were identified in all transconjugants with blaNDM-1-carrying plasmids and corresponded to the IncFIA group (Table 4).
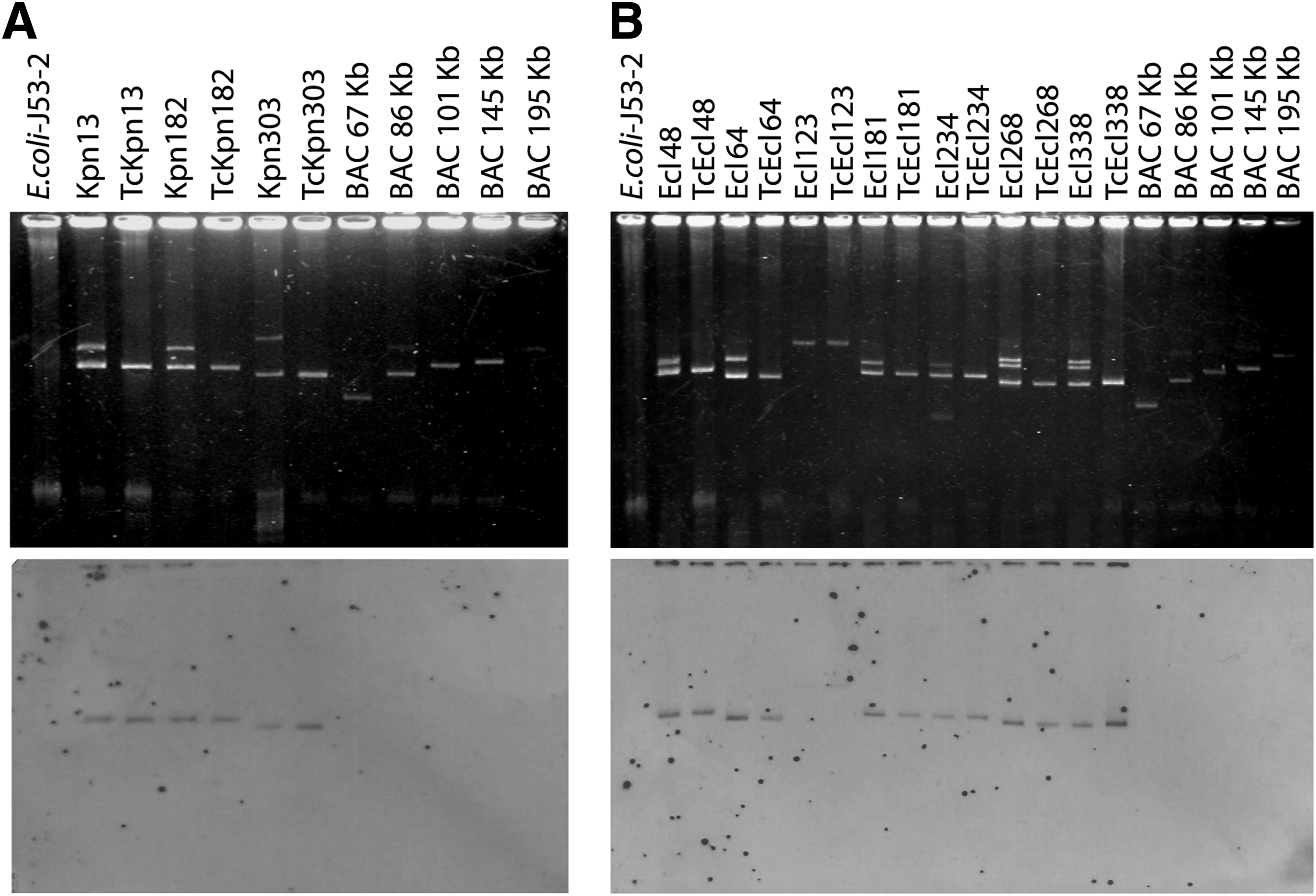
Detection of blaNDM-1 in conjugative plasmids of Klebsiella pneumoniae and Enterobacter cloacae isolates.
RFLP profiles of the 10 conjugative plasmids that carry blaNDM-1 revealed three different restriction profiles. Figure 2 shows the restriction profiles obtained from EcoRI-digested transconjugant plasmids. The restriction profile P1 contained five clusters; in this profile, two transconjugant plasmids with different sizes were grouped: 101 kb (TcEcl 48, TcEcl 181, TcEcl 234, TcKpn 182, TcKpn 13) and 85 kb (TcEcl 268, TcEcl 338, TcEcl 64). Unique profiles (P2 and P3) were also obtained from the transconjugant plasmids of 95 kb (TcKpn 303) and 195 kb (TcEcl 123), respectively (Fig. 2).
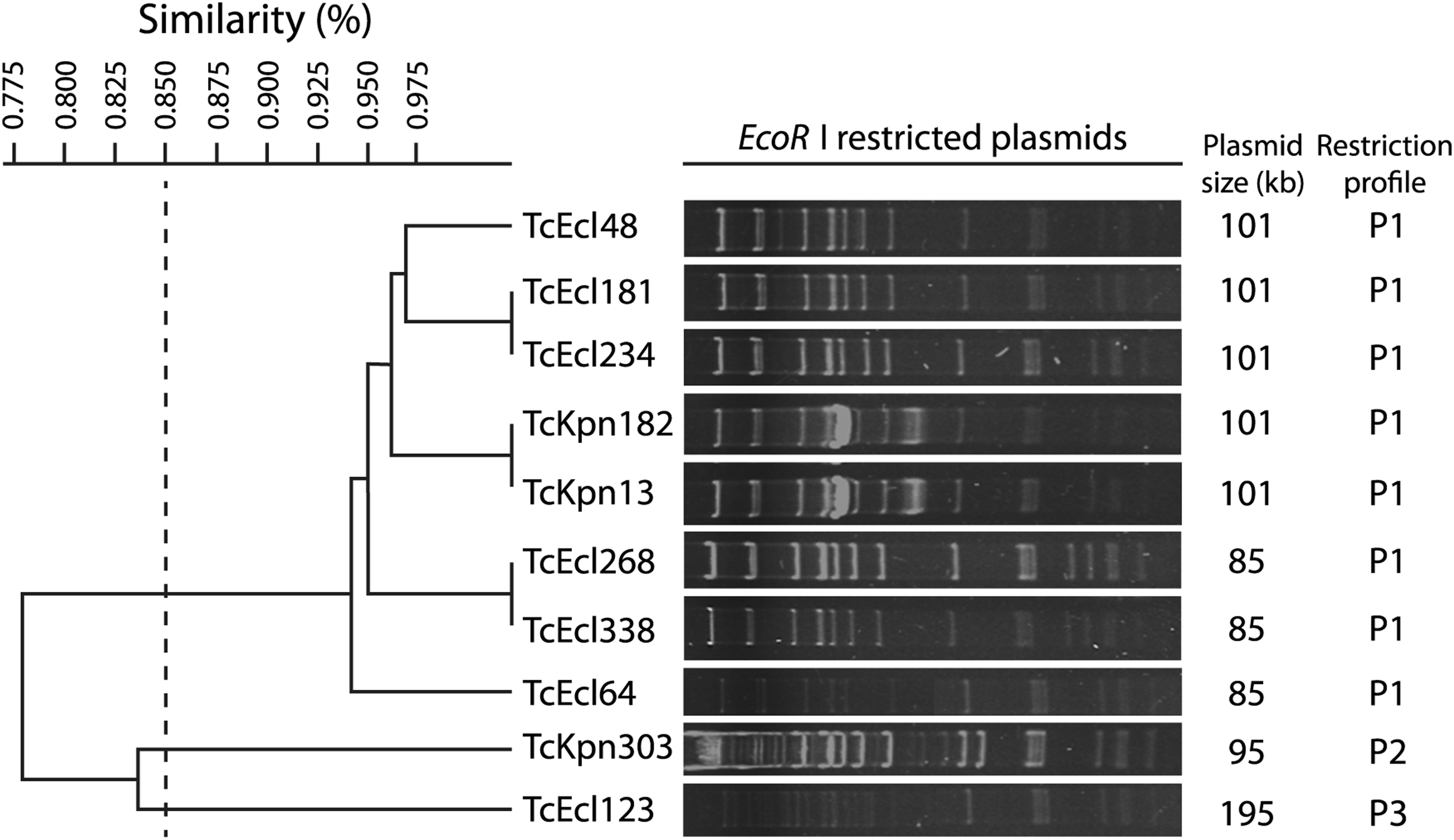
Dendrogram of EcoRI restriction patterns of IncFIA plasmid carrying blaNDM-1 from 10 transconjugants. Three restriction profiles were identified (P1–P3). The dashed line represents the 85% similarity level used in cluster designation. TcKpn, transconjugant of K. pneumoniae; TcEcl, transconjugant of E. cloacae.
Genotyping
PFGE was performed with blaNDM-1-positive strains. Isolates 13 and 182 of K. pneumoniae belonged to clone K1 (Fig. 3) and were isolated within a 5-month period from the general surgery and orthopedic wards, respectively (Table 4). Strain 303 of K. pneumoniae belonged to clone K2, which was obtained from the ICU. In the case of E. cloacae, five strains that belonged to clone E1 were isolated during a period of 9 months in the ICU, general surgery and orthopedic wards (Fig. 3 and Table 4).

Dendrogram represents the Dice similarity index of PFGE XbaI profiles for
Discussion
The emergence and global spread of multidrug-resistant Gram-negative bacteria constitute a serious threat to global public health. In particular, carbapenem-resistant Enterobacteriaceae isolates have been associated mainly with increased morbidity and mortality. 27 Transmissible carbapenem resistance in Enterobacteriaceae has been recognized for the last two decades and has recently been associated with NDM-type MBL. 27
Previous studies have identified blaNDM-1 among clinical isolates of multidrug-resistant Enterobacteriaceae in Mexico City and in three states from distinct geographic regions in Mexico (Nuevo León, Jalisco, and Morelos).8–13 In this study, we report the identification of NDM-1 in three K. pneumoniae isolates and seven E. cloacae isolates that caused nosocomial infections from patients in a Mexico City hospital. These isolates carried only carbapenemase-type blaNDM-1 and did not contain the carbapenemase genes, blaIMP, blaVIM, blaKPC, and blaOXA-40-like. Although the presence of 19 NDM variants has been previously identified in different regions of the world,5,6 in Mexico, only NDM-1-producing bacteria have been reported.8–13
The antimicrobial susceptibility results of the first K. pneumoniae isolate carrying the blaNDM-1 gene are indicative of multidrug resistance since it showed resistance to all β-lactams, aminoglycosides, nitrofurantoin, trimethoprim/sulfamethoxazole, and colistin and was susceptible only to tigecycline and ciprofloxacin (Table 3). The two isolates that were subsequently isolated were pandrug resistant. This pattern of pandrug resistance in Mexico has been recently reported by the SENTRY Antimicrobial Surveillance Program 28 and by Bocanegra-Ibarias. 11 Colistin resistance observed in this study is in keeping with the increased use of colistin for management of carbapenem-resistant, Gram-negative bacterial infections, which have been reported in several countries worldwide, including Mexico where two studies have reported the emergence of colistin-resistant K. pneumoniae.11,29,30 The mutations in chromosomally mediated, two-component regulatory systems 17 must be responsible for expression of the colistin resistance phenotype; for this reason, blaNDM-1-positive colistin-resistant K. pneumoniae does not carry the mcr-1 gene, which is responsible for colistin resistance mediated by plasmids.
Transconjugants of K. pneumoniae acquired resistance to β-lactams and aminoglycosides, which indicates that the conjugative plasmids carrying blaNDM-1 must carry genes of acetyltransferases (AAC), phosphotransferases (APH), or nucleotidyl transferases (ANT) that confer resistance to aminoglycosides and have been reported to be encoded in plasmids.31,32 These results strongly suggest that resistance to the rest of the antibiotics, including resistance to colistin, is encoded in chromosomes. 17 We observed resistance to β-lactams and aminoglycosides in five of the seven E. cloacae transconjugants and only to β-lactam in the other two isolates, which suggests that resistance to aminoglycosides could be localized at the chromosomes associated with methyltransferase ArmA or RmtB.32,33
We found it noteworthy that K. pneumoniae and E. cloacae were harboring two to three large plasmids with great plasticity (one of them carrying blaNDM-1), which allowed them to be transferred by conjugation. In Mexico, the genetic transfer of NDM-1 in K. pneumoniae and E. cloacae has been reported in conjugative plasmids from 101 to 150 kb.9–11 In the present study, the transfer of NDM-1 was also carried out in plasmids of similar size; 95 and 101 kb in K. pneumoniae and 85 and 195 kb in E. cloacae.
Restriction analysis revealed that plasmids of 101 kb in two isolates of K. pneumoniae (13 and 182) and three E. cloacae isolates (48, 181, and 234) and the 85 kb plasmids in three E. cloacae isolates (64, 268, and 338) are strongly related since they showed the same restriction pattern P1, which can be considered as a single epidemic plasmid involved in the NDM-1 gene dissemination between K. pneumoniae and E. cloacae isolates. It is important to note that all these P1 plasmids as well as the plasmids with restriction patterns P2 and P3 belonged to the same incompatibility group IncFIA. IncFIA is a new plasmid incompatibility group identified among NDM-1-carrying Enterobacteriaceae in Mexico.8–11
The temporal analysis on detection and distribution of the three NDM-1-producing K. pneumoniae isolates identified one of each isolate in a different hospital ward during a 10-month period. The first isolate (Kpn 13) was detected in February 2015 and the second (Kpn 182) in July 2015 and both belonged to the same clone K1. The antimicrobial susceptibility analysis showed that the first isolate was susceptible to ciprofloxacin and tigecycline, while the second was resistant to both antibiotics (Table 3). These results suggest that during the time that the bacteria persisted in the hospital, mechanisms of chromosomal resistance (mutations in gyrA and parC that confer resistance to quinolones34,35 were activated and the mutation in the ramR repressor of the acrAB efflux pump that confers tigecycline resistance 36 ) occurred, which allowed it to become a pandrug-resistant strain. In addition, these two isolates harbored a similar conjugative plasmid P1.
With regard to NDM-1-producing E. cloacae isolates, the first isolate was detected in March 2015, 1 month after identifying the first isolation of NDM-1-producing K. pneumoniae. Five of the seven E. cloacae isolates corresponding to clone E1 were recovered from the ICU (one isolate), orthopedic service (one isolate), and surgical ward (three isolates), these last ones during a period of 9 months (Table 4). E. cloacae isolates 48, 181, and 234 belonged to the same E1 clone, carried a conjugative plasmid of similar size (101 kb), and had a similar enzyme restriction pattern P1. Although isolates 64 and 268 belonged to clone E1 and isolate 338 to clone E3, all harbored a conjugative plasmid of 85 kb, which showed a restriction pattern similar to P1. Isolate 123 was the only one of clone E2 in addition to carrying a large 195 kb conjugative plasmid with a P3 restriction pattern different from that of the other plasmids. These results indicate that (i) these clones can survive for long periods of time in the hospital environment, allowing the horizontal transfer of blaNDM-1 to other bacteria that act as efficient donors and recipients, 37 and (ii) the transfer of blaNDM-1 is carried out using similar or different plasmids.
The similarity between plasmids of 101 kb in the two isolates of K. pneumoniae 13 and 182 is consistent since both isolates also belong to the similar clone. Likewise, five E. cloacae isolates belonging to the same clone E1 had the identical restriction pattern of plasmid P1, three of them showed the same plasmid size of 101 kb, and two other isolates harbored a plasmid of 85 kb.
In our study, it was not possible to establish the direct epidemiological relationship between the patient–patient contacts to explain the direct transmission of NDM-1 carrier isolates, nor was a reservoir detected that could allow persistence of these strains for a long time in the hospital environment. However, we definitely documented the emergence, persistence, and dissemination of blaNDM-1 carried in IncFIA plasmids among K. pneumoniae and E. cloacae isolates in this hospital environment in a relatively short period of time. These observations are of great concern not only because of the therapeutic implications of pandrug-resistant bacteria but also because they indicate a growing dispersion of these plasmids in Mexico. They also underscore the urgent need for implementing closer epidemiological surveillance and preventive measures among health personnel in this hospital to prevent dissemination of pandrug-resistant bacteria harboring blaNDM-1.
Footnotes
Acknowledgments
This work was supported by Consejo Nacional de Ciencia y Tecnología, México, grant number PDCPN 247489 and Programa de Apoyo a Proyectos de Investigación e Innovación Tecnológica (PAPIIT) grant number IN221617 to María Dolores Alcántar Curiel. The authors are grateful to Ing. Omar Agni García from the Universidad Nacional Autónoma de México, Ciudad de México, for assistance with the graphic design.
Disclosure Statement
No competing financial interests exist.