
Abstract
A total of 345 unique (one isolate per patient) clinical carbapenem-resistant Klebsiella pneumoniae isolates were recovered in our hospital over the past 3 years (2016–2018), with 325, 14, and 6 isolates carried blaKPC-2, blaNDM-1, and blaOXA-232, respectively. The six OXA-232-Kp isolates were recovered in neurosurgery intensive care unit in 2018. All OXA-232-Kp belonged to ST15, differed by one or two pulsed-field gel electrophoresis (PFGE) bands, and belonged to the same clone. However, three isolates (RJ18-04-6) displayed elevated carbapenem resistance, with imipenem minimum inhibitory concentration (MIC) of 16–32 μg/mL. In contrast, other three isolates (RJ18-01-3) had imipenem MIC of 0.5–1 μg/mL. S1-PFGE revealed two different plasmid profiles and all strains had at least four plasmids. Strain RJ18-01 and RJ18-06 were selected for whole-genome sequencing, and a ColE2-type 6,141 bp blaOXA-232-carrying plasmid was identified. No blaOXA-232 gene was located on chromosome or other plasmids. Furthermore, no β-lactamase resistance gene other than blaCTX-M-15 was identified. Polymerase chain reaction-based sequencing of blaOXA-232-carrying plasmid was performed, and all OXA-232-Kp shared identical blaOXA-232-carrying plasmid sequences. The expression and copy number of blaOXA-232 in RJ18-04-6 were significantly higher than those of other isolates. The hydrolysis activity of nitrocefin and carbapenems were about six-fold higher in RJ18-04-6. Since only one copy of blaOXA-232 gene was spotted on the plasmid, the elevated carbapenem resistance could be attributed to the increased copy number of blaOXA-232-carrying plasmids. OmpK35 porin deficiency was observed in one isolate with decreased carbapenem susceptibility and two isolates with elevated carbapenem resistance, suggesting that OmpK35 deficiency did not significantly alter carbapenem MICs in OXA-232-Kp.
Introduction
OXA-48-like carbapenem-hydrolyzing class D β-lactamases have been increasingly reported in Enterobacteriaceae, with OXA-48 being the most widespread. 1 OXA-232 is a newly described variant, with five amino acid substitutions differing from OXA-48. 2 The plasmid-borne blaOXA-232 gene has been widely identified from Asia, with related outbreaks being reported in Singapore, 3 Brunei Darussalam, 4 and Korea. 5 Furthermore, clonal disseminations of OXA-232-producing ST15 Klebsiella pneumoniae in adults and neonates have also been reported in East China.6,7 The emergence and wide dissemination of OXA-48-like carbapenemases pose a significant therapeutic challenge to public health.
As one of the OXA-48-like carbapenemases, OXA-232 hydrolyzes penicillins at a high level and carbapenems at a low level, sparing expanded-spectrum cephalosporins.2,8 The catalytic activity of OXA-232 for imipenem was even lower than those of OXA-181 and OXA-48. 8 The majority of OXA-232-producing K. pneumoniae (OXA-232-Kp) isolates only exhibited decreased carbapenem susceptibility, with minimum inhibitory concentration (MIC) of imipenem of 1–2 μg/mL.6,9 However, a considerable number of OXA-232-Kp isolates have been reported with increased carbapenem resistance, with imipenem MIC of >16 μg/mL,7,10 and the underlying mechanisms remain unknown. In this study, we described a clonal dissemination of six OXA-232-producing ST15 K. pneumoniae isolates in adult patients in Shanghai, China, with three of them displaying elevated carbapenem resistance, and the underlying mechanisms have also been investigated.
Materials and Methods
Clinical isolates
A total of 345 unique (one isolate per patient) clinical carbapenem-resistant K. pneumoniae (CRKP) isolates were recovered in our hospital over the past 3 years (2016–2018). All isolates were screened of carbapenemases (blaKPC, blaNDM, blaIMP, blaVIM, and blaOXA-48). 11 Clinical characteristics of the patients with OXA-232-Kp were retrospectively analyzed.
Antimicrobial susceptibility testing
MICs were determined for meropenem, imipenem, and ertapenem using the Clinical and Laboratory Standards Institute (CLSI) reference broth microdilution method. 12 Other antimicrobial agents included piperacillin, ceftazidime, aztreonam, cefepime, piperacillin/tazobactam, ciprofloxacin, amikacin, tigecycline, and polymyxin B. Quality control strains tested concurrently included Escherichia coli ATCC 25922 and ATCC 35218. MICs were interpreted according to CLSI breakpoints, with the exception of tigecycline and polymyxin B where EUCAST breakpoints were applied. 13
Molecular typing
Multilocus sequence typing with seven housekeeping genes (gapA, infB, mdh, pgi, phoE, rpoB, and tonB) was performed for these OXA-232-Kp isolates. 14 Pulsed-field gel electrophoresis (PFGE) analysis was also performed with XbaI-digested DNA (www.cdc.gov/pulsenet/pathogens/pfge.html). Isolates sharing a restriction pattern of less than or equal to three-band differences were considered to be the same clone.
Screening for β-lactamases
Polymerase chain reaction (PCR) detection of β-lactamase genes encoding extended-spectrum β-lactamases (blaCTX-M, blaSHV, and blaTEM) and plasmid-borne AmpC β-lactamase (blaACC, blaCIT, blaDHA, blaEBC, blaFOX, and blaMOX) was performed as previously described.15,16 PCR amplicons were sequenced and compared with sequences available in the GenBank database using basic local alignment search tool (BLAST) searches.
Analysis of outer membrane proteins
The coding sequences of ompK35 and ompK36, with their respective promoter regions, were amplified as previously described. 17 Outer membrane proteins were isolated according to the rapid procedure of Carlone et al. and separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE). 18
Efflux pump inhibitor tests
For OXA-232-Kp, MICs of meropenem and imipenem in combination with PAβN (phenylalanine-arginine β-naphthylamide) (25 μg/mL), 19 an inhibitor of RND pumps of Enterobacteriaceae, were determined. A fourfold decrease in MIC after addition of PAβN was considered significant.
Whole-genome sequencing
Strain RJ18-01 and RJ18-06 were selected for whole-genome sequencing. The genomic DNA was extracted using the QIAamp DNA Mini Kit (QIAGEN, Germany).
Whole-genome sequencing was performed using the long-read PromethION platform (Nanopore, Oxford, United Kingdom) at 200-fold coverage. Reads were assembled using CANU version 1.7. The sequences were polished by Nanopore reads to decrease the errors rates. Sequence similarity was evaluated by BLAST searches. Protein-coding genes were initially identified and annotated using rapid annotation using subsystem technology. Antimicrobial resistance genes were predicted using ResFinder (Center for Genomic Epidemiology).
Plasmid analysis
S1-PFGE was employed to locate the plasmids of OXA-232-Kp. PCR-based sequencing of blaOXA-232-carrying plasmid was performed for all isolates, and the primers used are listed in Supplementary Table S1. Plasmid DNA was extracted from all OXA-232-Kp isolates 20 and was introduced by electroporation into E. coli DH10B. Putative transformants were selected on Luria-Bertani agar containing 50 μg/mL ampicillin. PCR screening for blaOXA-232 was employed for selection of transformants, and antimicrobial susceptibility testing was also performed.
Determination of gene expression
Quantitative real-time PCR was employed to assess the transcriptional expression level of blaOXA-232 in OXA-232-Kp, as well as repA and incA gene for plasmid replication control, with the housekeeping gene, rpoB, as the internal control. The copy number of blaOXA-232 was also measured relative to rpoB, as previously described for blaKPC-2. 21 Standard curves were generated for both the target (blaOXA-232) and the endogenous control (rpoB) using 10-fold dilutions of template DNA at known concentrations.
β-lactamase hydrolysis activity
The hydrolysis activity of β-lactamase was monitored for all OXA-232-Kp isolates at 482 or 298 nm, respectively, using crude supernatants of sonicated bacterial cells at 37°C, with 1 mM nitrocefin or carbapenems as the substrates.22,23 The reaction was performed at 35°C. One unit of enzyme activity was defined as the amount of enzyme that hydrolyzed 1 nmol of substrates per minute.
Results
The emergence of OXA-232-Kp with elevated carbapenem resistance
All 345 CRKP isolates had positive detection of carbapenemases genes, with 325, 14, and 6 isolates carried blaKPC-2, blaNDM-1, and blaOXA-232, respectively. Six OXA-232-Kp isolates were all recovered from sputum in neurosurgery intensive care unit (NICU) from February to April of 2018. The majority of patients with OXA-232-Kp suffered from cerebral hemorrhage, and received carbapenem-based antimicrobial therapy, while one patient died largely due to the serious underlying conditions (Table 1).
Clinical Characteristics of Patients with OXA-232-Kp Infection or Colonization
AMK, amikacin; C, colonization; CIP, ciprofloxacin; I, infection; IMP, imipenem; MEM, meropenem.
All OXA-232-Kp belonged to ST15, differed by one or two PFGE bands, and belonged to the same clone (Fig. 1). However, they displayed varied resistance to carbapenems, with MICs of imipenem of 16–32 μg/mL in three isolates (Table 2), while all OXA-232-Kp remained susceptible to tigecycline and polymyxin B. The first OXA-232-Kp with elevated carbapenem resistance (RJ18-04) occurred only 7 days after the identification of RJ18-01. Two different plasmid profiles were observed through S1-PFGE and all strains had at least four plasmids (Fig. 1). PCR detection of β-lactamase genes indicated that all OXA-232-Kp carried blaCTX-M-15. Interestingly, all of the six corresponding OXA-232 transformants displayed similar resistance profiles to carbapenems, and were susceptible to expanded-spectrum cephalosporins, such as ceftazidime and cefepime (Table 2).

PFGE patterns and outer membrane protein profiles of OXA-232-Kp.
Antimicrobial Susceptibility Testing for OXA-232-Kp and the Corresponding OXA-232 Transformants
TF18-01 to TF18-06 refers to the corresponding OXA-232 transformants of RJ18-01 to RJ18-06.
The numbers in bold indicate that IMP and MEM MICs differed significantly among OXA-232-Kp.
ATM, aztreonam; CAZ, ceftazidime; ETP, ertapenem; FEP, cefepime; MIC, minimum inhibitory concentration; PIP, piperacillin; TGC, tigecycline; TZP, piperacillin/tazobactam.
Increased copy number of blaOXA-232 gene in OXA-232-Kp with elevated carbapenem resistance
The genomes of RJ18-01 and RJ18-06 were 5.7 Mb in size. Strain RJ18-01 harbored a 5,287,250 bp chromosome and 242,614, 145,260, 128,536, and 78,445 bp plasmids, as well as five small plasmids (6–20 kb). In contrast, strain RJ18-06 harbored a 5,277,546 bp chromosome and 175,296, 145,260, 128,536, and 78,445 bp plasmids, as well as four small plasmids (6–23 kb). The genome sequences displayed ∼99% similarity between RJ18-01 and RJ18-06. Two specific segmental insertions, type VI secretion system (5,246–5,264 kb) and tellurite resistance gene cluster (18–34 kb), were detected in RJ18-01 and RJ18-06, respectively (Fig. 2).
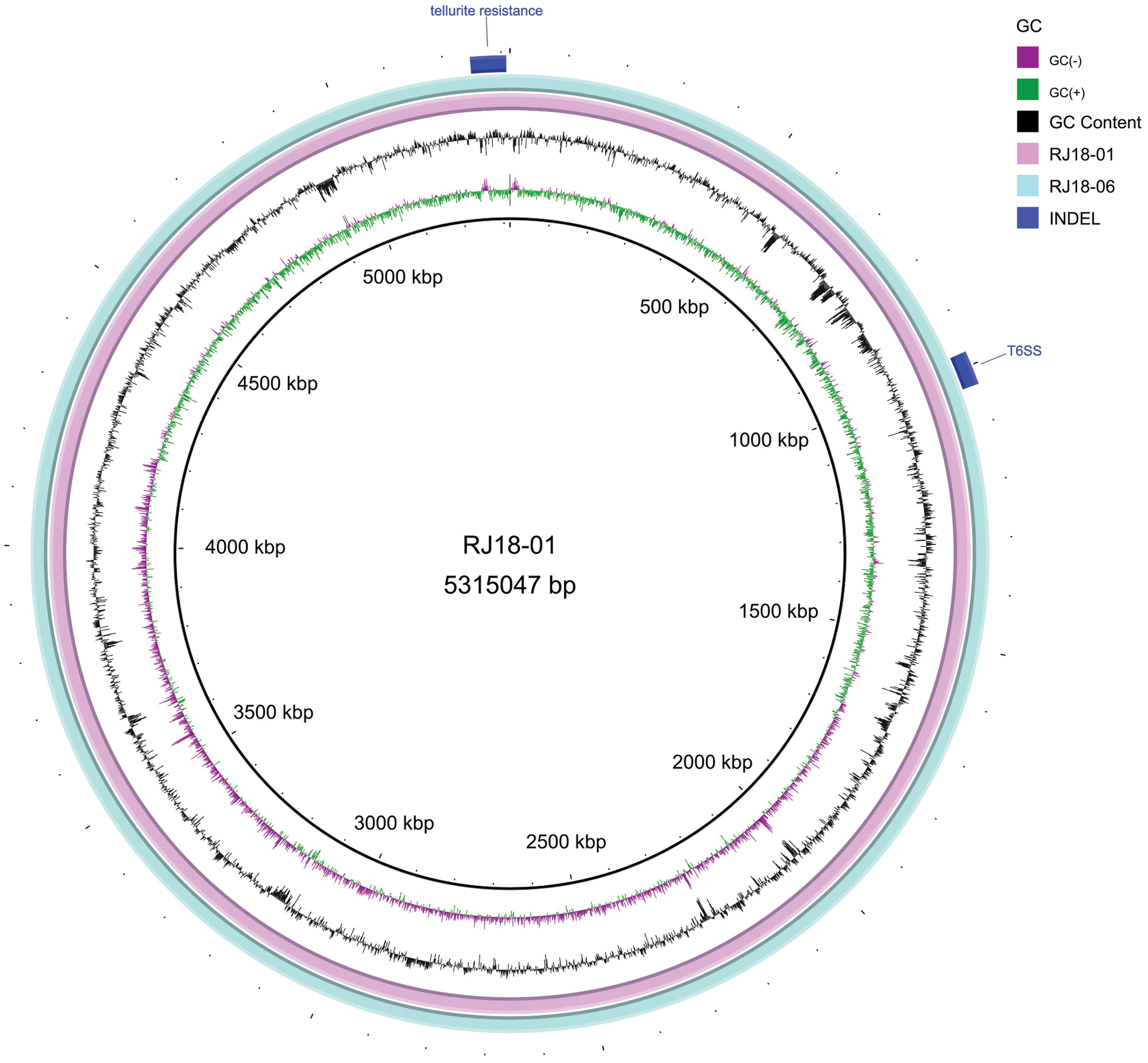
Comparative analysis of the whole-genome sequences of RJ18-01 and RJ18-06. Two specific segmental insertions, type VI secretion system (5,246–5,264 kb) and tellurite resistance gene cluster (18–34 kb), were detected in RJ18-01 and RJ18-06, respectively. Color images are available online.
Whole-genome sequencing identified a ColE2-type blaOXA-232-carrying plasmid in RJ18-01 and RJ18-06, designated pRJ18, with the size of 6,141 bp. No blaOXA-232 gene was identified on chromosome or other plasmids. The blaOXA-232-carrying plasmid contained nine open reading frames, MobC, MobA, MobB, MobD, ΔISEcp1, OXA-232, RepA, ΔEre like, and ΔLysR like. Its sequence was almost identical to the previously reported blaOXA-232-carrying plasmid pOXA-232 (GenBank accession number JX423831), with only two nucleotide substitutions in the noncoding region. Furthermore, no β-lactamase resistance gene other than blaCTX-M-15 was identified in the genomes of RJ18-01 and RJ18-06 with ResFinder.
PCR-based sequencing analysis revealed that all OXA-232-Kp shared identical blaOXA-232-carrying plasmid sequences. The blaOXA-232 gene was located in a truncated Tn2013 transposon in all isolates. The insertion sequence ISEcp1 was almost entirely deleted, with only 207 bp of its right-end extremity being identified upstream of the blaOXA-232 gene (Fig. 3). The relative expression and copy number of blaOXA-232 in the three isolates with elevated carbapenem resistance were significantly higher than those of the other three isolates (Fig. 3). Enzymatic analysis also showed that the hydrolysis activity of nitrocefin and carbapenems was also about sixfold higher in OXA-232-Kp with elevated carbapenem resistance (Fig. 4).

Relative blaOXA-232 expression level, gene copy number, and genetic contexts in OXA-232-Kp.
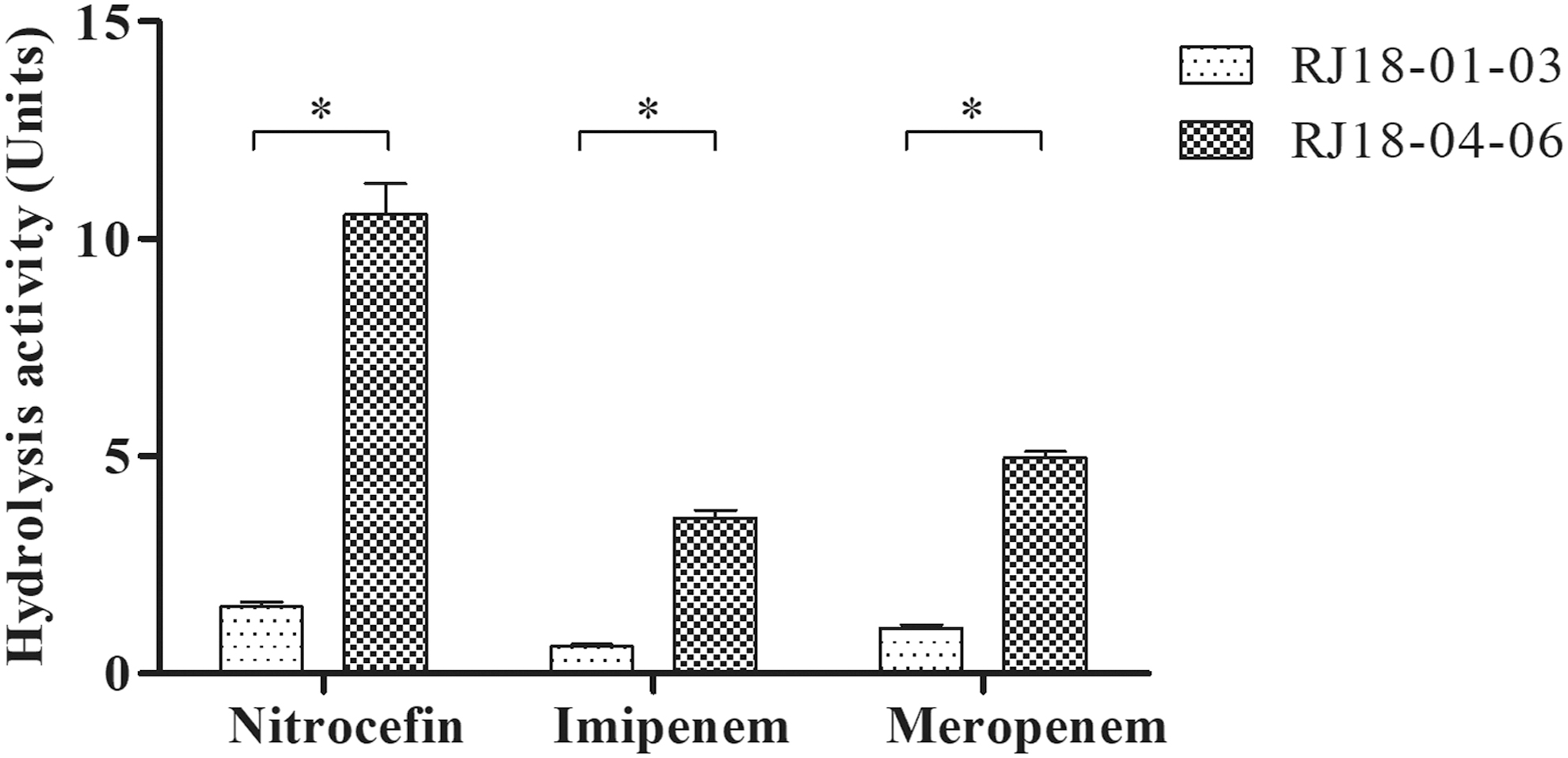
Enzymatic assays in OXA-232-Kp. Nitrocefin, imipenem, and meropenem were employed as the substrates. RJ18-01-03 refers to the three isolates of RJ18-01, RJ18-02, and RJ18-03. RJ18-04-06 refers to the three isolates of RJ18-04, RJ18-05, and RJ18-06. Data were the average of results obtained from three independent experiments and were presented as mean ± standard deviation. *p < 0.05, Student's t-test.
Replication analysis of the blaOXA-232-carrying plasmid
Sequence alignment revealed that replicase-encoding gene repA in this study showed 95% nucleotide sequence identity and 80% coverage to the repA region of the ColE2-type blaGES-5-carrying plasmid pEc1-35771cz (GenBank accession number MF370188) from an Enterobacter cloacae isolate. All OXA-232-Kp shared identical repA, incA, and the flanking genetic regions of replicon. The relative expression of repA and incA in the three isolates with elevated carbapenem resistance was significantly higher than those of the other three isolates (Supplementary Fig. S1). However, when normalized with the gene doses (gene copy number), no significant difference on the gene expression of repA and incA was observed among the six OXA-232-Kp isolates (Supplementary Fig. S1).
OmpK35 deficiency in OXA-232-Kp with elevated carbapenem resistance
The ompK35 sequence of RJ18-01, RJ18-05, and RJ18-06 had an A deletion after base pair 85 when compared to the reference sequence (GenBank accession number KX528042), resulting in an early frameshift and a premature stop codon following the amino acid 62 codon, producing a truncated protein that was nonfunctional, while other three isolates shared identical ompK35 sequence with the reference sequence. In contrast, all OXA-232-Kp isolates shared identical ompK36 sequence. No significant change in promoter region or ribosome binding site of ompK35 and ompK36 was observed.
SDS-PAGE analysis showed that clinical isolates of RJ18-01, RJ18-05, and RJ18-06 lacked OmpK35 porin, but retained normal levels of OmpK36, whereas both OmpK35 and OmpK36 outer membrane protein bands could be observed in the other three isolates (Fig. 1).
Role of the efflux pump in carbapenem susceptibility
The addition of PAβN did not decrease the MICs of meropenem and imipenem by more than twofold, indicating a lack of PAβN efflux inhibition on carbapenem activity in OXA-232-Kp (Supplementary Table S2).
Discussion
In this study, we reported a clonal dissemination of OXA-232-producing ST15 K. pneumoniae in adult patients in Shanghai, China. Since the nosocomial spread of OXA-232-Kp in ICU has also been reported elsewhere,4,5 the ICU has become a preferred ward for the prevalence of OXA-232-Kp. Although the clonal dissemination of ST15 OXA-232-Kp has previously been reported in neonates in Shanghai, 6 OXA-232-Kp isolates identified in this study may have acquired evolutionary significance for the elevated carbapenem resistance. The emergence and rapid dissemination of OXA-48-like carbapenemase in China pose a significant therapeutic challenge to public health.
Three OXA-232-Kp isolates displayed elevated carbapenem resistance in this study, with MICs of imipenem of 16–32 μg/mL. The relative expression and copy number of blaOXA-232 in these three isolates were significantly higher than those of other isolates. Since no significant changes in the surrounding upstream genetic elements of blaOXA-232 were observed, the high expression of blaOXA-232 might be attributed to an increased copy number of blaOXA-232 gene. As only one copy of blaOXA-232 gene was spotted on the plasmid, the high level of carbapenem resistance might arise from the increased copy number of blaOXA-232-carrying plasmids. Despite the fact that OXA-232 possessed a relatively low carbapenem-hydrolyzing activity, the production of copious amounts of OXA-232 through upregulation of resistant plasmid replication can also result in increased carbapenem resistance. Similar phenomenon have also been observed for blaKPC-2-carrying plasmids in KPC-producing K. pneumoniae, 21 suggesting that the upregulation of plasmids copy number is a common approach for establishing elevated carbapenem resistance in K. pneumoniae.
Replication of the ColE2 plasmid requires a plasmid-encoded initiator protein (RepA). 24 RepA binds to the replication origin (oriV) and synthesizes primer RNA for DNA synthesis. The expression of RepA is negatively regulated by antisense RNA (RNAI) at a post-transcriptional level. 25 RNAI is encoded by the incA gene, located upstream of repA. RNAI is complementary to the 5′ nontranslated region of the RepA mRNA and prevents the RepA mRNA translation (Supplementary Fig. S2). However, all OXA-232-Kp in this study shared identical repA, incA, and the flanking genetic regions of replicon. The relative expression of repA and incA in the three isolates with elevated carbapenem resistance was significantly higher than those of the other three isolates. However, when normalized with the gene doses (gene copy number), no significant difference on the gene expression of repA and incA was observed. In other words, the high expression of both repA and incA genes might be the results of the increased plasmid copy number. Combined with the fact that the corresponding OXA-232 transformants all displayed similar resistance profiles to carbapenems, it could be hypothesized that the chromosome, rather than the plasmid replicon itself, might be involved in the upregulation of plasmid replication in OXA-232-Kp with elevated carbapenem resistance.
This study indicated that OmpK35 porin deficiency did not significantly alter carbapenem MICs in OXA-232-Kp, since OmpK35 deficiency was observed in clinical isolates with decreased carbapenem susceptibility (RJ18-01) and elevated carbapenem resistance (RJ18-05 and RJ18-06). Previous studies also revealed that a single deletion of OmpK35 had no significant effect on MIC shifts of carbapenems in K. pneumoniae. 26 It could be concluded that the production of OXA-232 was the key factor contributing to carbapenem resistance in OXA-232-Kp and OmpK35 deficiency might only serve as a minor cooperative factor.
In conclusion, the elevated carbapenem resistance was attributed to the increased copy number of blaOXA-232-carrying plasmids. The high expression of both repA and incA genes in OXA-232-Kp might be the results, rather than the cause of the increased plasmid copy number. OmpK35 deficiency did not significantly alter carbapenem MICs in OXA-232-Kp.
Accession Numbers
Complete sequences of the chromosome of strain RJ18-01 and RJ18-06, and the blaOXA-232-carrying plasmid pRJ18 have been deposited in GenBank under accession numbers CP043670, CP043669, and MH807441.
Ethical Approval
All procedures performed in this study involving human participants were in accordance with the ethical standards of the Institutional Review Board Ethics Committee of Renji Hospital. For this type of study, formal consent is not required.
Footnotes
Disclaimer
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by National Natural Science Foundation of China (grant nos. 81802065 and 81702062), Shanghai Sailing Program (grant no. 18YF1413300), and a research fund from Renji Hospital for young scholars (grant no. PYIII-17-010).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.