
Abstract
A total of 161 Streptococcus pneumoniae were collected between 2013 and 2015 in Tehran, Iran. The strains were tested for antimicrobial susceptibility and minimum inhibitory concentrations, serotyped, and genotyped by multilocus sequence typing (MLST) and pulsed-field gel electrophoresis (PFGE). Penicillin-binding proteins (PBPs) were also typed by restriction fragment length polymorphism (PBP-RFLP). Out of 161 strains, 32 isolates (20%) were highly resistant to penicillin. The most frequent serotypes among the penicillin-nonsusceptible S. pneumoniae (PNSP) were 14 (24%), 23F (18%), and 19F (17%). RFLP of pbp2b, pbp2x, and pbp1a genes revealed 8, 6, and 7 different patterns, respectively. Analysis of 93 PNSP isolates displayed 80 PFGE types with 8 common types constituting 21 (23%) isolates. The remaining 72 isolates (77%) were single types. MLST indicated a high degree of genetic diversity among the 93 PNSP with 36 different sequence types. Six internationally known penicillin resistant clones were identified in our isolates among which Spain23F-1 (ST81), Spain6B-2 (ST90), and Spain9V-3 (ST156) were the predominant clones. The results indicated international identifiable clones of S. pneumoniae, especially Spain23F-1 with high penicillin resistance could play a major role in spread of antimicrobial resistance in Iran. The extensive sequence variation in PBP2x, PBP2b, and PBP1a in resistant strains of clinical and commensal S. pneumoniae was suggestive of a widespread homologous recombination within S. pneumoniae populations.
Introduction
S
Emergence of high rate of PNSP, multidrug resistant (MDR) strains, and bacterial serotype variations demands regional and local investigations of S. pneumoniae isolates. The information could be used to decide for a right choice of pneumococcal vaccine in each country which, in turn, benefits medically and economically.
Only a few publications, with limited information, have addressed the genetic diversity of S. pneumoniae in detail in the Middle East region, including Iran. The present report has examined the genetic characteristics of PNSP isolates and suggested the appropriate type of pneumococcal conjugate vaccine (PCV) in reducing the dissemination of S. pneumoniae in Iran.
Materials and Methods
Sample collection and identification
A total of 161 pneumococcal isolates, 88 clinical and 73 normal flora isolates, were collected between 2013 and 2015. Clinical isolates were collected from different hospitals and laboratories in Tehran, Iran. The isolates were obtained from eye (n = 23), blood (n = 19), sputum (n = 13), cerebrospinal fluid (CSF; n = 9), trachea (n = 7), maxillary sinus (n = 6), otitis (n = 4), abscess (n = 2), pleural aspirate (n = 2), urine (n = 2), and bronchoalveolar lavage (BAL; n = 1). The normal flora isolates (73 isolates) were collected from nasopharynx of healthy individuals who did not take any antibiotics for at least 6 months prior sampling. All procedures were done in accordance with the Declaration of Helsinki (1975) and its modification in 2013. 7 Microbiological test for identification of species was performed as described previously. 8 Identification of isolates was also confirmed by lytA and ply genes using species-specific primers. 9
Serotyping
All isolates were serotyped as described before using the Quellung reaction with antisera (Statens Serum Institut Copenhagen, Denmark).
Antibiotic susceptibility tests
Disk diffusion method was done on Mueller-Hinton agar with 5% defibrinated sheep blood according to the Clinical and Laboratory Standards Institute (CLSI) guideline. 10 Antibiotic susceptibility was determined for oxacillin (1 μg), tetracycline (30 μg), chloramphenicol (30 μg), vancomycin (30 μg), and clindamycin (2 μg). Antibiotic discs were purchased from Mast Diagnostics Ltd. (Merseyside, United Kingdom). Minimum inhibitory concentration (MIC) interpretive criteria were used according to CLSI guidelines for penicillin, ceftriaxone, cefotaxime, and amoxicillin using E test (Liofilchem, Italy) on Mueller-Hinton agar according to the manufacturer's instructions. All isolates with MIC value ≥0.12 μg/mL were classified as PNSP. 10 S. pneumoniae strain ATCC 49619 was used as the quality control strain.
Penicillin-binding protein typing
S. pneumoniae penicillin-binding protein (PBP) genes (pbp1a, pbp1b, pbp2a, pbp2b, pbp2x, and pbp3) were amplified using published primers. 11 PCR assay was made in a total volume of 25 μL containing 10 mM Tris-HCl (pH 8.3), 1.5 mM MgCl2, 0.2 mM each dNTPs, 0.5 U of Taq DNA polymerase (HT Biotechnology, Cambridge, United Kingdom), and forward and reverse primer 40 pmol. PCR was done under the following conditions: an initial cycle at 95°C for 5 min; followed by 30 cycles of denaturation at 94°C for 1 min, annealing at 57°C for 1 min, and extension at 72°C for 1 min; with final extension at 72°C for 10 min. 9 Sequencing of the products was carried out using the ABI capillary system (Macrogen Research, Seoul, South Korea) after purification. Addition of primers to amplify an additional fragment of the pbp1a, AAMPF (5′ ACCTACTCACAAC TGGGATGGATG), and AAMPR (5′TGGTTGTGCTGGTTGAGGATTCTG) was used as previously described. 12 For restriction fragment length polymorphism (RFLP), the PCR products were digested separately with 10U HinfI (pbp2x, pbp1a) and HaeIII (pbp2b), according to the manufacturer's instruction. 11 All restriction enzymes were purchased from Roche Molecular Biochemicals (Mannheim, Germany). RFLP-PCR fragments were scanned, and different patterns were compared using BioNumerics software (Applied Maths, Sint-Martens Latem, Belgium) using the Dice coefficient of similarity and cluster analysis with the unweighted pair group method with arithmetic averages (UPGMA).
Pulsed-field gel electrophoresis
Pulsed-field gel electrophoresis (PFGE) method was performed as described before. 13 Briefly, genomic DNA was digested with SmaI, and electrophoresis was done with ramped pulse times beginning with 5 sec and ending with 20 sec at 6 V/cm for 23 hr. The banding patterns were read by Dice analysis and clustered by the UPGMA.
Multilocus sequence typing analysis
Multilocus sequence typing (MLST) was performed using seven gene targets, including aroE, gdh, gki, recP, spi, xpt, and ddl. 14 Among the 90 isolates studied, 4 isolates failed to show any MLST patterns. The amplified products were analyzed by PubMLST site (www.pubmlst.org). A sequence type (ST) was assigned to each unique combination of seven allele numbers. The relationship between ST and the isolate was defined using eBURST version 3 (http://eburst.mlst.net). Clonal complexes, groups of ST sharing six or seven identical housekeeping alleles, were analyzed and compared with the clones reported in the Pneumococcal Molecular Epidemiology Network (PMEN; www.pneumogen.net/pmen).
Statistical analysis
The statistical analysis was performed using SPSS version 20.0 (IBM Corp.). Differences in the proportions among categorical variables were assessed using Fisher's exact test or the χ 2 test as appropriate.
Results
Serotype distribution among pneumococcal isolates
Out of 161 isolates, 88 (55%) isolates were collected from patients in 10 hospitals and 73 (45%) normal flora from healthy individuals in Tehran. Among the pneumococcal isolates, 150 (93%) were serotyped. The serotypes were 14 (17%), 23F and 3 (15% each), 19F (13%), 9V and 19A (9% each), 6A and 11A (6% each), 6B (3%), 18C and 4 (2% each), and 6C, 7F, 33C, and 1 (1 isolate each). The remaining 11 isolates were categorized as nontypable. Vaccine coverage rate for 150 typable isolates for PCV7, PCV10, PCV13, and PPV23 was 62, 63, 93, and 93%, respectively. The two serotypes found in this study, 6C and 33C, are not included in any form of PCV vaccines.
Antibiotic susceptibility
The PNSP was detected in 58% of our isolates (57 clinical and 36 normal flora isolates). The highest rate of antibiotic resistance was 94% for trimethoprim–sulfamethoxazole followed by tetracycline (75%), erythromycin (60%), chloramphenicol (59%), and clindamycin (52%). The rate of penicillin-resistant S. pneumoniae (PRSP) isolates was 15% and 89% in nonmeningitis and meningitis cases, respectively. No significant difference was observed in the rate of MDR isolates between the meningitis and nonmeningitis cases. Among the 152 nonmeningitis isolates, 67 (44%), 61 (40%), and 24 (16%) were susceptible, intermediate, and resistant to penicillin, respectively. Moreover, out of 9 isolates obtained from meningitis, 8 (88%) were resistant to penicillin belonging to serotype 14 (Table 1). Overall, the serotype 14 PNSP isolates were more significantly found in the meningitis than in nonmeningitis cases. In contrast, serotypes 23F, 19F, 19A, and 9V were significantly detected in the nonmeningitis cases (Table 1).
Association Between Serotypes and Susceptibility to Penicillin in Streptococcus pneumoniae Isolates with Respect to the Site of Isolation (Meningitis vs. Nonmeningitis)
The isolates with <5% of the cases were not included.
Indicates the percentage of isolates within each column (category).
Indicates statistically significant by comparing meningitis and nonmeningitis cases for each serotype (p ≤ 0.05).
I, intermediate; N, number of isolates; R, resistant to penicillin; S, serotype.
PNSP characterization
Out of 88 clinical cases and 73 normal flora isolates, 57 (65%) and 36 (49%) were PNSP (p ≤ 0.05), respectively. The source of clinical PNSP isolates (n = 57) was from eye infection (n = 15, 26%), blood (n = 13, 23%), sputum (n = 8, 14%), CSF (n = 8, 14%), sinus (n = 4, 7%), trachea (n = 3, 6%), pleural aspirate (n = 2, 3%), otitis (n = 2, 3%), abscess (n = 1, 2%), and BAL (n = 1, 2%). Out of nine meningitis isolates, eight showed MIC ≥0.12, which were considered as PRSP. Among the 21 invasive and 36 noninvasive isolates, 15 (15/21, 71%) and 12 (12/36, 33%) isolates were PRSP, respectively. The prevalence of PRSP isolates was more significant in the clinical isolates than normal flora (27 clinical compared to 5 PRSP normal flora isolates) (p < 0.05). Serotype 14 was the predominant serotype in CSF (63%) and blood (54%) among the PNSP specimens. Serotypes 14 (22%) and 23F (22%) were predominant in 36 PNSP normal flora isolates.
Serotype distribution in PNSP isolates
Twelve serotypes were found among 93 PNSP isolates with 3 nontypable isolates. Serotype 14 (26%) was the predominant serotype, followed by 23F (19%), 19F (18%), 19A (9%), 9V (7%), 11A (6%), 6A (4%), and 6B and 18C (3% each). The three serotypes that were identified in ≤2% of the isolates were 3 (2%), 4 (2%), and 7F (1%). The vaccine coverage rates of 90 typable PNSP isolates by PCV7, PCV10, PCV13, and PPV23 were 78%, 79%, 95%, and 95%, respectively.
Antimicrobial susceptibility and serotype association in PNSP isolates
Among the 161 isolates, 5 serotypes (18C, 14, 19F, 6B, 23F) significantly exhibited higher level of resistance to penicillin than the other serotypes (p < 0.05). Serotype 18C exhibited the highest resistance (100%, 3/3) followed by 14 (88%, 23/26), 19F (80%, 16/20), and 23F (74%, 17/23). The rate of MDR (resistant to 3 different classes of antibiotics) among 161 isolates was 66%. Among the PRSP and penicillin intermediate susceptible pneumococci (PISP) strains, 91% (29/32) and 66% (40/61) of the isolates were MDR, respectively. Vaccine coverage for MDR-PNSP isolates by PCV7, PCV10, PCV13, and PCV23 was 81%, 81%, 91%, and 87.5%, respectively. The rates of coverage of MDR-PISP isolates by the same vaccines were 46%, 46%, 61%, and 59%, respectively.
Serotype distribution among 32 PRSP isolates was 14 (38%), 19F (22%), 23F (16%), 18C and 19A (6% each), and 6B, 6A, and 9V (3% each). One isolate was nontypable (3%). In addition, distribution of serotypes among 61 PISP isolates was as follows: 23F (20%), 14 (18%), 19F (15%), 19A (10%), 11A (8%), 9V (8%), 6A (5%), 3 and 6B (3% each), and 18C, 7F, and 4 (2% each). Two isolates were nontypable (4%). Serotype 14 was the most prevalent serotype in PNSP clinical (26%) and normal flora isolates (22%). Serotype distribution was not statistically different between the clinical and normal flora isolates (p > 0.05). Among the clinical isolates, serotype 14 (12/21, 57%) was the most prevalent serotype in the invasive isolates. In contrast, serotype 19F (10/36, 28%) was the most prevalent serotype in the noninvasive isolates.
Genotyping of PBP genes by RFLP analysis of PNSP isolates
Digested PBP-PCR products resulted in 8, 6, and 7 profiles for pbp2b, pbp2x, and pbp1a, respectively. For pbp2b, the largest group (profile B1) was composed of 32 isolates followed by profiles B2 and B3 with 18 and 15 isolates, respectively. For pbp2b, the most prevalent pbp profile in PISP and PRSP was profile B1 with 31% (19/61) and 34% (11/32) isolates, respectively. Serotype 14 was the most prevalent serotype in the profiles B1, B2, and B3 comprising 20%, 33%, and 20% of the isolates, respectively (Fig. 1). For pbp1a, the largest group (profile A1) comprised 36% of the isolates. Majority of profile A contained isolates with serotypes 14 (36%) and 19F (30%). Out of 32 PRSP isolates, 63% showed A1 pattern. The pbp2x showed the most diverse profiles with the largest group, profile X1, covering 68% of the strains. Serotype 14 (25%) was the most dominant serotype in profile X1 (Fig. 1). All isolates with B4X1A1 pattern exhibited penicillin resistance with MIC ≥2.0 μg/mL.

UPGMA dendrogram showing PFGE patterns of clinical and normal flora PNSP isolates. Each pulsotype is shown in relation to the source of isolation and phenotypic and genotypic characteristics. Eye I, eye infection; MIC: minimum inhibitory concentration; NOR, normal flora; PFGE, pulsed-field gel electrophoresis; PNSP, penicillin-nonsusceptible S. pneumoniae; UPGMA, unweighted pair group method with arithmetic averages.
PBP sequence variations
The diversity of amino acids in PBP2b (codons 210–675), PBP2x (codons 280–600), and PBP1a (codons 310–650) was determined for all isolates, and great heterogeneity was observed. The deduced amino acid substitutions at positions in or close to the three conserved motifs were identified (Supplementary Table S1. In total, we observed that 178, 133, and 203 amino acids were changed for PBP2b, PBP2x, and PBP1a, respectively, compared with the R6 control sequence. Based on the changes of PBP2b amino acid sequence, eight different groups (G1–G8) were found among the isolates. The most and least numbers of amino acid differences belonged to G1 (55%) and G8 (2%), respectively. All isolates contained the substitution of adenine for thymine at the amino acid position 445 (Supplementary Table S1), which is next to the second PBP2b conserved motif serine-serine-asparagine (SSN). PBP1a contained seven groups, G1 and G6 having the highest (35%) and the lowest (5%) changes in the amino acid sequence, respectively.
In conserved region of PBP2x (STMK), all groups had amino acid substitution of STMK→SAMK. In LKSG region, the three groups (G1, G2, and G5 with 68%, 6%, and 4% isolates, respectively) showed substitution of LKSG→VKSG. Except for G6, all PBP1a isolates had amino acid substitution in the STMK conserved region. Four groups had substitution of STMK→SAMK, and G4 and G7 showed substitution to SSMK.
Among the PRSP isolates, serotype 14 showed the highest number of amino acid substitutions (up to 38 amino acids) (Table 2). The results showed that the majority of serotype 14 in the G1 group (70%) carried an amino acid substitution in PBP2x region. Similarly, in PBP1a and PBP2b regions, 56% and 26% of the isolates carried amino acid substitutions, respectively (Table 2).
Amino Acid Substitutions in Penicillin-Binding Proteins in Relation to Their Serotypes
Isolates with ≤5% are not shown.
G, restriction fragment length polymorphism patterns for each gene were grouped by visual judgment; N, number of amino acid substitutions; Non, indicates nontypable; PBP, penicillin-binding protein.
PFGE of PNSP isolates
Analysis of 93 isolates displayed 80 PFGE types with 8 common types (CT) constituting 21 (23%) isolates with 95% similarity. The remaining 72 isolates (77%) were single types. Among 61 PISP and 32 PRSP isolates, 55 and 29 different PFGE types were found, respectively. CT3, CT4, CT6, and CT7 were shared by PRSP and PISP isolates (Fig. 1). In contrast, CT1, CT5, and CT8 were found only in PISP isolates. All isolates with CT2 were PRSP. The predominant serotypes in CT1, CT2, CT5, CT6, CT7, and CT8 were serotype 3 (100%), 14 (100%), 14 (100%), 14 (100%), 23F (100%), and 23F (100%), respectively. In contrast, CT3 showed more than one serotype [19A (50%), 19F (50%)] and CT4 [14 (50%), 19F (25%), 19A (50%)].
MLST of PNSP isolates
A total of 90 isolates were analyzed by MLST. The three predominant sequence types (ST3130, ST81, and ST557) accounted for 38% of the PNSP isolates. Our biggest complex was associated with clone complex 63 comprising four STs (63, 3130, 2678, and 5004) with 29 isolates. The eBURST analysis revealed 36 different STs, including 25 singletons and 4 groups (11 different STs with 47 isolates), based on similarities in 6 out of 7 loci (Fig. 2).
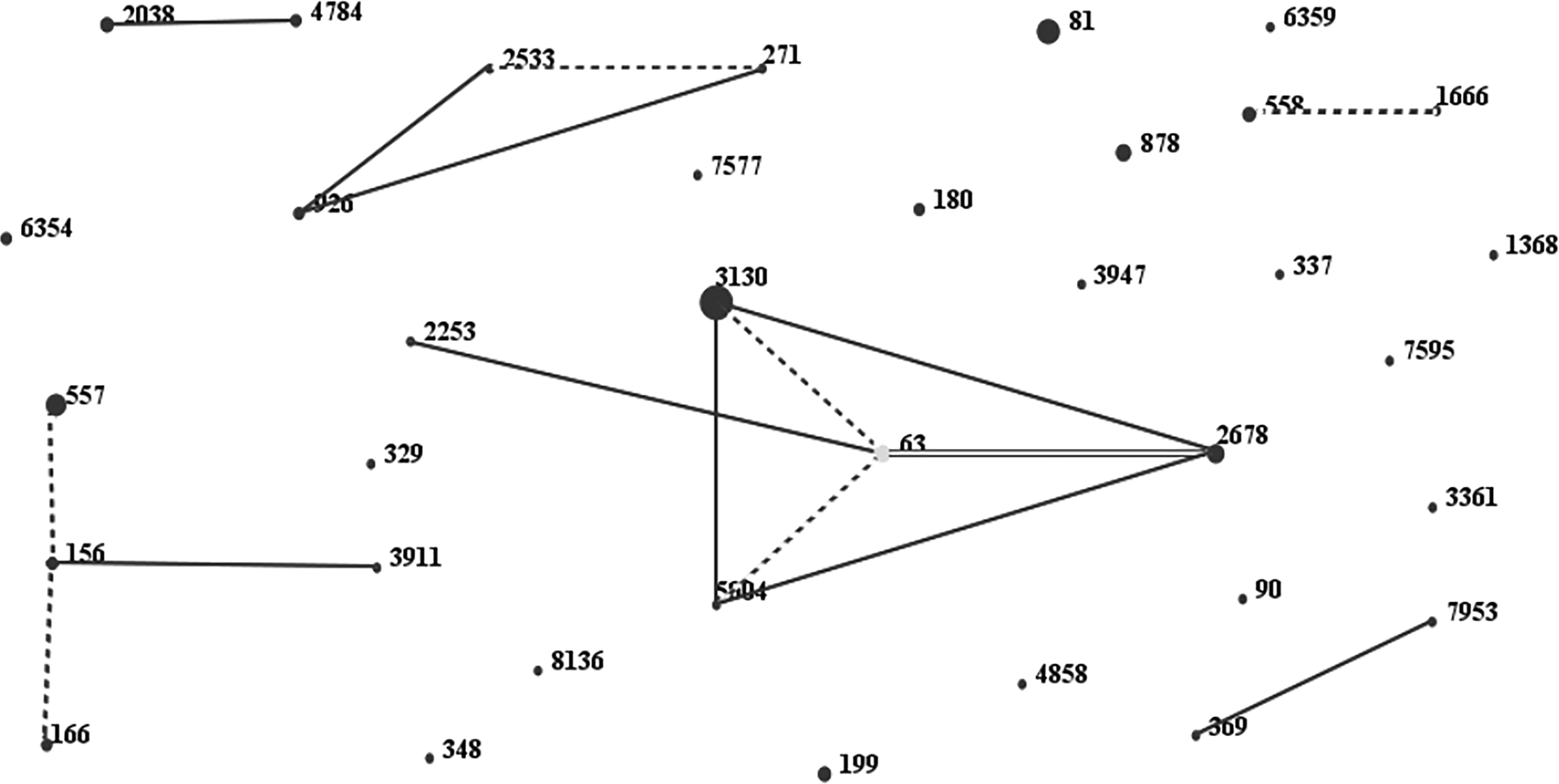
PNSP isolates as shown by eBURST analysis. One spot indicates one ST. The size of one spot relates to the number of pneumococcal isolates with the same ST. The lines designate the presence of SLV (dotted line) and DLV (solid line) links among particular ST. DLV, double-locus variant; SLV, single locus variant; ST, sequence type.
Group 1 consisted of 29 isolates with 4 STs (63, 3130, 2678, 5004) and ST63 as sequence founder, group 2 comprised 12 isolates with 3 STs (557, 166, 156), group 3 comprised 2 isolates with 2 STs (2533, 271), and group 4 comprised 4 isolates with 2 STs (558, 1666). The three predominant STs, ST3130 (28%), ST81 (17%), ST557 (12%), accounted for 51% of the isolates. Distribution of MLST was statistically significant in some serotypes. ST63 showed correlation with serotypes 14 (100%) and ST81 with 23F (90%) (Table 3). No new undefined ST was found among our isolates. Out of 29 isolates in the group 1, the predominant serotypes were 14 (n = 15, 52%) and 19F (n = 11, 38%). In group 2, 3, and 4 the predominant serotypes were 14, 19A, and 18C with 5 (42%), 2 (100%), 2 (50%) isolates, respectively. Moreover, the predominant category of the MLST was singleton with 43 isolates (48%) with serotype 23F as the major serotype (Table 4).
Distribution of Multilocus Sequence Typing in Relation to Serotypes
MLST sequence types ≤2 isolates are not shown.
MLST, multilocus sequence typing.
Correlation Between Multilocus Sequence Typing Groups and Serotypes
Among the clinical isolates, ST63 and ST81 were observed only in the invasive and noninvasive isolates, respectively. Serotype switching was observed in isolates with STs 3130, 81, 557, 2678, 878, 156, 166, 2038, 4784, and 6354. ST3130 was associated with serotypes 19F, 14, and 19A, while ST557 was associated with serotypes 14, 19F, 9V, 11A, and 23F.
Discussion
S. pneumoniae phenotypic and genetic characteristics are affected by geographical distributions and in clinical cases by anatomical sites of isolation and age. It is, therefore, of global and national interest to examine the features of S. pneumoniae and its burden on public health in each country. The outcome of such studies, even though it might look local, could benefit the global health status by implementing pneumococcal vaccine immunization program.
The high rate of penicillin resistance in our isolates was evident. The overall rates of PISP and PRSP in our isolates were 38% and 20%. Similar rate of resistance has been reported in United States and China (around 56%).15,16 In the present study, an increased MIC for many serotypes (14, 18C, 19A, 19F, 23F, 6A, 6B, 9V) was observed. Among PNSP, 44%, 39%, 29%, and 25% of the isolates showed MIC ≥2.0 for 19F, 14, 23F, and 19A serotypes, respectively. Dissemination of serotype 19A isolates has been increased, and they have become resistant to penicillin in many parts of the world. This has been attributed to the selection pressure after widespread use of PCV7.17–19 Serotype 19A isolates, in this study, were mostly penicillin susceptible indicating that the isolates were affected by the local environmental milieu. Only 25% of the 19A serotypes were resistant in this study, suggesting steady increase before implementation of national PCV program in Iran where no national PCV is carried out at the present time.
Tai 19 has conducted serotype analysis in seven Asian countries and has found that four (6B, 14, 19F, and 23F) out of seven serotypes present in PCV7 were isolates which predominantly are collected from clinical settings. In this study, we found that the three most prevalent serotypes (14, 23F, 19F) accounted for 60% of PNSP isolates. The results showed that the overall serotype distribution in Iran was resembled to the countries before the introduction of PCV,15,19,20 suggesting that the use of PCV7 and PCV13 could potentially cover 62% and 93% of the infected individuals, respectively. Furthermore, our results indicated that PCV23 may have equal potential coverage as PCV13 for the PNSP isolates. Therefore, it can be suggested that because of the cost of implementing pneumococcal national vaccination program, PCV13 is the preferred vaccination for the Iranian population and, perhaps, in developing countries.
MLST analysis indicated that all isolates studied here possessed STs which have been reported in the PMEN (http://web1.sph.emory.edu/PMEN/pmen_table 1.html). Our predominant clones, including [Spain23F-1 (ST81), Spain6B-2 (ST9-), Spain9V-3 (ST156), Sweden15A-25 (ST63), Netherlands 3 -31 (ST18-), Netherlands15B-37 (ST199)], are considered as MDR international clones. Out of these 6 international MLST clones, three clones [Spain 23F-1 (ST81), Spain9V-3 (ST156), Sweden15A-25 (ST63)] were more significantly present than the other clones. On the contrary ST320 [a double-locus variant of Taiwan19F-14 (ST236)], which has been reported in many countries as problematic strain and commonly found in Europe, North America, China, and in at least 10 other Asian countries,20,21 was not observed in the present study. In addition, we noted the presence of ST878 isolates in our samples. This strain has been reported by the Spanish Pneumococcal Reference Laboratory (SPRL) to be on the rise at the global scale. 22
S. pneumoniae ST81 was one of the first identified pandemic penicillin-resistant clone. 23 It has been reported that ST81 has undergone capsule switching from 23F to 14, 19A, and 19F. 24 With exception of one isolate belonging to serotype 4, we did not observe any significant class switching for ST81. In contrast, ST557 (single-locus variant of ST156Spain9V-3 clone) was present in the isolates with different serotypes, and an eBURST analysis showed ST557 as the predicted founder of a large clonal complex. We found high serotype switching in ST557 isolates (five different serotypes out of seven isolates). Such a high capsular switching has been reported in different countries in postvaccination era, 25 which is contrary to the present situation in Iran where no public vaccination is available.
PBP alterations have been extensively described as the major mechanism leading to penicillin resistance in S. pneumoniae. It has been indicated that the changes within PBP2b and PBP2x are essential for lower level of β-lactam resistance, and changes within PBP1a are crucial for high-level resistance to penicillin.26–28 In all of our PNSP, we identified mutations in at least one of the genes for PBP2b and PBP2x. Our results showed that up to 10% of the amino acids could be substituted in PBPs, which are in accordance with the report by other investigators. 29 Analysis of PBP2b has revealed the lack of substitution in or close to the active site of serine of the conserved SVVK motif in our pneumococcal isolates. These findings are in agreement with results of other studies, suggesting that the SVVK motif might not be involved in penicillin resistance.30,31
Our analysis of PBP2x and PBP2b motifs showed that the most predominant mutations in PNSP isolates were Thr338→Ala and Thr445→Ala, respectively. It has been reported that Thr338 substitution is mostly found in the clinical isolates with β-lactam resistance.32,33 In this study, we found such substitution in the clinical, as well as the normal flora, isolates. Moreover, our data showed that Thr445→Ala substitution adjacent to the conserved SSN motif of PBP2b was present in all PNSP strains. All PNSP isolates with characteristics of intermediate or high level of resistance to penicillin contained mutations at T338 and T445 positions, respectively, suggesting that these mutations may, in part, dictate the level of penicillin resistance. Similar to the reports from other countries,11,34 our PNSP isolates showed that the amino acid substitution of Ala618→Gly in PBP2b gene, adjacent to the third conserved (KTG) motif, was found in highly penicillin resistant isolates (three out of six isolates). The amino acid substitution at Leu546→Val preceding the third conserved motif (KSG) of PBP2x, which is associated with high-level β-lactam resistance,33,35 was observed in 3 groups (G1, G2, and G5) with 73 isolates. The MICs for these isolates were ≥1 μg/mL. In addition, in accordance with the report by other investigators, 32 we found PBP1a with T371→A or S substitution. This type of substitution has been associated with high level penicillin resistance as well.
In conclusion, the results indicated that the mutational motifs were responsible, to different extent, for penicillin resistance in our isolates. In addition, the extensive sequence variations in the transpeptidase region of PBP2x, PBP2b, and PBP1a were observed in this study covering both intermediate and resistant strains of clinical, as well as commensal, S. pneumoniae, suggesting widespread homologous recombination within the pneumococcal species.
Conclusions
The present study explores the genetic and phenotypic variations among Streptococcus pneumoniae isolates in Iran. There are only a few studies, without much of the details, about this subject matter from the Middle East. This in-depth analysis shows the need for use of conjugated pneumococcal vaccine 13 which is not in use in Iran at the present time. Our results support the notion that the identification and continuous monitoring of the S. pneumoniae clones are needed globally.
Footnotes
Acknowledgments
This work was supported by a grant from the Deputy of Research, Iran National Science Foundation. The funders had no role in study design, data collection and explanation, or the decision to submit the work for publication.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.