
Abstract
Antibiotic resistance is one of the greatest challenges in the health system nowadays, representing a serious problem for public health. Initially, antibiotic-resistant strains were restricted to the hospital environment, but now they can be found everywhere. Globalization, excessive use of antibiotics in animal husbandry and aquaculture, use of multiple broad-spectrum agents, and lack of good antimicrobial stewardship can be listed as the factors most responsible for the spread of antibiotic resistance. The increase in the prevalence of antibiotic-resistant pathogens implies having fewer antimicrobial agents to treat infections. The estimate is that by 2050, there will be no effective antibiotic available, if no new drug is developed or discovered. This raises the need to search for alternative methods of controlling antibiotic-resistant pathogens. Considering this problem, the objective of this review is to outline the most frequent antibiotic-resistant bacteria and describe the advantageous and limitations of alternative methods that have been proposed to control them.
Introduction
One of the greatest challenges in the health system nowadays is the rise of pathogenic species resistant to antibiotics. Infections caused by multidrug-resistant (MDR) bacteria are increasingly common and represent a serious problem for public health. Initially, these species were restricted to the hospital environment, but now they can be found everywhere. Among the gram-positive species, Staphylococcus aureus, Enterococcus faecium, Enterococcus faecalis, and Streptococcus pneumoniae are the most frequent problem. 1 Among the gram-negative strains, Escherichia coli, Klebsiella pneumoniae, Pseudomonas aeruginosa, and Acinetobacter baumannii have been most common.2,3
Antimicrobial resistance in these strains is genetically determined and most commonly mediated by the acquisition of extra-chromosomal genetic elements via horizontal gene transfer. 4 Low permeability of the outer membrane in gram-negative bacteria, efflux pumps, production of degrading enzymes, and modification of targets are examples of mechanisms used by bacteria to resist the toxicity of antibiotics. Globalization, excessive use of antibiotics in animal husbandry and aquaculture, use of multiple broad-spectrum agents, and lack of good antimicrobial stewardship can be listed as the factors most responsible for the spread of antibiotic resistance species. 2
The increase in the prevalence of antibiotic-resistant pathogens implies having fewer antimicrobial agents to treat infections caused by these bacteria. 5 The estimate is that by 2050, there will be no effective antibiotic available to treat infections if no new drug is developed or discovered. 6 This raises the need to search for alternative methods of controlling antibiotic-resistant pathogens, and there are in fact many research groups around the world actively looking for new solutions. The objective of this review is to list the most frequent antibiotic-resistant bacteria and describe alternative methods that have been proposed to control them.
Common Antibiotic-Resistant Bacterial Species
Health care-associated infections raise morbidity and mortality rates worldwide. The increase in mortality is directly related to antimicrobial resistance which makes antibiotic therapy more restrictive, thus also making it difficult to treat infections caused by multiresistant microorganisms. At the beginning of the 21st century, infections with carbapenem-resistant gram-negative bacilli, mainly Enterobacteria, became a big public health problem. 7 MDR gram-negative bacteria including A. baumannii, Pseudomonas aerugionosa, extended-spectrum beta-lactamase (ESBL)-producing Enterobacteria, and carbapenem-resistant Enterobacteria (CRE) are considered the main causative agents of nosocomial infections. 8 Methicillin-resistant S. aureus (MRSA) and vancomycin-resistant Enterococcus (VRE) have been reported recently to be the most common bacterial pathogens, and besides, hospitals have also been isolated from foods of animal origins, water, and animals (Tables 1 and 2). MDR P. aeruginosa, Carbapenem-resistant Enterobacteriaceae, and A. baumannii have been mainly associated with clinical samples, but some strains have also been isolated from foods, animals, and water (Tables 3–5).
Methicillin-Resistant Staphylococcus aureus Isolated from Different Sources and Countries in the World
AmongStaphylococcus aureus strains.
—, Not reported; MRSA, methicillin-resistant S. aureus; CA-MRSA, community-associated MRSA; LA-MRSA, livestock-associated MRSA.
Vancomicin-Resistant Enterococcus Isolated from Different Sources and Countries in the World
Among Enterococcus strains.
Carbapenem-Resistant Enterobacteriaceae Isolated from Different Sources and Countries in the World
Carbapenem-Resistant Acinetobacter Baumannii Isolated from Different Sources and Countries in the World
Among A. baumannii strains.
Multidrug-Resistant Pseudomonas aeruginosa Isolated from Different Sources and Countries in the World
Among P. aeruginosa strains.
All these strains have been associated with lethal infections, and in this section, their main characteristics will be addressed.
Methicillin-resistant S. aureus
S. aureus is a gram-positive ubiquitous strain known to produce several virulence factors that facilitate disease causation and help rapidly develop antimicrobial resistance against antimicrobial agents used for its control, a feature that increases the importance of this microorganism as a pathogen. 9 Initially, infections caused by S. aureus were easily controlled by penicillin. However, S. aureus acquired a plasmid-encoded beta-lactamase that conferred resistance to penicillin already in the 1940s, shortly after its introduction for clinical use. To solve this problem, a new, semisynthetic, narrow spectrum, beta-lactamase-resistant antibiotic named methicillin was developed and introduced to combat penicillin-resistant strains in 1959. Unfortunately, as evidenced by current scenarios, methicillin-resistant strains arose quickly. 4
Since its emergence in 1961, MRSA has spread worldwide, and infections caused by this microorganism are regarded as one of three major infectious diseases threatening human health. This bacterium apart from causing infections in cutaneous lesions can result in severe cases of pneumonia, meningitis, endocarditis, septicemia, and even systemic infections, with risk of death. 10 Compared to infections caused by S. aureus strains sensitive to methicillin, those caused by MRSA usually have more severe clinical manifestations and are the most difficult to treat, as methicillin resistance indirectly affects other virulence factors and enhances the pathogenesis of the bacterium. 11
According to the data published by the Center for Disease Control and Prevention (CDC) of the United States, more than 80,000 illnesses and 11,000 deaths in the hospital setting were caused by MRSA during 2011. 12 The European Centre for Disease Prevention and Control also published in 2013 that S. aureus is among the most frequently isolated pathogen from health care-associated infections. Togneri et al. 10 made a retrospective review for 12 years (2002–2013) of infections caused by S. aureus in adult patients (AP) and in pediatric patients (PP) visiting the Hospital Interzonal General of Treble Evita de Lanús in Argentina and concluded that methicillin resistance increased from 28% to 78% in AP and stayed around 50% in PP. Chmielarczyk et al. 13 isolated MRSA strains from patients in 12 different hospitals in southern Poland and the prevalence of MRSA was higher in patients aged 80 years and more. These results are just a few examples that show how common and persistent MRSA strains are worldwide.
Initially, MRSA was restricted to hospital environment (HA-MRSA: health care-associated MRSA), but rapidly, since the 1990s, this microorganism has also been found associated with community settings (community associated MRSA [CA-MRSA]) and has been isolated from the most diverse sources worldwide (Table 1). Apart from HA-MRSA and CA-MRSA, livestock-associated MRSA (LA-MRSA) have been isolated from several animals such as cattle, pork, chicken, and horse, among others and associated with zoonotic pneumonia, endocarditis, and necrotizing fasciitis since 1975.10,14 The fact that MRSA has been frequently isolated from livestock animals has been worrying, as it has revealed an expanded reservoir of MRSA. 15 Consequently, MRSA has also been isolated from animal based food products, including meat and milk products, such as cheese, 16 making it also an important food-borne microorganism. 17
The first case of LA-MRSA from a human source was described in 2005, and van Loo et al. 18 observed that LA-MRSA strains in human population were responsible for >20% of all MRSA in the Netherlands. Since then, transmission of MRSA from animals to humans and vice-versa has been constantly described. 19 These findings show that some S. aureus strains might not be strongly host species-specific, and LA-MRSA can cause infection in humans. 20 It is believed that some LA-MRSA lineages have derived from human strains following genetic adaptation.
Molecular and microbiological studies have demonstrated that HA-MRSA and CA-MRSA strains have distinct genetic and phenotypic profiles.21,22 HA-MRSA is considered an opportunistic pathogen and CA-MRSA is capable of causing infections in healthy people. Since its inception, CA-MRSA has become the most significant pathogen in several parts of the world and its prevalence has surpassed the cases acquired in hospital. 10 Moreover, CA-MRSA is also infiltrating health care settings and infecting patients in many regions worldwide. There is little evidence of the spread of HA-MRSA outside the hospital setting 23 although a HA-MRSA variant strain causing infections in the community has been reported. 24
As is well established by several authors, methicillin resistance in S. aureus is mediated by a penicillin-binding protein (PBP2A) encoded by the mecA gene, that is carried on a mobile genetic element designated staphylococcal cassette chromosome mec (SCCmec). Moreover, methicillin resistance determinants are different among HA-MRSA and CA-MRSA. HA-MRSA strains present a relatively large mobile genetic element known as SCCmec types I, II, or III, which confers resistance to many classes of non-beta-lactam antibiotics. In CA-MRSA, a smaller element, including SCCmec IV and V, confers resistance, and these strains are susceptible to many non-beta-lactam antibiotics. 21 In addition, a large numbers of CA-MRSA strains produce the Panton Valentine Leukocidin, potent virulence factor that has greater cytotoxic activity against polymorphonuclear cells. 25
Vancomycin (VAM) and daptomycin (DAP) are often the antibiotics of last resort for many S. aureus infections. 26 However, MRSA strains have shown a MDR phenotype, and the most worrying problem is the appearance of strains exhibiting resistance or reduced susceptibility to these antibiotics. Three classes of vancomycin-resistant S. aureus that differ in vancomycin susceptibilities have emerged: vancomycin-intermediate S. aureus (VISA), heterogeneous VISA (hVISA), and high-level vancomycin-resistant S. aureus (VRSA). 27
VRSA species arose due to the acquisition of the vanA gene from vancomycin-resistant enterococci and have been reported in the United States, 28 Iran, 29 and India. 30 The isolation of VISA and hVISA seems to be a serious emerging problem as they show high spreading potential. After the first reports of VISA and hVISA from Japan, 31 it did not take long for this resistance phenotype to be recognized around the world. 27 The in vivo development of VISA and hVISA has led to treatment failures and prolonged hospitalization. 32
DAP is bactericidal against MRSA and VISA strains and was used to treat skin/soft-tissue infections caused by MRSA in 2003 in the United States. 33 Syrogiannopoulos et al. 34 showed that DAP alone or in combination with other antimicrobial agents was efficacious to control MRSA infections in children. However, studies are showing that clinical S. aureus strains have developed DAP resistance during DAP treatment. 35 These findings raise the need to research for alternative methods for the control of MRSA.
Vancomycin-resistant Enterococcus
Enterococci are indigenous flora of the gastrointestinal tracts of animals and humans, and the species E. faecium and E. faecalis have heightened interest because of their ability to cause serious infections and their intrinsic resistance to antimicrobials, including Vancomycin. 36 VRE faecium (VREfm) has disseminated rapidly in hospitals in many parts of the world since 2012. In contrast, vancomycin resistance has been reported considerably less frequently in E. faecalis globally. 37
VRE was first isolated in Europe in 1986 from clinical samples. One year later, the first VRE isolates were identified in the United States. In Europe, the cases were related to community infections, transmitted by animal-based food products to humans, and associated with the use of Avorpacina (an antimicrobial of the group of glycopeptides) as a growth promoter in livestock. In the United States, the predominance of VRE was observed in hospital setting which spread rapidly in the 1990s, probably due to the use of vancomycin. A similar situation occurred in Europe, where the spread of hospital-related VRE occurred a decade later in the 2000. 38 VRE has been an emerging problem in some Brazilian hospitals since the 1990s. The first report of a VRE infection in Brazil occurred in 1996 in Curitiba. Since 1997, the occurrence of outbreaks has been documented in hospitals from other Brazilian cities as well. 39
VRE was described as “serious” (the second highest threat level among organisms causing severe human infections) by CDC. Among hospitalized patients, over 20,000 experience an infection due to VRE each year and it is estimated that 1,300 deaths occur directly due to these resistant infections in the United States. 12 VRE is a common cause of nosocomial infections and has also associated with urinary tract infections, hospital-acquired bloodstream infections, endocarditis, abdominal and pelvic abscesses, and chronic periodontitis. 40
Resistance to vancomycin is determined by one of nine resistance determinants (vanA, B, C, D, E, G, L, M, and N), but the vanA and vanB genotypes predominate worldwide. These genetic determinants could be carried in the mobile genetic element, such as Tn1546, mostly located on conjugative plasmids (variants of the vanA and vanB-type) or located on the chromosome (vanC). 41 The first case of VRE transmitting the vanA gene to MRSA (VRSA) was reported in the United States in 2002, increasing the threat of increased colonization and infections by VRE. 42
Linezolid is the first-line drug for treatment of VRE infections, but the first reports of Linezolide-resistant VRE have appeared, reducing treatment options. 38 Some authors have also suggested daptomycin as an effective agent in the treatment of VRE infections. 43
Carbapenem-resistant enterobacteria
The emergence of CRE is one of the major public health problems in the world. Carbapenems are important for the empirical treatment of critically ill patients at the risk of multiresistant bacterial infection. 44 They have been used as a drug of choice for the treatment of infections caused by ESBL producing enterobacteriaceae for years, which is also one of the main factors responsible for the emergence of CREs through selective pressure. 44
Among the Enterobacteriacea, K. pneumoniae is the most common bacterium exhibiting carbapenem resistance followed by Enterobacter species. Others such as E. coli have been reported less frequently. CREs have emerged in recent decades, but have become one of the major concerns of hospital infection control services. High prevalence of infections by these bacteria is present in several countries on all continents, leading to an important restriction in treatment options. 45 It is considered endemic in the north-eastern United States (mainly in the state of New York), Puerto Rico, Colombia, Greece, Italy, Israel, China, and Brazil, and is also an important cause of nosocomial infections in these countries. 46
Early reports of CREs were related to overexpression of ampC, and ESBL associated with loss or modifications of porins. However, later they were confirmed to produce a new type of enzymes (carbapenemases) with the capacity to inactivate any type of beta-lactam, including the carbapenems. 47
The first carbapenemase was identified more than 20 years ago in 1982, in Enterobacter cloacae, at a hospital in London, England. 48 Subsequently, Serratia marcescens enzymes (SME) were described in S. marcescens. 49 In 1995, in Japan, blaIMP-1 gene was described in a plasmid in Serratia. In 2001, in the United States, first blaKPC-1 gene was identified in K. Pneumonia. 50 Since then, the number of new carbapenemases have increased and proliferated around the world, generating one of the major therapeutic and epidemiological concerns. 51
According to the classification proposed by Ambler, the betalactamases are divided into four groups (A–D) with respect to their functional and molecular properties. Groups A, C, and D are serine enzymes, whereas group B are metalloenzymes. Carbapenemases belong mainly to three major groups of betalactamases A, B, and D, with differences not only in their genetic and biochemical properties but also in their clinical characteristics due to difference in their antimicrobial resistance and epidemiological features. 52
Carbapenemases represent the most versatile enzymes in the beta-lactamase family. The group of carbapenemases class A includes members of families SME, IMI (Imipenemase), NMC (Non Metalobetalactamase), GES (extended spectrum Guyana betalactamase), and K. pneumoniae Carbapenemase (KPC). Of these, KPCs are the most predominant, found mainly on plasmids in K. Pneumonia.47,49 Since its initial recognition, KPC has been also detected in many other strains, including Klebsiella oxytoca, Enterobacter spp., E. coli, Salmonella spp., Serratia spp., Citrobacter freundii, Proteus mirabilis, A. baumannii, P. aeruginosa, and Pseudomonas putida. 49
Class D carbapenemases, known as oxacillinases (OXA), consist of OXA derivatives, a family detected in A. baumannii, but already also described in enterobacteria. These beta-lactamases exhibit activity against carbapenems but do not degrade cephalosporins. 47
The metalobetalactamases belong to the families IMP, VIM, SPM, GIM, and SIM and were detected mainly in P. aeruginosa. One of the major threats among this carbapenemase group is New Delhi Metalobetalactamase (NDM), which was discovered in 2008 in Sweden in K. pneumoniae from an Indian patient. NDM has been found to exhibit a rapid distribution in hospitalized patients throughout the world in other enterobacterias as well. 53
CREs are spread across several countries and have emerged as a major threat to hospitalized patients. In the case of infections caused by K. pneumoniae-producing carbapenemases, the mortality rate may be as high as 75%, but the associated factors such as age, comorbidities, functional status, and underlying disease may influence this rate. 54
Carbapenem-resistant A. baumannii
The genus Acinetobacter consists of gram-negative, aerobic cocobacillus that are ubiquitous, immobile, nonfermenting, catalase positive, and oxidase negative. The A. calcoaceticus-A. baumannii complex is responsible for most of the community or hospital-acquired infections. 55
The different species of Acinetobacter present in diverse natural habitats can be isolated from the soil, water, vegetables, and animal and human hosts. They are part of the commensal flora of human skin and mucous membranes. A. baumannii can survive in a variety of settings in the hospital environment: in dialysis machines, mechanical ventilation systems, water sources, skin and mucous membranes of health professionals and patients, medicinal preparations, and disinfectants. 56
For a long time Acinetobacter was considered an opportunistic agent with low pathogenicity. However, the presence of several virulence factors that allow its survival in the hospital environment and enhance its capacity to cause disease have resulted in it being one of the main causes of nosocomial infections in many countries. 57 Recently, the World Health Organization announced this microorganism as the first priority pathogen, for which research and development for new antibiotics are urgently needed. 58
A. baumannii is increasingly implicated for causing health-associated infections, which confer a high risk of morbidity and mortality to patients. This bacterium can also be highly resistant to antimicrobials, especially those isolated from the patients in the intensive care units. Infections caused by MDR A. baumannii strains may worsen patient outcomes due to inadequate initial therapy, limited treatment options, and high toxicity of available therapies. Risk factors for infection and colonization by MDR A. baumannii include prolonged hospitalization especially in intensive care unit, mechanical ventilation, central venous catheterization, urinary catheterization, previous antimicrobial exposure, increased disease severity, and exposure to surgical and invasive procedures. 59 In the United States, A. baumannii is responsible for 12,000 cases of health-associated infections annually, of which 7,200 are multiresistant causing 500 deaths per year. 60
This microorganism has progressively accumulated resistance to penicillins, cephalosporins, quinolones, and aminoglycosides. Consequently, carbapenems have become the therapy of choice for serious infections. The mechanisms of resistance of A. baumannii can be intrinsic or acquired and are mediated by several factors, such as loss of membrane permeability and, more significantly, the production of betalactamases, enzymes that degrade beta-lactam antibiotics. Betalactamases are the most important cause of bacterial resistance, mainly in gram-negative bacilli. 61
Resistance to carbapenems may occur by combining different mechanisms such as change in the affinity to PBPs and efflux pumps. However, the main forms of resistance to carbapenems are the expression of carbapenemases of group B and D of Ambler, metallo-β-lactamases, and OXA, respectively. 61
Multidrug-resistant Pseudomonas
P. aeruginosa is a nonfermenting gram-negative bacillus, widely distributed in nature and in hospital environment. Responsible for nosocomial infections, it is one of the most important opportunistic pathogen causing bloodstream infection, urinary tract infection, and ventilator-associated pneumonia, especially in critically ill patients receiving intensive care. 62 Moreover, it is also highly resistant to many currently used drugs making it a major public health concern. 63
P. aeruginosa is intrinsically resistant to several antimicrobials and has great versatility to acquire new genes that confer resistance to many other drugs. The antibiotic resistance of this bacterium is mainly due to the low cell wall permeability of this microorganism, which restricts the uptake of antibiotics, associated with wide resistance mechanisms, such as efflux pumps and enzymes, which modify or degrade antibiotics and drug targets. 64
Carbapenems are usually part of the first line of therapeutic choice for treatment of Multdrug-resistant Pseudomonas (MDR) P. aeruginosa infections. Resistance to carbapenems occurs mainly due to the impermeability to the drug, loss of porin, and action of efflux pumps, but the production carbapenemases is the most important mechanism. 65
The main carbapenemases expressed by P. aeruginosa are from class B of Ambler, called metallo-ß-lactamases (IMP, VIM, SPM, GIM, NDM, and SIM families). These enzymes confer resistance to carbapenems and are encoded in plasmids and integrons of class 1, which are responsible for their rapid global spread by horizontal transfer. 66
P. aeruginosa resistant to carbapenems (PARC) has become one of the major problems for hospitals. Outbreaks of infection caused by PARC have been reported by several countries, including Brazil. 66 The emergence of antimicrobial resistance is directly related to an increase in the patient's hospital stay, increased hospitalization costs, and an increase in the mortality rate. In PARC infections, mortality may reach 53.6%. 67
Alternative Methods Proposed for the Control of Multiresistant Pathogens: Advantages and Limitations
With the emergence of MDR pathogens, alternative methods for their control have been studied by several research groups. Among these, the use of bacteriocins, essential oils (EOs), antibodies, and phage therapy have been reported as promising by several authors. In addition, research has also been focused on the use of quorum-sensing inhibitors (QSI) and nanotherapy (Fig. 1). Bacteriocins have been proposed since the 1960s, and antibodies and bacteriophages have been present since 1970's. EOs have been studied and proposed to control MDR pathogens since 1990, and more recently (since the year 2000), quorum sensing inhibitors and nanotherapy have also gained status as a promising method for the control of these resistant strains. A brief review about these alternative methods will be made in the next subsections addressing their importance and limitations.
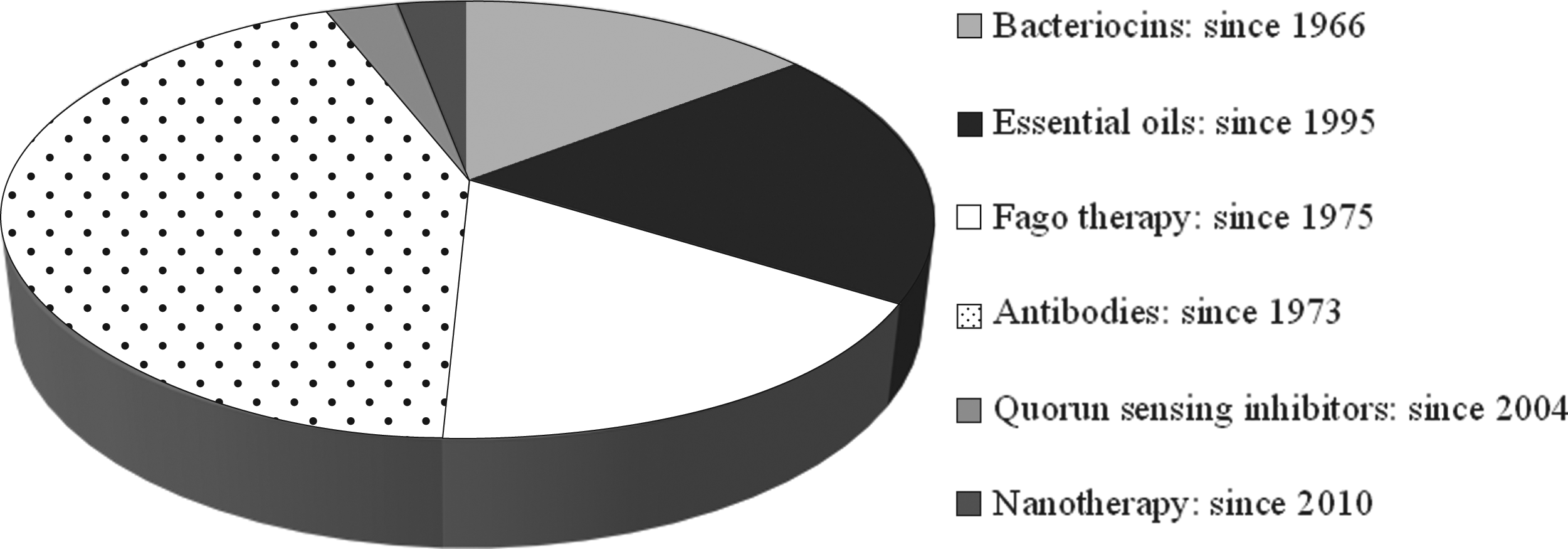
Published articles to date on the use of alternative methods in the control of multidrug-resistant pathogens. Source: PubMed.
Bacteriocins
Bacteriocins are antimicrobial peptides ribosomally synthesized by almost all bacterial species and have a varied mechanism of action and spectrum of activity. A good review on bacteriocin structure, classification, and general characteristics can be found in Snyder and Worobo. 68 The most studied bacteriocins with known industrial applications are those produced by lactic acid bacteria, mainly due to their generally recognized as safe (GRAS) status. Since their discovery, these bacteriocins have been extensively studied for their application in food industry and, more recently, their therapeutic potential has been reported by several authors. Bacteriocins have been proposed as potential anticancer 69 and anticarcinogenic agent 70 and also suggested for the treatment of skin infections 71 and many other diseases. 72
Besides, some authors suggest that bacteriocins may become a potential candidate for replacing antibiotics to treat MDR pathogens in the future. 73 Okuda et al. 74 tested the effect of three different bacteriocins against MRSA biofilms and showed that the bacteriocins formed stable pores on the membrane of target cells and were effective to prevent or cure biofilm-associated infections. Duracin 61A was found to be effective against Clostridium difficile, VRE, and MRSA, showing synergistic effect when used in combination with reuterin or vancomycin. The authors concluded that durancin 61A alone or in combination with other bacteriocins or antibiotics may provide a possible therapeutic option for the treatment of infections by these pathogens. 75
Shokri et al. 76 showed the potential use of a bacteriocin produced by an E. faecium strain against VRE. Phumisantiphong et al. 77 isolated and characterized a novel bacteriocin produced by a strain of Enterococcus faecalis that showed high antibacterial activity against VRE and MDRE (MDR enterococci). The lantibiotic NAI-107 also showed bactericidal activity against MRSA and VRE in animal models. 78 Lacticin 3,147 and nisin were tested against MRSA and VRE strains and also showed potent activity. 79 Wang et al. 80 tested the efficacy of sublancin for the prevention of MRSA-related intraperitoneal infection in mice and verified that the antimicrobial activity involved the destruction of the bacterial cell wall and considerably reduced the mortality and weight loss of MRSA-challenged mice.
Many properties of bacteriocins, such as high stability, low toxicity, and both broad and narrow spectra of activity, make them good alternative to antibiotics. 81 In addition, some bacteriocins, such as lantibiotics, have a dual mechanism of action not shared by other therapeutic compounds in use, which reduces the probability of selecting resistant strains. 82 Unlike most antibiotics, which are secondary metabolites, bacteriocins are ribosomally synthesized, which allows bioengineering to improve its efficiency. Bacteriocins are also sensitive to proteases in the human stomach, not posing any risk of toxicity to the humans or to gut flora. 73
Among the lantibiotics, nisin is the most studied bacteriocin that has a GRAS status and has been approved by Food and Drug Administration (FDA) as food additive since 1988. 83 In the last decades, nisin has also been proposed for therapeutic usage. 84 Studies have shown that nisin can prevent the growth of drug-resistant bacterial strains, such as MRSA, S. pneumoniae, Enterococci, and C. difficile. It is well known that bacteriocins have antimicrobial activities mainly against gram-positive strains. However, many studies have demonstrated that nisin in combination with other antibiotics can also be effective against gram-negative pathogens. 85
As described above, bacteriocins have the potential to control antibiotic-resistant pathogens and are among the best alternative methods proposed for this purpose. However, certain points need to be carefully studied before their clinical use: the first being the bacteriocin resistance in vitro. Resistance to bacteriocin in vitro has been mostly associated with physiological adaptation, unlike antibiotic resistance that is related to genetic modification. 86 Considering that bacteriocins have not yet been used on the same scale as antibiotics, an existing bacteriocin-resistant population does not exist. Thus, before the therapeutic use of bacteriocins, it would be interesting to develop strategies to avoid in the future the current problem with resistance to traditional antibiotics. The best way to use bacteriocins therapeutically could be in combination with traditional antibiotics.
The second challenge involves development of a better way to administer bacteriocin therapeutically, since enzymes present in the gastrointestinal tract inactivate them. Topical, intranasal or intravenous use may be an alternative. Some preliminary studies 87 showed that nisin administered intranasally inhibited the growth of S. aureus in the respiratory tract of immunocompromised rats. Heunis et al. 71 also demonstrated that nisin-containing nanofiber in wound dressings has the potential to treat S. aureus skin infections. Nanoparticles (NPs) could also be used as delivery system for bacteriocins. 88
Essential oils
Another alternative tool that has been frequently discussed by several authors as a solution to control MDR pathogens are EOs extracted from medicinal plants. 89 EOs are defined as volatile, natural, and fragrant liquids that can be extracted from different parts of the plants especially leaves and flowers, and are produced by plants to protect themselves from diverse pathogenic microbes. Due to their antimicrobial activities, EOs have been extensively studied to be used for the treatment of a wide range of microbial infections. 90 EOs have shown antimicrobial activity against MRSA, 91 MDR strains of K. oxytoca, 92 b-lactamases and carbapenemases producing E. Coli, 90 erythromycin-resistant Group A streptococci, 93 and MDR-A. baumannii 89 among others.
EOs have shown potential as inhibitory agents for use in MRSA biofilm-related infections. 94 Lahmar et al. 95 showed that three different EOs were effective in reducing the resistance of MRSA to amoxicillin, tetracycline, piperacillin, ofloxacin, and oxacillin, and resistance of A. baumannii to amoxicillin and to ofloxacin in interactive manner. They also showed synergism with the antibiotics ofloxacin and novobiocin against the ESBL producing E. coli. In another study, the bactericidal activity of herbal volatile oil extract was tested against MDR A. baumannii and exhibited potent antibacterial activity with minimal bactericidal concentration (MBC) values around 0.5. 89 EOs and nonvolatile compounds derived from chamaecyparis obtusa also showed antimicrobial activity against MRSA and VRE strains. 96 Cinnamon bark oil was reported to have antimicrobial activity against clinical isolates of MDR P. aeruginosa with minimum inhibitory concentration (MIC) of 0.0562–0.225% v/v and MBC of 0.1125–1.8% v/v. Synergistic effect was also observed when used in combination with colistin. The authors proposed that this oil could be used as an alternative for the treatment of infections caused by MDR P. aeruginosa. 63
Many other studies are showing the beneficial effects of EOs when two or more EOs are mixed together or when used in combination with commercial antibiotics. 91 When blended with antimicrobial agents, the constituents of EOs can unlock the cell membrane channels, thus opening the passage of antimicrobial agents to reach their internal target sites. 97 This is a great strategy to avoid selection of resistant strains in the future.
Low water-solubility and high vapor pressure are some characteristics that limit the utilization of EOs in commercial applications. 98 Consequently, EOs are difficult to be used in aqueous-based products, and they have a tendency to volatilize over time, thereby reducing their antimicrobial activity. An effective means of overcoming some of these limitations could be the utilization of EOs vapors, currently being studied for decontaminating environment and wound dressings. 99 Moreover, to improve water dispersion and protect EOs from degradation, nanosized formulations emerge as a viable solution. 100
Ghaderi et al. 101 produced nanoemulsion-based delivery systems containing EOs that were effective against gram-positive and gram-negative strains related to upper respiratory tract infections. Cardamom oil loaded chitosan nanoparticles also showed excellent antimicrobial potential against ESBL and MRSA. Rai et al. 97 also suggest that the use of EOs in combination with NPs may exert synergistic antimicrobial activity, leading to the development of novel approach for treatments of MDR pathogens. Besides, EOs could also be incorporated into topical creams and nasal sprays to treat skin wounds and upper respiratory tract infections, respectively. 102
Bacterial strains showing resistance to EOs are not described yet. Moreover, Turchi et al. 103 showed that the exposure of S. aureus strains to a subinhibitory concentration of EOs displayed an increased sensitivity in more than 95% of the cases. These results suggest that resistance to EO's could be difficult to arise. At the same time, EOs have not been used on the same scale as antibiotics, so an existing EO-resistant population does not exist. Therefore, to avoid emergence of resistant strains, similar to bacteriocins, the best way to use EOs in the control of MDR pathogens could be in combination with another antimicrobial agents, such as commercial antibiotics, bacteriocins, or phage therapy.
Phage therapies
Phage, also known as bacteriophage, is a virus that infects bacteria. Their therapeutic potential in medicine to control MDR pathogens is due to their specificity and potency in inducing lethal effects in the host bacterium by cell lysis. 22
Bacteriophage therapy was first utilized in the 1930's, but the discovery of antibiotics, a more comfortable way to control infectious diseases, led to a rapid decline in the interests and investments within this field of research. Although phage therapy was given the GRAS status by the FDA in 2006, it is still not approved for use in humans. The key regulatory barriers continue to prevent its approval by the US FDA and by the European Medicines Agency. 104
The worldwide rise of MDR pathogens and the decrease in research and development of new antibiotics have stimulated interest in phage therapy and several studies have shown its efficacy against antibiotic-resistant pathogens. 105 Phage therapy has been shown to be effective against typhoid fever and S. aureus bacteremia 106 and also against MRSA strains. 107 Rasool et al. 107 showed that the phages exhibit lytic activity against MRSA both in vivo and in vitro experiments in another report, Jennes et al. 108 described that phage therapy was effective in the treatment of colistin-only-sensitive P. aeruginosa septicemia in a patient with acute kidney injury. Cheng et al. 104 showed a high efficiency of a broad host range lytic phage against E. faecalis strains, including vancomycin-resistant strains. Chadha et al. 109 showed the potential of liposome entrapped phage cocktail for treating K. pneumoniae-mediated infections. Gelman et al. 110 tested a combination of bacteriophages and antibiotics against VRE Enterococcus faecalis in a mouse model and concluded that this combination imparts an additional beneficial effect on the treatment success, as a single injection of the bacteriophage cocktail was sufficient to completely reverse the 100% mortality trend caused by VRE.
The use of phage therapy has been proposed via intravenous 106 and oral routes, 111 as liposome-entrapped phage cocktail, 109 and for vaccine development. 112 Some advantages of using phage therapy instead of antibiotic include lower developmental costs; 100% bactericidal nature; high specificity thus also preventing secondary infections; and the requirement of only a single dose or phage multiplication at the infection site compared to antibiotics that require several doses. In addition, phage therapy can be used in combination with traditional antibiotics, or a combination of diverse phages as a cocktail to increase its antibacterial spectrum. 113
The use of whole phage to treat infection could have some disadvantages, as the genetic material in temperate phage could increase the virulence of certain species of bacteria through transduction of virulence genes. An example is the acquisition of the gene encoding the Panton Valentine Leucocidin toxin, causing “scalded skin syndrome,” 114 common in CA-MRSA strains that increase their pathogenicity. In this way, instead of the whole phage, some studies have proposed the use of bacteriophage-encoded endolysins that destroy the bacterial cell wall, resulting in cell death through lysis. 105 Some advantages of using endolysins as pointed by Fischetti et al. 115 include their ability to retain specificity without affecting commensal flora, rapid bacterial lysis, genetically nontransducible nature, and lesser probability of acquiring resistance due to the essential bacterial binding sites. These endolysins have demonstrated antibacterial activity against gram-positive and gram-negative strains and also against MDR pathogens such as MRSA. 116
Quorum-sensing inhibitors
QSI are another alternative method to control MDR pathogens proposed in the last years. Quorum sensing (QS) is an intercellular bacterial communication used to coordinate group behaviours in a cell density-dependent manner. At high concentrations, pathogens can switch their transcription profiles to an invasive phenotype, including genes related to antibiotic tolerance and virulence determinants, and cause disease. 117 In this way, QS systems constitute important antivirulence targets, as they often regulate the expression of several virulence genes simultaneously. 118 QSI act by inhibiting cell-to-cell communications and, consequently, disease evolution, enabling the host immune system to prevent bacterial colonization and/or to clear an established infection. This antimicrobial control relies on reducing the burden of virulence rather than killing the bacteria. 119
Hansen et al. 120 tested the use of lactam hybrid analogs of solonamide B and autoinducing peptides to inhibit the Agr QS system, which is a major inducer of virulence in CA-MRSA. The authors concluded that this and closely related compounds were 20- to 40-fold more potent in AgrC inhibition than the starting hit compound. Baldry et al. 121 also showed that solonamide B and analogs alter immune responses to S. aureus, but do not exhibit adverse effects on immune cell functions. However, the authors also stated that application of compounds inducing an Agr-negative state may have adverse interactions with host factors in favour of host colonization. RNAIII-inhibiting peptide was also appointed by Simonetti et al. 122 as useful for the control of MRSA in infected wounds by inhibiting QS systems.
Hraiech et al. 123 have shown that a lactonase isolated from Sulfolobus solfataricus efficiently inhibited QS in P. aeruginosa, reducing the mortality in rats from 75% to 20% based on early treatment. QS system from this bacterium was also inhibited by bioactive molecules extracted from leaves of Kalanchoe blossfeldiana. 124 D'Angelo et al. 125 tested FDA-approved drugs as antivirulence agents targeting the pqs QS system of P. aeruginosa and found that clofoctol (approved for clinical treatment of pulmonary infections caused by gram-positive bacteria) has considerable clinical potential as an antivirulence agent for the treatment of P. aeruginosa lung infections. Anti-QS system against P. aeruginosa was also observed in a study with EOs derived from Ferula (Ferula asafoetida L.) and Dorema (Dorema aucheri Bioss.), exhibiting anti-QS activity at 25 μg/mL of concentration. 126
In the last 20 years, various QSI from plants, animals, and microorganisms have been characterized and animal and plant infection models have demonstrated their antibacterial efficacy against QS pathogens. 119 QSI could thus serve as a good alternative to treat infections caused by MDR pathogens. However, its application in clinical medicine still requires more research. According to Baldry et al. 121 application of QSI might have adverse interactions with host factors in favour of host colonization.
Antibodies
The use of antibodies has also been proposed as a promising strategy for the containment of MDR bacterial strains, as they can enhance phagocytosis or activation of complement proteins. To maximize antibody action, some authors have proposed the use of more than one isotype, generally a mix of Immunoglobulin M (IgM), Immunoglobulin G (IgG), and Immunoglobulin A (IgA).127,128 A polyclonal preparation enriched with IgM immunoglobulins commercially available is Pentaglobin® (Biotest AG, Dreieich, Germany), which contains 76% IgG, 12% IgA, and 12% IgM (IgGAM). To date, there are at least three other licensed monoclonal antibodies products against infectious targets: Palivizumab, 129 Raxibacumab, 130 and Panobacumab. 131
Several studies have shown the effectiveness of this alternative treatment, which is also one of the most studied methods (Fig. 1). Lu et al. 132 tested Panobacumab, a fully human monoclonal antibody of the IgM/κ isotype directed against the lipopolysaccharide (LPS) O-polysaccharide moiety of P. aeruginosa, which enhances the phagocytosis of this pathogen. The authors concluded that this therapy is safe and is associated with high clinical cure and survival rates in patients developing nosocomial P. aeruginosa infection. Rossmann et al. 128 demonstrated that IgGAM induces in vitro killing of MDR clinical isolates of K. pneumoniae through enhancement of phagocytosis. Results obtained by Giamarellos-Bourboulis et al. 127 also showed that IgGAM was effective as an adjunct to antimicrobial treatment for the management of septic shock caused by MDR gram-negative bacteria. Diago-Navarro et al. 133 observed that anticapsular antibodies promoted extracellular processes killing, complement deposition, deployment of neutrophil extracellular traps, and opsonophagocytosis of carbapenem-resistant Klebsiella pneumonia. The author concluded that the tested antibodies could ultimately treat or protect patients infected or at risk of infection by this MDR bacterium.
Antibody conjugates with antimicrobial agents have also been shown to be effective to control MDR strains by several other authors. Lehar et al. 134 verified that an antibody–antibiotic conjugate eliminates intracellular S. aureus01, and this conjugate was superior to vancomycin for treatment of bacteraemia. Antibody-directed photodynamic therapy was very effective in killing different MRSA strains, in all growth phases and may be a good candidate for a novel treatment of MRSA infections. 135
As stated by Szijártó et al., 136 passive immunization was a standard treatment option in the preantibiotic era, and as we move toward a possible postantibiotic era, it may be prudent to reconsider the merits of this therapy. The actual advances in the research and biopharmaceutical manufacturing capacities enable the development of highly purified humanized antibodies against a range of pathogenic microorganisms. Moreover, MAbs are directed against nonhuman targets and in general have an excellent safety record. Similar to other alternative methods, antibodies can complement antibiotic therapy. A challenge of this therapy is to find molecular determinants of the pathogens that are accessible on the cell surface. 136
Nanotherapeutics
Besides the methods described above, some authors have also proposed the use of nanotechnology in the control of MDR strains. Some of the NPs proposed are silver NPs, 137 Gold NPs, 138 Photoexcited Quantum dots, 139 anf Core–shell Magnetic Nanoparticle (MNP) 140 among others. As stated by Singh et al. 141 “Nanomaterials possess unique properties, and multiple bactericidal mechanisms render them more effective than conventional drugs.” In addition, Zaidi et al. 88 also affirm that NPs have potential to overcome the problem of antibiotic resistance in the present days and “to revolutionize the diagnosis and treatment of bacterial infections.”
The use of NPs can enhance the antimicrobial activity of available drugs by functioning as drug delivery systems that can be targeted precisely. 88 NPs material could be metallic or organic and has been found to synergize the killing effect of antibiotics 142 and could also have the same effect in combination with other antimicrobial agents, such as bacteriocins and EOs.100,105 Generally, the antimicrobial activity of NP involves a triple mechanism of action: oxidative stress, metal ion release, and nonoxidative stress, 143 together with the effect of any other antimicrobial that can be combined with NPs. This multiple mechanism of action makes it more difficult to acquire resistance. Moreover, NPs enhance drug solubility, concurrent delivery of multiple drugs, and prolonged systemic circulation. 144
Yuan et al. 137 used silver NPs to control drug-resistant strains of S. aureus and P. aeruginosa and showed that it induced cell death of these strains. Niemirowicz et al. 139 anslyzed the synergistic effects of core–shell MNPs when used in combination wth cathelicidin LL-37 (a human antibacterial peptide), synthetic ceragenins (CSA-13 and CSA-131), and classical antibiotics (vancomycin and colistin), against MRSA. The authors concluded that MNPs potentialize the effect of these antimicrobial agents and can be considered as a method of delaying and overcoming bacterial drug resistance. Pei et al. 145 developed a polymeric particle formulation for encapsulation and intracellular delivery of vancomycin to MRSA-infected macrophages and showed that this NP enhanced bacterial activity of the antibiotic against MSRA. For in vivo application, the stability of this formulation remains to be improved. 145 Combination of metal oxide NPs with ciprofloxacin, erythromycin, methicillin, and vancomycin was reported to effectively reduce the MICs of these antibiotics against VRE. 146 Khan et al. 147 tested the photo inactivation of MDR strains of E. coli and K. pneumoniae by monomeric methylene blue conjugated gold NPs and obtained 97% killing of MDR bacteria. They put forward the possibility of this NP-based photodynamic therapy as a potential therapeutic approach against MDR infections. Similarly in another report, trimethyl chitosan-capped silver NPs were shown to have high antibacterial activity with a MIC of ≤12.25 μg/mL against clinically isolated MDR A. Baumannii. 148 NPs thus seem to be a good alternative with high potential to solve the emerging bacterial MDR.
Concluding Remarks
Antibiotic resistance is one of the greatest health problems we will have to face in the coming years. Considering that little advancement has been made in the discovery of novel antibiotics especially those effective against drug-resistant strains, use of alternative methods could be the best way to resolve this problem. As discussed in this review, there are several promising alternatives to circumvent this problem. It is up to the competent authorities to accelerate and encourage further research and release appropriate financial support for the successful implementation of these methods. The alternative methods listed in this review probably will be successfully used in combination with the antibiotics available and not completely replace antibiotics as treatment agents.
Footnotes
Acknowledgments
The authors thank the Conselho de Desenvolvimento Cientıfico e Tecnologico (CNPq), Brasılia, Brazil, the Fundação de Apoio a Pesquisa a InovaçãoTecnologica do Estado de Sergipe (FAPITEC), Sergipe, Brazil, for providing fellowships and financial support, which support the research with bacteriocins, essential oils, and antibiotic-resistant pathogens in our laboratory.
This research did not receive any specific grant from funding agencies in the public, commercial, or nonprofit sectors.
Disclosure Statement
No competing financial interests exist.