
Abstract
Objective:
The aim of the study was to decipher the mechanisms and associated genetic determinants responsible for increased carbapenem resistance among Proteus mirabilis clinical isolates.
Methods:
The entire genetic structure surrounding the β-lactam resistance genes was characterized by PCR, gene walking, and DNA sequencing.
Results:
A series of clinical P. mirabilis isolates were consecutively recovered from different patients at the Military hospital of Sofia, Bulgaria. They showed variable levels of resistance to carbapenems. All isolates produced the same carbapenemase VIM-1 that was chromosomally encoded. We showed that increased resistance to carbapenems was related to an increased number of blaVIM-1 gene copies.
Conclusion:
We showed here that increased carbapenem resistance in P. mirabilis may result from increased expression of the blaVIM-1 carbapenemase gene through multiplication of its copy number.
Introduction
Reduced susceptibility to imipenem is intrinsic in Proteus mirabilis, and this feature is mainly related to decreased expression of penicillin-binding protein PBP1a or reduced binding of imipenem to PBP2. However, high-level resistance to carbapenem may be observed in that species due to loss of outer membrane porin. 1 The emergence of multidrug-resistant P. mirabilis isolates producing acquired extended-spectrum β-lactamase (ESBL), AmpC, and carbapenemases has also been described. 2 In P. mirabilis, few isolates producing the carbapenemases KPC-2 (class A) 3 or OXA-23 (class D) 4 have been reported, along with some NDM (class B) producers. 5 However, the most common carbapenemase identified in that species is the metallo-β-lactamase VIM-1.6–8
Interestingly, a previous study showed that variable levels of carbapenem resistance observed among different VIM-1-producing Klebsiella pneumoniae isolates might result from additional porin deficiency, particularly due to loss of OmpK36. 9 In that study, it was shown that some isolates might possess two copies of the blaVIM-1 gene, giving rise to higher hydrolysis rates of imipenem. 9
We investigated here the genetic bases of the variable carbapenem resistance levels of a series of blaVIM-1-positive P. mirabilis isolates.
Materials and Methods
Bacterial strains, antibiotic susceptibility testing, plasmid extraction, and conjugation assays
Proteus mirabilis isolates were identified at the species level using the API 20E system (bioMérieux, La-Balme-les-Grottes, France). Escherichia coli TOP10 (Life Technologies, Cergy-Pontoise, France) and E. coli J53 reference strain were used in cloning and conjugation experiments, respectively. 10 Minimal inhibitory concentrations (MICs) of carbapenems were determined by Etest (AB bioMérieux, Solna, Sweden), and were interpreted according to Clinical and Laboratory Standards Institute breakpoints. 11 Mating-out assays were attempted using the VIM-1-producing P. mirabilis isolate PM5 as donor (representing the clonal strain) and E. coli J53 (azide resistant) as recipient. Selection was made using trypticase-soy plates supplemented with amoxillin 100 μg/mL and sodium azide (100 μg/mL). In addition, extraction of natural plasmids was attempted by using the Kieser method. 12
PCR, cloning, and sequencing
PCR approach was used to detect carbapenemase encoding genes, as previously described. 10 Cloning experiments were performed using plasmid pTOPO as vector, and expression in E. coli TOP10, and selection on trypticase-soy agar supplemented with ticarcillin (50 mg/L) and kanamycin (30 mg/L). Plasmid DNAs were extracted by using Qiagen columns (Qiagen, Courtaboeuf, France). Sequencing of the amplicons and recombinant plasmids was performed by the Microsynth company (Balgach, Switzerland). PCR mapping of the blaVIM-1-surrounding sequences was performed with primers localized in the class 1 integron. 9 PCR fragments generated with primers on which the XbaI restriction site was added were cloned in pTOPO-PCR Blunt vector (Invitrogen, Thermo Fisher).
Quantification of carbapenemase activity
The production of carbapenemase was first evaluated by using the Carba NP test, as previously described. 13 In addition, specific activities with imipenem as substrate were measured. In brief, a 10-mL broth culture of each tested isolate was centrifuged and submitted to sonication. Then, the pellets were, respectively, resuspended in 500 μL of 100 mM sodium phosphate buffer (pH 7) and submitted to sonication. Supernatants were then used for kinetic measurements. 14 Hydrolysis was measured by UV spectrophotometry using 100 μM imipenem as substrate and 10 μL of the bacterial crude extract. The protein content was measured using the Bio-Rad DC protein assay.
Quantification of the blaVIM-1 gene by real-time PCR amplification
Real-time experiments were performed at least three times in different PCR runs, using a Rotor-Gene Q (Qiagen, Hilden, Germany) with a KAPA SYBR FAST qPCR Kit (Kapabiosystems, Wilmington, MA). 15 The hcaT gene was chosen as the normalization control. The qPCR assays included primers VIM-1-qF1 (5′-GAG GTC CGA CTT TAC CAG ATT G-3′) and VIM-1-qR1 (5′-ATC ACC ATC ACG GAC AAT GAG-3′) for the blaVIM-1 gene, and included primers hcaT-Pm-qF1 (5′-CAC TGT GGC TTG CCT TAG AT-3′) and hcaT-Pm-qR1 (5′-CTC GCC CTT TAA CCA GAT AGA C-3′) for the hcaT gene.
The quantity of each target in each isolate was expressed as the difference in Ct values between the hcaT and the blaVIM-1 genes. PCR efficiency was ∼90% for both targets. Real-time PCR was performed by an initial denaturation at 95°C for 60 sec, then rounds were repeated at 95°C for 3 sec, 60°C for 20 sec, and 72°C for 10 sec using primers VIM-1-qF1 and VIM-1-rR1 on one hand, and hcaT-Pm-qF1 and hcaT-Pm-qR1 on the other hand. The Real-Time PCR reaction mix was prepared in a volume of 20 μL containing 20 ng of purified genomic DNA, using the KAPA SYBR FAST qPCR Kit Master Mix (2 × ) Universal, and 0.25 μM of each primer. Analysis was performed using the Rotor-Gene Q machine (Qiagen).
Clonal relationship
Genotyping was performed by pulsed-field gel electrophoresis (PFGE), using restriction enzyme NotI, as previously described. 16
Results and Discussion
Susceptibility testing and carbapenemase identification
During the March–June 2013 period, a total of 23 P. mirabilis isolates were recovered at the Military hospital of Sofia from urine of patients hospitalized in three different units, namely two intensive care units and a neurosurgical ward. They exhibited variable susceptibility patterns to carbapenems, with four isolates remaining susceptible, and the other showing resistance at variable levels. While MICs of imipenem ranged from 0.25 to >32 mg/L, those of ertapenem ranged from 0.004 to 4 mg/L, and those of meropenem from 0.047 to >32 mg/L (Table 1). This prompted us to evaluate whether these isolates might be corresponding to different clones. PFGE analysis showed that the four imipenem-susceptible isolates corresponded to a single clone, and the 19 other isolates to a second clone (data not shown). Results of the Rapid Carba NP test showed that all the 19 imipenem-resistant isolates produced a carbapenemase, but no carbapenemase activity was detected in the susceptible isolates. PCR and sequencing identified the carbapenemase gene blaVIM-1 in all imipenem-resistant isolates. Five isolates were retained for further analysis, with the blaVIM-1-negative isolate being PM1, and four blaVIM-1-positive isolates PM2 to PM5 exhibiting distinct resistance levels to carbapenems (Table 1). Those isolates all showed an identical susceptibility pattern to non-β-lactam antibiotics, being susceptible to fluoroquinolones, tetracycline, chloramphenicol, amikacin, tobramycin, gentamicin, and fosfomycin, but showing resistance to kanamycin and rifampicin.
MICs of Carbapenems for Proteus mirabilis PM1 (Carbapenemase Negative), and VIM-1-Producing Proteus mirabilis PM2, PM3, PM4, and PM5, Being Representatives of the Different Carbapenem Resistance Patterns
Respective blaVIM-1 copy numbers are 0, 1, 1, 2, and 4 for PM1, PM2, PM3, PM4, and PM5.
MICs, minimal inhibitory concentrations.
Carbapenemase activities of the P. mirabilis isolates
To precisely determine the capacity of the different isolates to compromise the efficacy of carbapenems upon hydrolysis, β-lactamase activities were measured using imipenem as substrate. While no hydrolysis could be detected with a crude extract of isolate PM1, enzymatic activities of 43 ± 3, 46 ± 5, 82 ± 3, and 196 ± 8 U/mg of protein (1 U of enzyme activity being defined as the activity that hydrolyzed 1 μmol of imipenem per min) were obtained for isolates PM2, PM3, PM4, and PM5, respectively.
Copy number of the blaVIM-1 gene
To assess whether the increased expression of the blaVIM-1 gene could be related to an increased copy number, quantitative PCR was performed. Isolate PM1 used as control gave a negative result, as expected. For isolates PM2 and PM3, a single copy of the blaVIM-1 gene was detected. Two copies were detected in isolate PM4 and finally, four copies were detected in isolate PM5. Interestingly, the blaVIM-1 gene copy number very well correlated with the MIC data (Fig. 1; Table 1).
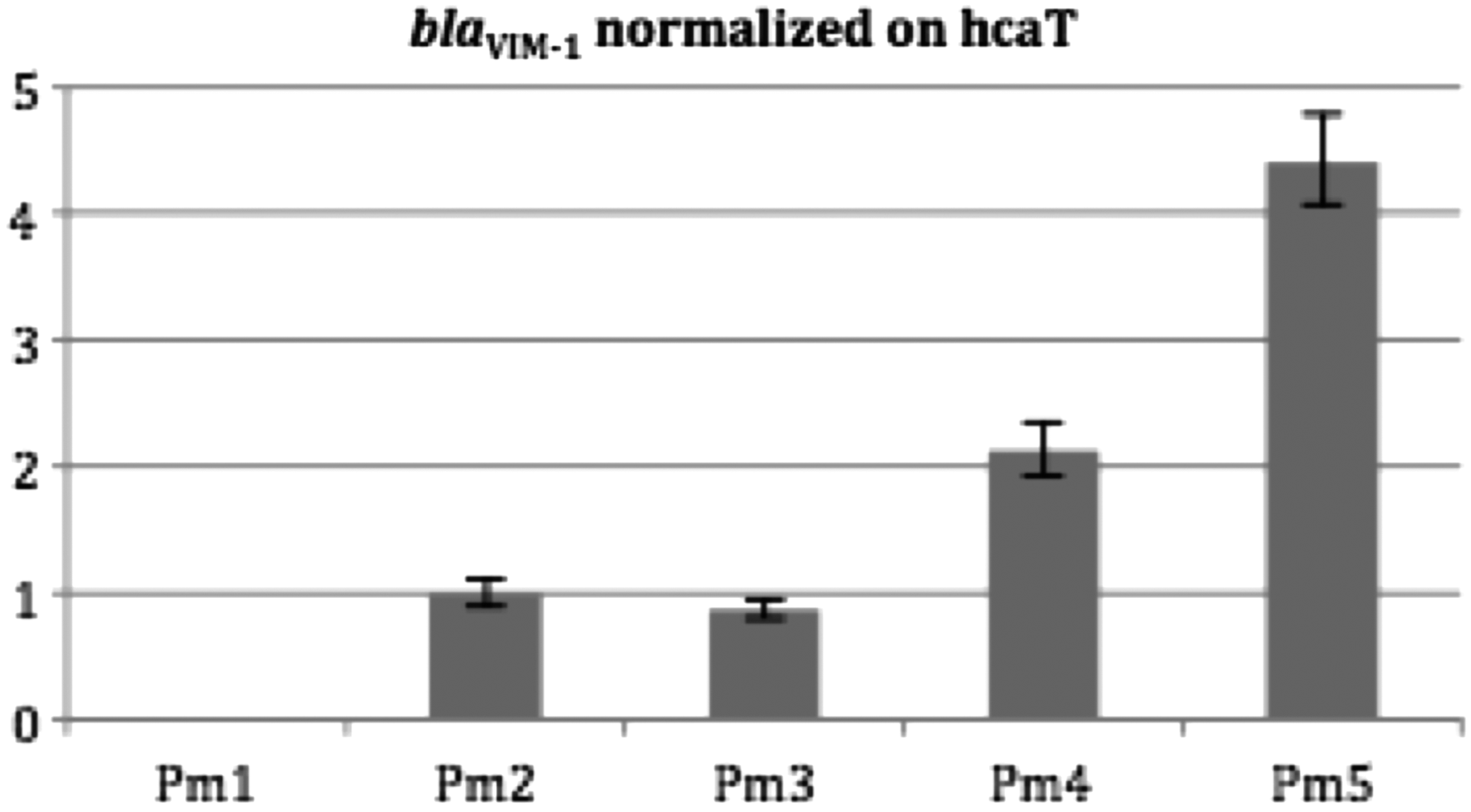
Copy numbers of the blaVIM-1 genes measured by quantitative PCR. Isolate PM1 (blaVIM-1 negative) was used as negative control. Isolates PM2, PM3, PM4, and PM5 (blaVIM-1 positive) were used for testing. Vertical axis; blaVIM-1 copy number normalized on hcaT. Horizontal axis; distribution of the different isolates. Error bars represent the standard deviation between the three experiments.
Genetic structures surrounding the blaVIM-1 gene
Recombinant plasmid pPM2, recovered after XbaI cloning experiments using DNA of P. mirabilis PM2, possessed an ∼8-kb insert and harbored the blaVIM-1 gene into a class 1 integron structure. In fact, the blaVIM-1 gene was the first cassette in the integron structure, followed by aacA7 encoding resistance to aminoglycosides, dfrA1 (resistance to trimethoprim), and aacA1 (resistance to aminoglycosides) (Fig. 2). The 3′-extremity of this class 1 integron was truncated by an IS1 element, and consequently lacked the usual qacEΔ1 and sul1 genes. The whole class 1 integron was bracketed by two copies of insertion sequences IS26, as previously identified in some blaVIM-1-positive strains. 9 Here, a likely IS26-mediated homologous recombination was at the origin of this integron acquisition. In fact, IS26 is very often identified in association with class 1 integron structures. It has been previously shown to be a key element in homologous recombination processes, leading to acquisition of some antibiotic resistance genes, such as blaBES-1. 17 In fact, such integron structure is very similar to the one (named In-e541) reported from a blaVIM-1-positive E. coli isolate recovered in Greece, but differed in its 3′-extremity with the insertion of IS1. 18

Schematic representation of the different blaVIM-1 integron structures identified in isolates PM2 and PM3
By PCR mapping, we showed that isolate PM2 possessed the same unique structure. A second copy of this whole blaVIM-1-positive integron was identified in isolates PM3 and PM4, bracketed by another IS26 element at its 3′-end extremity. Finally, four copies of the same structure were identified in isolate PM5, with IS26 elements always bracketing those different copies (Fig. 2). This structure was likely the result of an IS26-mediated recombination process, leading to multicopies of the blaVIM-1-positive integron.
In all isolates, sequencing showed that the promoter sequences located in the 5′-extremity of the class 1 integrons identified that are known to drive the expression of the right-end located gene cassettes were conserved. Therefore, the overall variability in blaVIM-1 expression level could not be attributed to heterogeneity of the corresponding promoter sequences.
Mating-out assays remained unsuccessful despite repeated attempts, further correlating with a chromosomal location of the blaVIM-1 gene in all positive isolates. In addition, repeated plasmid extractions did not reveal any plasmid band on gel electrophoresis, which is another correlating feature.
Conclusion
This study identified a series of multiresistant P. mirabilis clinical isolates, exhibiting variable susceptibility patterns to carbapenems. Here, we showed that the increased resistance patterns to imipenem observed among those clonally related isolates were explained by an increased blaVIM-1 copy number. Notably, the different blaVIM-1 genes identified were all chromosomally located, thus showing that the process of gene multiplication was not related to a plasmid mobilization. Our data showed that up to four copies of the blaVIM-1 gene could be identified, and that the multiplication process was actually related to transposition-independent IS26-related recombination events. The likely scenario might be that overtime selective pressure by β-lactams (and particularly carbapenems) might have led to the selection of isolates possessing those multiple blaVIM-1 copies. However, we could not establish a direct link with antibiotic usage in that hospital, due to difficulties to retrospectively trace the clonal dissemination within patients.
Tandem amplification of antibiotic resistance genes related to IS26 has been previously reported for the ESBL gene blaSHV-5 in E. coli. 19 In addition, chromosomal amplification of the blaOXA-58 carbapenemase gene has been identified in a P. mirabilis isolate. 20 In the latter case, this genetic event was related to a phage-type recombinase. Nonetheless, the occurrence of blaOXA-58 gene in Proteus spp. (and more generally in Enterobacteriaceae) remains exceptional, while that of the blaVIM-1 is frequently reported. 21 However, the multiplication of the blaOXA-58 carbapenemase gene through IS26-mediated recombination event has been identified in Acinetobacter baumannii, leading to an overtime increased production of OXA-58 and consequently increased MIC values for carbapenems, similarly to what we have observed in this study. 22 Recent studies showed that the involvement of IS26 in the mobilization and dissemination of antibiotic resistance genes occurs more frequently than that previously considered, by reorganizing plasmids by a replicative transposition mechanism. 23
Our study showed that the multiplication of a clinically relevant resistance gene in P. mirabilis can occur and potentially lead to increased resistance. Moreover, the chromosomal location of the duplicated integron probably confers persistence of the resistance phenotype, as opposed to resistance genes located on plasmids.
Footnotes
Acknowledgments
This work was supported by the Swiss National Science Foundation (project FNS-31003A_163432) and by the University of Fribourg.
Disclosure Statement
No competing financial interests exist.