
Abstract
Cholera, caused by the Gram-negative bacterium Vibrio cholerae, remains a major problem in developing countries. Although the disease can be managed by oral rehydration therapy, antibiotics are widely used nowadays to treat the disease. However, chemoprophylaxis has been proven to have no effect on the spread of the disease, but acts as a major driver for antimicrobial resistance (AMR). V. cholerae has evolved different ways to combat antibiotics used against them. This review comprehends the different molecular mechanisms of antibiotic resistance in V. cholerae, gaps in the development of new antibiotics, and the alternative strategies that can be used to treat the disease. The review advocates the use of antivirulence compounds rather than antibacterial compounds as a strategy to limit the increasing AMR. Also, the review expounds the role of community in preventing the diseases and tackling the global burden of AMR.
Introduction
Antibiotics have been regarded as the solution to cure bacterial infections for more than six decades. However, microorganisms have evolved different ways to combat new drugs used against them. The threat of infections caused by microbes that have developed resistance has increased greatly in the recent years, with more than 50,000 deaths each year across Europe and the United States alone. The death toll due to such infections in developing and underdeveloped countries is much higher. 1 There is significant difference globally in the patterns of antimicrobial resistance (AMR). Specifically, in Europe, the majority of the bloodstream-associated infections are caused by Staphylococcus aureus. In developing and underdeveloped nations, high emergence of resistance to tuberculosis (TB), malaria, and HIV has been documented. The variation in the AMR patterns in individual countries is linked to how heavily antimicrobial drugs are used. The consumption of antibiotics has increased globally by 36% from 2000 to 2015 with marked variations across regions. 2 The most prevalent AMR Gram-negative pathogens include Pseudomonas aeruginosa, Salmonella enterica, Vibrio cholerae, Klebsiella pneumonia, and Escherichia coli with aminopencillins being the most common treatment modality. As of 2010, India ranked first in the consumption of antibiotics globally at an average of 12.9 × 109 units (10.7 units/person) followed by China at 10.0 × 109 units (7.5 units/person) and the United States at 6.8 × 109 units (22.0 units/person).2,3
Cholera, caused by Gram-negative V. cholerae, is endemic to India. The acute diarrheal disease is managed with rapid and appropriate oral rehydration for moderate cases or intravenously in case of severe dehydration. Although antibiotics are not required to resolve the disease, appropriate antibiotics can decrease the stool volume, severity of infection, and excretion of the pathogen. This will eventually reduce the volume of rehydration fluids required and spread of infection. 4 However, there are insufficient data on the effect of chemoprophylaxis on the secondary transmission of the disease, and it is assumed to act as a major driver for AMR. 5
Antibiotics such as erythromycin, tetracycline, and quinolones were initially used to treat cholera.4,6 Of late, increasing frequency of resistant V. cholerae strains have been noticed in all cholera-endemic countries, including India.7–14 An emerging trend of fluoroquinolone resistance has also been noted in patients hospitalized with acute diarrhea in Kolkata, India. 15 Emergence of V. cholerae strains resistant to these antibiotics has forced clinicians to replace these antibiotics with doxycycline, a broad-spectrum semisynthetic tetracycline. The pattern of introducing successive new generation antibiotics in clinics/hospitals strongly demonstrates the rise of resistant bacteria in general.
The mechanism of antibiotic resistance in bacteria is well studied. Bacterial gene inheritance, cell physiology, and population dynamics are some of the common phenomena leading to single gene mutations that lead to a resistant phenotype bacteria. 16 Bacterial resistance can occur by the acquisition of mobile genetic elements (plasmids, transposons, and integrons) or by mutation in single or multiple genes.17–20 V. cholerae can readily acquire genetic elements from other bacterial species. Overuse of trimethoprim-sulfamethoxazole and tetracycline exerts a selective pressure on the normal gut flora to maintain respective resistance genes, and turns out to be an important source of resistance genes, which can be transferred to enteric pathogens, including V. cholerae. 21
General Mechanisms of Gene Transfer Conferring Antibiotic Resistance in Bacteria
A significant amount of information regarding the different mechanisms of the dissemination of antibiotic resistance has become clear in the past 20 years. 22 The three main mechanisms by which genes get transferred among bacteria are as follows (Fig. 1a–c):
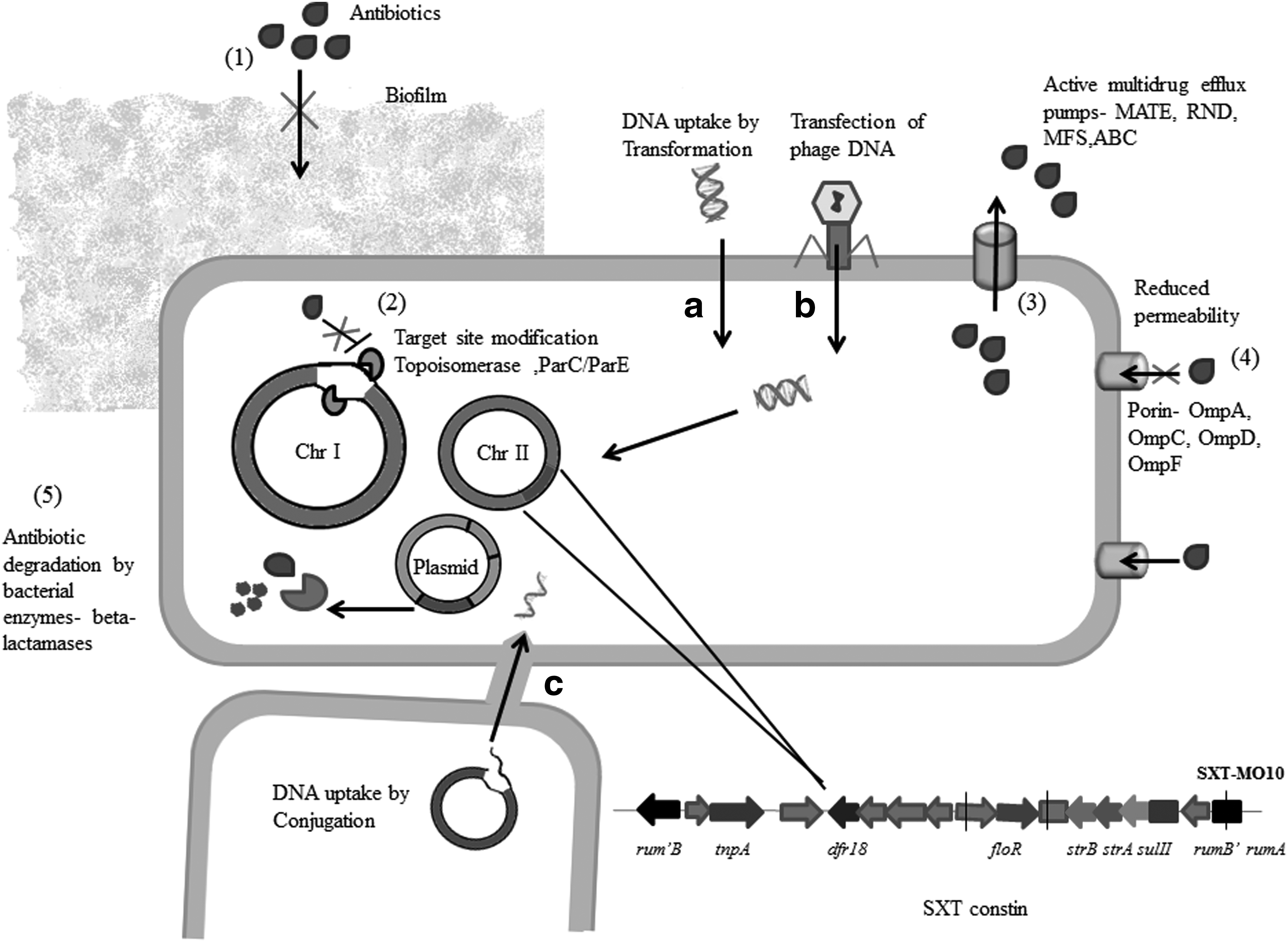
Methods of resistance gene acquisition by Vibrio cholerae (a–c) and its major mechanisms of antibiotic resistance (1–5).
a. Direct uptake of DNA (Transformation).
b. Transfer of genes harbored by bacteriophages (Transduction).
c. Process of mating (Conjugation).
Transformation occurs more predominantly in bacteria of the same species as they share genome homology. In vibrios, cross-species, cross-genera, and even cross-order transfers have been reported. 23
Transduction is phage mediated and is mostly host specific. Bacteriophages integrate its DNA into the bacterium through specific receptor sites and either persist as lysogenic phage or lytic phage. Phages are identified to be transferring entire blocks of virulence genes into the bacterial chromosome, changing a previously avirulent strain into a pathogen in a single step. Infection of V. cholerae with CTX phage is an example for transduction-mediated gene transfer of cholera toxin (ctx) gene cassette to the bacterial genome. 24 Also, filamentous phages like the episomally replicating VFJΦ was identified to transfer ampicillin and kanamycin resistance in V. cholerae. 25 Also, environmentally isolated phages transfer antibiotic resistance to pathogenic bacteria in laboratory conditions. 26 For instance, four phages isolated from effluent water sample were observed to horizontally transfer kanamycin resistance gene to uropathogenic E. coli, and phage 933W from E. coli 0157:H7 resulted in efficient transfer of tetracycline resistance genes to the E. coli laboratory strain, K-12. 27 However, there have not been many reports of phage-mediated antibiotic resistance gene transfer and stable expression of resistance in V. cholerae.
Conjugation allows mobile genetic elements be transferred from a donor to a recipient bacterial cell, through a contact transmission. Conjugation depends on conjugative plasmids that replicate independently and carry genes required for conjugation. 23 Conjugative plasmids can also integrate themselves into the bacterial chromosome.
Transposons have the ability of excising themselves from one locus and move to another locus, within the same bacteria or another bacteria. 28 Transposons can be transferred through transformation, conjugation and transduction and play a major role in AMR gene transfer and get integrated mainly at chromosomal locations known as integrons. 28
Mechanism of Antibiotic Resistance in V. cholerae
Most antibiotics act on bacteria by affecting any of its cellular function, usually in the synthesis of macromolecules and do not affect the eukaryotic cells. Different antibiotics have different targets such as those which inhibit bacterial cell wall synthesis (penicillin), target DNA, RNA, and protein synthesis (quinolones, aminoglycosides, tetracyclines etc.) or inhibit the synthesis of essential metabolites (sulfonamides and trimethoprim). Resistances to different classes of antibiotics have been identified in V. cholerae. The different mechanisms of antibiotic resistance have been represented in Fig. 1 (1)–(5). Lactamases, the antibiotic-specific enzymes produced by bacteria that can degrade β-lactam antibiotics, are one of the prevalent systems of resistance. Mutation of specific antibiotic targets also increases the frequency of resistance progression. Primary examples of this mode of resistance are mutations at gyrA (Ser83Ile) and parC (Ser85Leu) genes of quinolone resistance-determining regions that are targets of the fluoroquinolone drugs. 29 Several mutations have been described in both the quinolone target proteins that result in reduced binding affinities between the drug and the target. 29 Recent reports have highlighted that top-level quinolone resistance is brought about by a series of consecutive mutations in the target genes, rather than a single mutation. 30 Recently, five novel mutations TCG to AAT (Ser60Asn), TAC to TTT (Tyr65Phe), TCG to ATC (Ser85Ile), GCC to TCC (Ala128Ser), and AAA to CGG (Lys129Arg) were identified in the parC gene of V. cholerae that increased the minimum inhibitory concentration of ciprofloxacin. 31
Other mechanisms include inactivation of antibiotics simply by sequestration, metabolic bypass of a protein blocked by the antibiotic, or overproduction of a protein blocked by the antibiotic, such as sulfonamides rendered ineffective by overproduction of p-aminobenzoic acid. Resistance may be confirmed by reducing the uptake of antibiotics or by pumping them out of the cell by active efflux mechanisms such as multidrug and toxic compound extrusion (MATE), major facilitator superfamily, resistance–nodulation–cell division, and small multidrug resistance (MDR). NorM, another putative efflux pump of V. cholerae, is a member of the MATE family of transporters and inactivation of norM renders V. cholerae hypersensitive toward fluoroquinolones, like norfloxacin and ciprofloxacin. 32
V. cholerae and R plasmids
Although antibiotic resistance can result from the accumulation of chromosomal point mutations, the vast majority of clinically pertinent resistance in V. cholerae is due to interchange of genetic information among bacterial strains through plasmids and transposons.
Most of cryptic or uncharacterized plasmids isolated from V. cholerae encode antibiotic resistance R-factors. The plasmids (110–170 kb) are self-transmissible and usually belong to the incompatibility group C. Generally, R-plasmids are unstable in V. cholerae and are easily removed under drug-free conditions. 33 A similar observation had been made earlier by Kuwahara et al. after in vitro transmission of plasmids from Shigella spp. to V. cholerae. 34 However, stability of R-plasmids has also been reported from clinically isolated V. cholerae. 35 R-plasmids in V. cholerae have been found to carry genes encoding resistance to ampicillin, chloramphenicol, gentamicin, kanamycin, spectinomycin, streptomycin, sulfonamides, tetracycline, and trimethoprim, with up to seven resistance determinants on a single plasmid. 36 Most of these genes were acquired from Enterobacteriaceae family members. A large plasmid conferring resistance to more than six antimicrobials was identified to be transferable from V. cholerae to Aeromonas salmonicida, A. hydrophila, V. parahaemolyticus, V. anguillarum, Shigella spp., Salmonella spp., and E. coli. 37 Reports are also available with varying resistance genes capable of being transferred in vitro from fish pathogens to human pathogens, including V. cholerae and V. parahaemolyticus. 38 A 21 kb plasmid having resistance to cephalothin, which could be transferred to E. coli, was isolated widely from Vibrio strains from shrimp ponds. V. cholerae isolated from an outbreak in Matlab, Bangladesh, carried a multidrug-resistant plasmid transferable through conjugation with other bacteria, including E. coli.39,40 Recent studies have also highlighted the sporadic involvement of conjugative plasmids of the IncA/C group (ACPs) in genome plasticity and MDR acquisition in V. cholerae. 41 ACPs are large plasmids (>110 kb) that transfer efficiently by conjugation and code resistance for penicillins, cephalosporins, and carbapenems, conferred by allelic variants of the blaNDM gene. 42 MGIVchHai6, a novel mobilizable genomic island containing a mercury resistance transposon and an integron conferring resistance to chloramphenicol, trimethoprim, and streptomycin/spectinomycin, was identified from strains circulating in Haiti. 43 Also, Ceccarelli et al. identified two cryptic plasmids in clinical V. cholerae non-O1/non-O139 strains isolated from Haiti. Of this, plasmid pSDH-1 encodes 10 putative open reading frames (ORFs). ORFs 7 and 8 of pSDH-1 was identified to contain MobA/MobC proteins, which are a part of relaxosome complex that initiates the transfer of conjugative and mobilizable elements, while plasmid pSDH-2 encodes 11 putative ORFs. ORF3 of pSDH-2 codes for MipA, a scaffolding protein for murein synthesis, which plays a role in host cell persistence in the environment. orf4 and orf5 encode a putative ParD-ParE type II toxin-antitoxin system. 44 These plasmids were identified to have genomic constituents that could transfer antibiotic resistance when acquired under suitable conditions. Plasmid pMRV150 is an IncA/C family MDR-mediating plasmid identified in V. cholerae serogroup O139 strains isolated in 2004. Another IncA/C family plasmid pVC1447 isolated from V. cholerae encoded important components of the type IV secretion system and conjugative transfer. 41 They were identified similar to the sex pilus assembly protein of P. damselae and the type IV conjugative transfer system protein of S. enterica subsp. enterica. In addition, it also possesses three copies of the traF gene that provides synergistic functions in the transfer of plasmids. Also, pVC1447 was identified to possess two sets of ten antibiotic resistance genes, including tetracycline repressor protein (tetR), tetracycline resistance protein class D (tetD), chloramphenicol acetyltransferase II (catII), streptomycin 3-adenylyltransferase (aadA), dihydropteroate synthase (folP), and extended-spectrum β-lactamase TEM (blaTEM) multidrug efflux protein EmrE (qacEdelta1), macrolide efflux protein (mel), macrolide 2-phosphotransferase (mph2), and dihydropteroate synthase (sulI). 41
V. cholerae integrons
Integrons have a major role in the acquisition and spread of drug resistance genes in V. cholerae.17–20 An integron consists of three elements, the intI gene encoding an integrase, a recombination site, attI, and a promoter gene. Several classes of integrons have been characterized based on the integrase gene. All kinds of integrons (Classes 1, 2, and 3) represent multiresistance and have largely been implicated in the dissemination of resistance genes in V. cholerae. 20 Integrons act as gene apprehending systems that capture gene cassettes, which get integrated at the intI site. These mobile genetic elements code for a wide range of resistant determinants. 18 Multiple gene cassettes can be inserted into the integron gene sequence to confer multidrug-resistant phenotype to the bacteria. 17,28,45 Integrons are also assumed to play a role in the transfer of virulence factors such as the ctx genes (ctxA and ctxB), accessory colonizing factors (ace and zot), and genes for the biogenesis of toxin co-regulated pili. 46 The most common integrons in V. cholerae are Class 1 type characterized by the presence of two conserved segments (CSs), the 5′ and 3′ CS. The 5′-CS contains intI, attI site, and promoter, while the 3′-CS codes for sul1, conferring resistance to sulfonamides, and qacEΔ1, conferring resistance to quaternary ammonium compounds.18,47 In addition, the 3′-CS carries the ORF5 encoding a putative protein. Resistance gene cassettes are integrated between the 5′- and 3′-CS at the receptor attI1 site. More than seven different gene cassettes have been described within a class 1 integron and more than 60 gene cassettes have been identified to date. 48 More than one class 1 integron has been observed within the same bacteria. 49 Class I integrons have been detected in V. cholerae O1 strains isolated from Vietnam, 50 Thailand, 51 Mozambique, Africa, 52 Albania, Italy, 53 and Laos. 54 It was reported that class I integron was widely distributed among O1 strains isolated in India before the initial O139 outbreak in 1992, but was less frequent in post-O139 strains.55,56 It was recently reported that class I integrons containing different antibiotic-resistant gene cassettes, for example, genes encoding resistance to streptomycin, ß-lactams, rifampicin, kanamycin, gentamicin, and trimethoprim, are distributed among clinical strains of O1/O139 and non-O1/non-O139 isolated in India. 57 Class 2 integrons have been reported to play a role in mediating MDR phenotype in V. cholerae non-O1/non-O139 serogroups. 58 However, very little information is available about the distribution and role of Class 3 integrons in multidrug-resistant V. cholerae strains. 59
A fourth type class 4 integron or superintegron was identified in V. cholerae O1 strain 46 and also among other members of the family Vibrionaceae.60,61 This superintegron contains 179 cassettes in a 126 kb region, which corresponds to 3% of the total genome, indicating that it may have a role in genome evolution rather than antibiotic resistance. 46 The dfr6 gene in superintegron was identified in non-O1/non-O139 strain of V. cholerae, making them highly resistant to sulfa drugs. 62
V. cholerae SXT constin
SXT, an integrating conjugative element that encodes resistance to trimethoprim-sulfamethoxazole, streptomycin, and chloramphenicol, 63 was originally reported in V. cholerae O139 isolates. The SXT is a part of integrative conjugative elements (ICE), conjugative transposons family that includes R391, 64 R997, 65 and pMERPH. 66 ICEs contain a common set of genes coding for their conjugative transfer, integration, and excision and regulation.67–69 SXT-related ICEs are not limited to V. cholerae. 70 ICEs are self-transmissible mobile elements that are increasingly recognized as important mediators of horizontal gene transfer among prokaryotes. 71 These mobile elements have both plasmid- and phage-like features.
The SXT genetic element is referred to as SXT “constin,” denoting its conjugative, self-transmissible, and integrating nature. 72 The complete DNA sequence of SXT in V. cholerae O139 clinical strain MO10 (SXT MO10) has been analyzed, 73 which revealed to be 99.5 kb encoding resistance genes to sulfamethoxazole, trimethoprim, chloramphenicol, and streptomycin. SXT encodes tyrosine recombinases (Int) that catalyze its integration into the 5′end of prfC, a gene encoding peptide chain release factor 3. 74 SXT integrates site specifically into the chromosome in an int-dependent, recA-independent manner and its excision also requires int, facilitated by the Xis protein. 69
In the past decade, SXT-related ICEs have widespread in Asian and African V. cholerae. Currently, most, if not all, Asian V. cholerae clinical isolates contain SXT-related ICEs. 75 Besides, in Asia, the nontoxigenic environmental V. cholerae isolates of several serotypes have been reported to contain SXT- related ICEs. 59 The other Vibrio species in Asia have also been reported to harbor SXT-related ICEs. 76 The increasing number of reports of antibiotic resistance-conferring SXT-related ICEs in diverse pathogens presumably reflects the rampant and unregulated use of antibiotics. The difference in antibiotic-resistant genes in SXT-related constins suggests that these genes are not intrinsic features of this family and it appears that they have acquired antibiotic resistance elements as a mode to become transmissible across the bacteria.
There have been recurrent cholera epidemics in Africa. In the most recent systemic review on AMR of V. cholerae from sub-Saharan Africa, cholera reports from 16 of the 47 countries within the sub-Saharan African region covering Angola, Chad, Madagascar, Namibia, Senegal, South Africa, Togo, Cote d'Ivoire, Democratic Republic of the Congo (DRC), Ghana, Guinea Bissau, Mozambique, Tanzania, Zambia, Kenya, and Nigeria were studied from 29 articles. 77 Genotypic studies on the mutations in antibiotic resistance determinants (gyrA, parC, floR, strA, and strB) were detected in nine (31.1%) and ICEVchAng2 and ICEVchInd5 were reported from seven (24.1%) of the studies, while other studies did not perform genotypic analysis. 77 Dalsgaard et al. reported ampicillin, aminoglycosides, cotrimoxazole, and tetracycline resistance in V. cholerae isolated from Guinea Bissau and also demonstrated that resistant isolates possessed a multiresistance transmissible plasmid that encoded trimethoprim (dhfrXII) and aminoglycoside resistance. 78 Atypical El Tor V. cholerae O1 Ogawa was reported from Senegal, which was resistant to streptomycin and cotrimoxazole. 79 Analysis of a panel of V. cholerae clinical isolates from the DRC highlighted a loss of sensitivity to leading antimicrobial drugs, although strains remain susceptible to fluoroquinolones. However, a risk for emergence and spread of fluoroquinolone-resistant strains exists, as has been shown elsewhere in Africa. Because resistance to nalidixic acid is frequently associated with decreased susceptibility to fluoroquinolones, nalidixic acid resistance must be detected to monitor the emergence of highly resistant strains.
V. cholerae biofilm
Apart from being a pathogen with high genetic plasticity, V. cholerae is also a good biofilm former. Biofilms are surface-attached microbial communities encased in extracellular matrix produced by the bacteria. The pathogen forms biofilm in both aquatic and intestinal phases of its lifecycle and also on both abiotic and biotic surfaces. 80 Biofilms offer increased tolerance to antibiotics, to the bacterial cells persisting in them by restricting the diffusion of antibiotics. Also, the horizontal gene transfer occurs at a higher rate in the biofilms. Several mechanisms, including conjugation and transformation occur due to the close proximity of cells within the biofilm and availability of free extracellular DNA in the extracellular matrix (ECM). 81 Various genetic determinants (vpsA to vpsK, vpsL to vpsQ, c-di- GMP, HapR, and lux system) regulate the biofilm formation of V. cholerae. The differential expression of vibrio polysaccharide genes (vpsA to vpsK and vpsL to vpsQ) and matrix genes (rbmA, rbmC, and bap1) is regulated by a complex quorum-sensing machinery of V. cholerae. Another important mechanism by which the bacterial cells within the biofilm become drug resistant/tolerant is by forming a quiescent subpopulation often known as the persistent population. In V. cholerae, the stress response pathway, especially the Save Our Soul (SOS) (RecA), is important for the survival of persistent cells. 82 SOS is a global response to DNA damage which initiates DNA repair. The pathogen is identified to gain resistance to antimicrobials, such as aminoglycosides, chloramphenicol, and tetracycline, by stimulating the SOS response. 83 Thus, the biofilm formation and persister subpopulation development of V. cholerae pose additional challenges in the treatment of the disease.
Gaps in the Research for New Antibiotic Development
The development of new antibiotics has slowed considerably in past decades. Only two classes of new antimicrobials (oxazolidinones and lipopeptides) have been developed over the past 30 years. 80 These antimicrobials are designed to treat infections caused by multidrug-resistant Gram-positive bacteria and are not very efficient against Gram-negative bacteria like vibrios. 81 The other antibiotics that have been developed and reached the market belong to existing classes of antibiotics and are not efficient against most of the microorganisms already resistant to antibiotics in these classes. In 2012, 109 antibiotics were in the pipeline to enter into clinical trials. Among these, 70% reached the early stages of production. In contrast, only 31 potential candidates reached phase II trials and only nine candidates into phase III trials. 81 However, even the development of new antibiotics, which is time and resource intensive, is no promise to the problems associated with antibiotic resistance as microorganisms are constantly evolving resistance mechanisms.
Whole-genome sequencing (WGS) allows both detection and identification of various resistance determinants and their arrangements in bacterial chromosomes. Most bacterial genomes, even from strains susceptible to all antimicrobials, contain resistance genes or their precursors. 82 Adaptive laboratory evolution with simulated antibiotic stress allows us to expose such precursor genes that can confer antibiotic resistance to susceptible strains. Using such in vitro evolution techniques in combination with WGS allows us to understand evolution in “real time.” 83 Extensive studies with modern techniques like WGS and laboratory evolution models are progressing slowly.
Alternatives to Antibiotic Therapy
Considering the enormous rise of antibiotic resistance and evolution of multidrug-resistant vibrios and other bacterium there is an urgent need to find proper strategies to tackle infections. Recent research focuses on the efficacy of application of nonpathogenic bacteria from the natural flora to combat pathogenic bacteria (probiotic therapy). Probiotic therapy is important in decreasing the spread of antibiotic resistance by reducing the use of antibiotics. Bdellovibrio bacteriovorus and Micavibrio aeruginosavorus were examined in cell cultures and animal models against various pathogens, including, V. cholerae. 84 There are various reports of gut microbes that are a part of the normal flora inhibiting the pathogen itself or its toxin activity, thereby attenuating the disease progression. Specifically, the presence of Ruminococcus obeum can obstruct the intestinal colonization by V. cholerae through furanone signal autoinducer-2 production, thereby repressing several colonization factors. 85 The challenge will be to develop a commercially viable probiotic product, specifically for cholera.
Scientists in the Soviet Union had developed phage therapies in the 1920s. Phages have several advantages over antibiotic therapy. Phages specifically target the pathogenic bacteria, leaving the beneficial bacteria unharmed. Although bacterial resistance to phages is a rare event, researchers have ready replacements of substitute therapeutic phages. Peptides with antibacterial activity have been isolated from frogs, alligators, and cobras. Plants and fungi also produce peptides that can abolish the bacteria. The effect of leucrocin, an antibacterial peptide from white blood cell extracts of crocodile, against clinical strains of V. cholerae is well studied. 86 It targeted the cytoplasmic membrane of the pathogen. However, recent reports suggest that V. cholerae have even evolved strategies to neutralize net negative charge of cell surface molecules with amine-containing substitutions and thereby evade the interactions of cationic antimicrobial peptides, with bacterial membranes. Nevertheless, there is a quest for novel natural compounds that have antibacterial property as they are much safer than antibiotics. Another reliable source of natural compounds is the plants. There are many herbal plants with high medicinal value. Historically, traditional therapeutics of plant sources has been used to treat cholera. 87 The active compounds present in these medicinal plants act on V. cholerae with various pharmacological processes. Some compounds show direct antimicrobial activity against the pathogen, and some inhibit the binding of ctx to GM1 receptors. 88 The antibacterial activity of essential oils from spices such as black cardamom, cinnamon, chili, mustard seed, black pepper, ginger, and turmeric has been well documented. 89 Several antivirulent strategies such as targeting V. cholerae adhesion, colonization, toxin production, and quorum sensing have been developed recently by chemical compounds, and natural compounds such as phytochemicals and nanotechnology. Virstatin (4-[N-(1,8-naphthalimide)]-n-butyric acid), a chemical compound was identified to posttranscriptionally block ToxT, which indirectly reduces the toxin production of the pathogen without affecting bacterial viability. 6-Gingerol, a phytochemical, was also reported to have antivirulent activity by competitively binding to cholera toxin (CT) and blocking the toxin binding to GM1 gangliosides. 90 Zinc oxide nanoparticles were also identified to have similar effects as they disrupt the secondary structure of CT and thereby block its interaction with GM1 receptor. 91 Another major antivirulent approach is the antibiofilm strategy. Various compounds of bacteria, plants, and molecules of animal origin have found to have antibiofilm activity. Bacillus isolate 240B1 codes Acyl-Homoserine Lactone (AHL)-inactivating gene aiiA producing lactonase, and thereby reduces quorum sensing, biofilm formation, and virulence of vibrios. Also, it has been established to control bacterial infection in aquaculture and has been proven to be more effective than antibiotics. 92 Many plant extracts and phytochemicals have been recorded to have antibiofilm activity against V. cholerae. 93 Lactoferrin, a multifaceted molecule of the innate immune system found primarily in milk, has demonstrated to possess antiadhesive activity by iron chelation, thereby reducing the biofilm forming ability of many pathogens, including V. cholerae. 94 Inhibition of autoinducer synthesis, degradation of autoinducer, and inhibition of autoinducer-quorum sensing (QS) regulator interaction are various methods by which antibiofilm compounds work. In recent years, advancements in nanotechnology have enabled unparalleled avenues in developing nanotools to prevent and abolish biofilms. Many metal oxides and polymer nanoparticles have been identified to have potent antibiofilm activity. Also, metal nanoparticle-DNA aptamer conjugates have been identified to be effective against vibrio biofilms, and Flash NanoPrecipitation of water-dispersible CAI-1 autoinducer nanocarriers has been identified to inhibit biofilm formation in the pathogen.95,96 Although these compounds have not been translated to treat cholera, their efficacy has been evaluated in vivo, which produced promising results.
Cholera Prevention as a Tool for Slowing Development of Antibiotic Resistance
Cholera prevention can be achieved by good sanitation practices and use of safe drinking water. Cholera outbreaks transpire when sanitation facilities are limited, usually in the wake of natural disasters like earthquakes, hurricanes, and floods. 97 In several investigations, it has been observed that there are cholera outbreaks in the months of September–October where a significant correlation can be observed between the water temperature, rainfall, and phytoplankton and zooplankton blooms. 98 Initial strategies that should be followed to prevent a cholera epidemic include periodic monitoring of water bodies for the presence of V. cholerae that are both culturable and unculturable states. First responses during outbreaks include chlorination of public and private water sources and public awareness programs to promote personal and community hygiene. Water, sanitation, and hygiene (WASH) interventions before and at times of outbreak have been a very useful strategy for prevention and control of cholera. World's leading health organizations such as the WHO and CDC are engaged in global WASH activities that help prevent cholera and thereby cut down the use of antidiarrheal antibiotics. Common WASH interventions include the following: promotion of hand wash and personal hygiene, construction of latrines, and water treatment and safe water supply. 99
Cholera vaccines are also used widely in cholera-endemic regions to prevent the disease. 100 WHO recommends cholera vaccines only in combination with other preventive measures among the high-risk population. Although the protection conferred by cholera vaccines are moderate, herd immunity can multiply the effectiveness of vaccination. 101 Shanchol (Shantha Biotech, India), Dukoral (SBL Vaccines) mORCVAX (Vabiotech, Hanoi, Vietnam), and Euvichol (EuBiologic Co, Ltd., Chuncheon, South Korea) are the marketing names of four WHO prequalified oral cholera vaccines widely used against cholera. A recent prominent cohort study has proven that Shanchol do not have any detrimental effects on pregnant women. 102 In 2016, WHO approved a single-dose live oral cholera vaccine, Vaxchora, in the United States for adults 18–64 years of age traveling to a cholera-endemic area. Use of such preventive measures may help in reducing the antimicrobial use and thereby decrease the chances of developing AMR.
Communal participation to tackle the challenge of AMR is critical element in cholera eradication. Proper awareness regarding hitches of AMR can lead to better community participation. Appropriate awareness of methods to prevent the diseases such as understanding of WASH practices and use of proper latrines are the most important. Under the WASH program, UNICEF aims at providing clean and safe water to people, especially children in resource-poor countries. In many of the underdeveloped and developing countries, there are reports of open defecation, which leads to spread of infections. “Open defecation free” is a phrase first used in community-led total sanitation programs, which is now a thrust area that drives development of the country as well as limits cholera-like diseases. Simple sanitation facilities like the “Peepoo bag,” “a personal, single-use, self-sanitizing, fully biodegradable toilet that prevents feces from contaminating the immediate area as well as the surrounding ecosystem,” have been implemented in resource-poor countries. 103 Apart from safe drinking water and awareness of cultural acceptance for personal toilet bags, basic hygiene like washing hands, proper cooking of foods (especially seafoods), keeping food away from contaminants, and washing fruits and vegetables before cooking can bring down cholera incidence in cholera-endemic countries. Needless use of antibiotics, especially the quinolone and tetracycline, for even suspected cholera or cholera-like diarrheal cases should be strictly regulated.
Conclusion
The evolution of V. cholerae resistance against antibiotics bears testament to the genetic flexibility and adaptability of the bacterial genome under antibiotic stress, making these microbes ultimate survivors. The emergence of new gene cassettes conferring antibiotic resistance against the new classes of antibiotics is a cause for concern and the integration of virulence factors and resistant determinants on the same gene cassettes may have even greater implication for public health. Depending on the phenotypic characterization of antibiotic-resistant V. cholerae may result in the underestimation of true genetic potential of bacteria to resist antimicrobial compounds. Culture-independent metagenome and resistome analysis will provide a better picture of the genetic potential of bacteria against new classes of antibiotics. Nucleotide sequence analysis of the AMR gene determinants will help identify whether certain anthropogenic process is selecting the specific genotypes. It is critical to understand underlying mechanisms of the dissemination of AMR among V. cholerae. Using in vitro evolution techniques such as the adaptive laboratory evolution with antibiotic stress, followed by WGS to analyze the resistome profile allows us to understand evolution in “real time.” Schematic representation of the work flow of adaptive laboratory evolution and resistome analysis is shown in Fig. 2. The most promising and significant approach to tackle the increasing antibiotic resistance would be an alternative therapy using natural compounds. Compounds that inhibit or act on the bacterial toxins rather than killing the bacteria as such would be an alternative therapy as such approaches reduce AMR progression. High-throughput in silico approaches to screen novel therapeutics and understand their mode of action on V. cholerae are highly warranted.
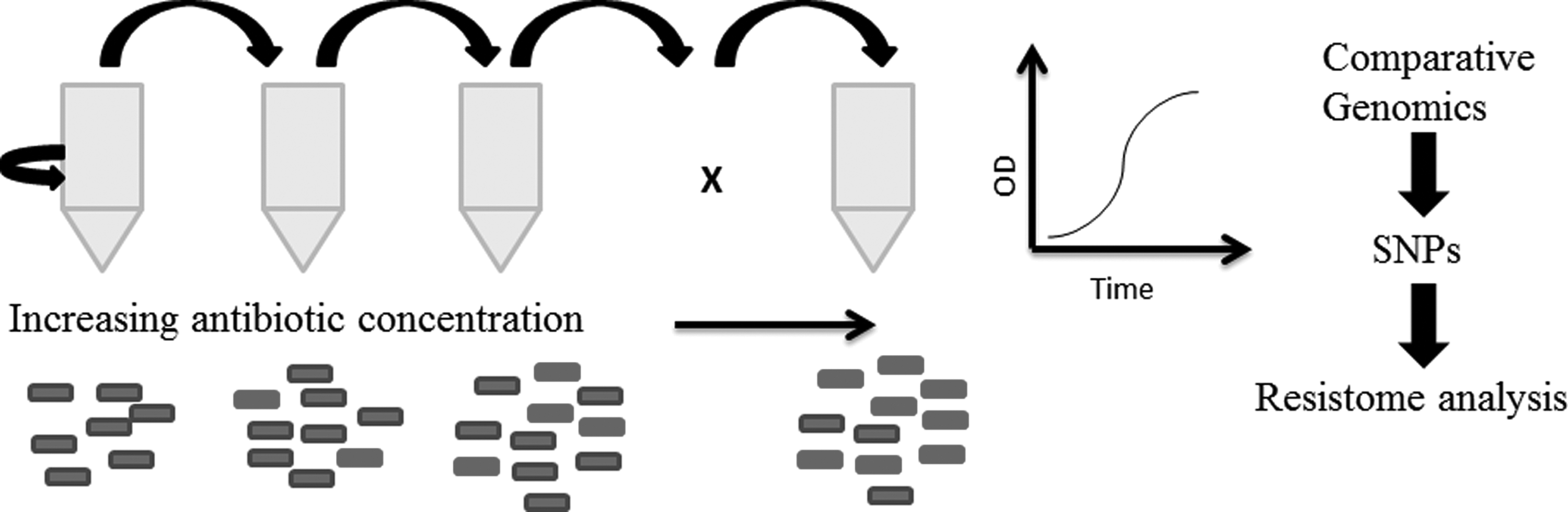
Schematic representation of the work flow of adaptive laboratory evolution and resistome analysis. Bacterial cells grown under increasing concentration of antibiotic. Individual bacterial cells in the population become resistant through random mutations occurring due to the error in DNA repair mechanism. These mutations can be analyzed by whole genome sequencing of the resistant bacterial population and comparing it to that of the sensitive population. Potentially beneficial mutations can be “reverse engineered” into the wild-type sensitive bacteria to empirically validate the cause of resistance.
Footnotes
Acknowledgment
L.N. is thankful to Department of Science & Technology (DST), Govt. of India, for providing the INSPIRE fellowship (Fellow code: IF 140851).
Authors' Contributions
S.T. conceived the idea and L.N. drafted the article. S.S.G. and J.B.J. contributed in the “Cholera prevention” section. R.T. underwrote substantially in the “Antibiotic resistance mechanisms” section. S.T. and R.T. critically reviewed the article. All authors read and approved the article.
Disclosure Statement
No competing financial interests exist.