
Abstract
It is becoming increasingly recognized that the environment plays an important role both in the emergence and in dissemination of antibiotic-resistant bacteria (ARB), Mechanisms and factors facilitating this development are, however, not yet well understood. The high detection rate of CTX-M genes in environmental sources provides an opportunity to explore this issue. In this study, 88 CTX-M-producing Escherichia coli were isolated from 30 pig feces samples from 30 pig farms and 201 environmental samples. CTX-M-producing E. coli was detected with the following frequencies in the different types of samples: pig feces, 73%; river water, 64%; river sediment, 52%; wastewater, 31%; drinking water, 23%; outlet sediment, 21%; soil, 17%; and vegetables, 4.4%. Dissemination of CTX-M-producing E. coli to different environmental matrices was evaluated by analyzing the genetic relatedness of isolates from different environmental sources, and putative transmission routes through bird feces, pig feces, drinking water, river sediment, river water, and wastewater were hypothesized. Dissemination through these routes is likely facilitated by anthropogenic activities and environmental factors. Wild birds as potential vectors for dissemination of CTX-M-producing E. coli have the capacity to spread ARB across long distances. Regional dissemination between different environmental matrices of CTX-M-producing E. coli increases the exposure risk of humans and animals in the area.
Introduction
At present, antibiotic-resistant bacteria (ARB) constitute a great threat to human health worldwide. If the situation is left unchecked, it has been estimated that the global costs up until 2050 will be 100 trillion USD and an increase in deaths per year up to a staggering 10 million in 2050. 1 To prevent this development, and tackle the global challenge of antibiotic resistance, a “One Health” perspective, which takes into account the connections between human and animal health and the environment, is required. 2
It is becoming increasingly recognized that the environment functions as a reservoir of ARB and antibiotic resistance genes (ARGs), and plays an important role for the dissemination and evolution of these environmental contaminants. ARGs can be readily transferred to humans through contaminated environmental matrices, which contribute to the dissemination and persistence of ARB in the general population. 3 For example, environmental exposure to antibiotic-resistant Escherichia coli has been associated with fecal carriage in humans, including carriage of pathogenic E. coli, which further indicates the risk to humans posed by dissemination of ARB between different environmental matrices. 4
The dissemination mechanisms and risk factors pertaining to dissemination of ARB and ARGs are not well understood and need to be further elucidated to enable risk assessment and design of suitable interventions. 5 To date, most studies focus on ARB and ARGs associated with clinical settings or animal breeding, whereas the environmental sector is usually neglected. Furthermore, environmental studies on ARB and ARGs are usually limited by focusing on a single type of environmental matrix; studies simultaneously investigating multiple environmental sources are rarely undertaken.6,7 Lastly, the detection rates of ARB and ARGs for resistance to last-resort antibiotics are relatively low, which increases the difficulty of drawing conclusions from the gathered data. For example, one study reported that the detection rate of total carbapenem-resistant gram-negative bacilli from a Swedish river downstream of a wastewater treatment plant was 4.5% and 2.5% in influent sewage water and environmental waters, respectively. 8 Other studies have reported on a prevalence rate of 1.3–3.6% of the plasmid-mediated colistin resistance gene mcr-1 among isolates of E. coli from environmental samples.9,10 Therefore, the mentioned ARB are not suitable as biomarkers for environmental dissemination of ARB and ARGs across different environmental matrices.
Extended-spectrum beta-lactamases (ESBLs) are enzymes that hydrolyze the β-lactam ring of penicillins and third-generation cephalosporins, such as cefotaxime and ceftazidime. 11 Since the 2000s, CTX-M genes have become the predominant ESBL type. 12 Among Enterobacteriaceae, E. coli is most frequently associated with production of ESBLs. 13 E. coli is a commensal micro-organism and opportunistic pathogen of the gastrointestinal tract in humans and many animals, and is particularly suited to disseminate ARGs in the environment through the fecal route. 14 E. coli is also the most prevalently isolated Enterobacteriaceae from environmental sources, and is well adapted for survival outside of animal hosts. 15 CTX-M-producing E. coli is widely disseminated and has been reported among various environmental matrices including river water, well water, municipal wastewater, soil, and vegetables.16–20 Accordingly, CTX-M-producing E. coli may be a suitable indicator for studying the mechanisms of environmental dissemination of antibiotic resistance through fecal pollution. 21 In this study, the prevalence and characteristics of CTX-M-producing E. coli among various environmental sources in a rural area of China were investigated to elucidate the dissemination of ARB and ARGs among different animal and environmental matrices.
Materials and Methods
Sampling
The study site is located in a region of rural Shandong province in eastern China. We chose to conduct the research in a rural area because household pigs live in proximity to humans and many residents work as farmers (i.e., they have extended periods of contact with animals and the environment). Twelve villages that hold a number of available backyard pig farms were included in our study. In each village two to three households with backyard farms (defined as harboring <50 pigs) with at least 1 pig were randomly selected. The pig farms chosen usually did not harbor other animals aside from pigs. Additional information regarding the scale of the pig farming in the sampled villages is presented in Supplementary Table S1. During July and September 2015, 30 pig feces samples and 201 environmental samples were collected (Fig. 1) from 12 villages: villages A (n = 27), B (n = 19), C (n = 15), D (n = 22), E (n = 19), F (n = 17), G (n = 19), H (n = 21), I (n = 19), J (n = 18), K (n = 16), and L (n = 19). The environmental samples collected were of wastewater (n = 36), outlet sediment (n = 14), river water (n = 25), river sediment (n = 25), drinking water (n = 43), soil (n = 23), vegetables (n = 23), bird feces (n = 6), and human waste (n = 6). The number of samples and types of samples are summarized in Table 1. The pig feces samples were collected from 30 pig farms (a single sample per farm) and were collected by pooling fresh pig droppings on the ground. Wastewater directly discharged from the pig farms (i.e., without treatment) were collected by using sterilized bottles. Samples of bird feces were only collected from village A; samples were collected twice daily for 3 days from the ground of a villager's yard, which was frequented by swallows. Human waste samples were collected from public toilets, and since some villages lacked public toilets, only six samples of human waste from six villages were collected. Waste samples of animals and humans were collected using the ESwab collection and transport system (Copan, Brescia, Italy). Water samples (from rivers, streams, and wells) were collected by gathering 1 L of water in sterile glass bottles. The sediment samples were taken by a sediment grabber and the sediment cores were sliced into 5 cm layers. At least 30 grams of river sediment and soil samples from vegetable gardens were collected in sterile plastic bottles. Edible parts of different vegetables were collected in plastic bags, including beans (n = 10), cucumbers (n = 6), peppers (n = 3), celery (n = 2), and leeks (n = 2).
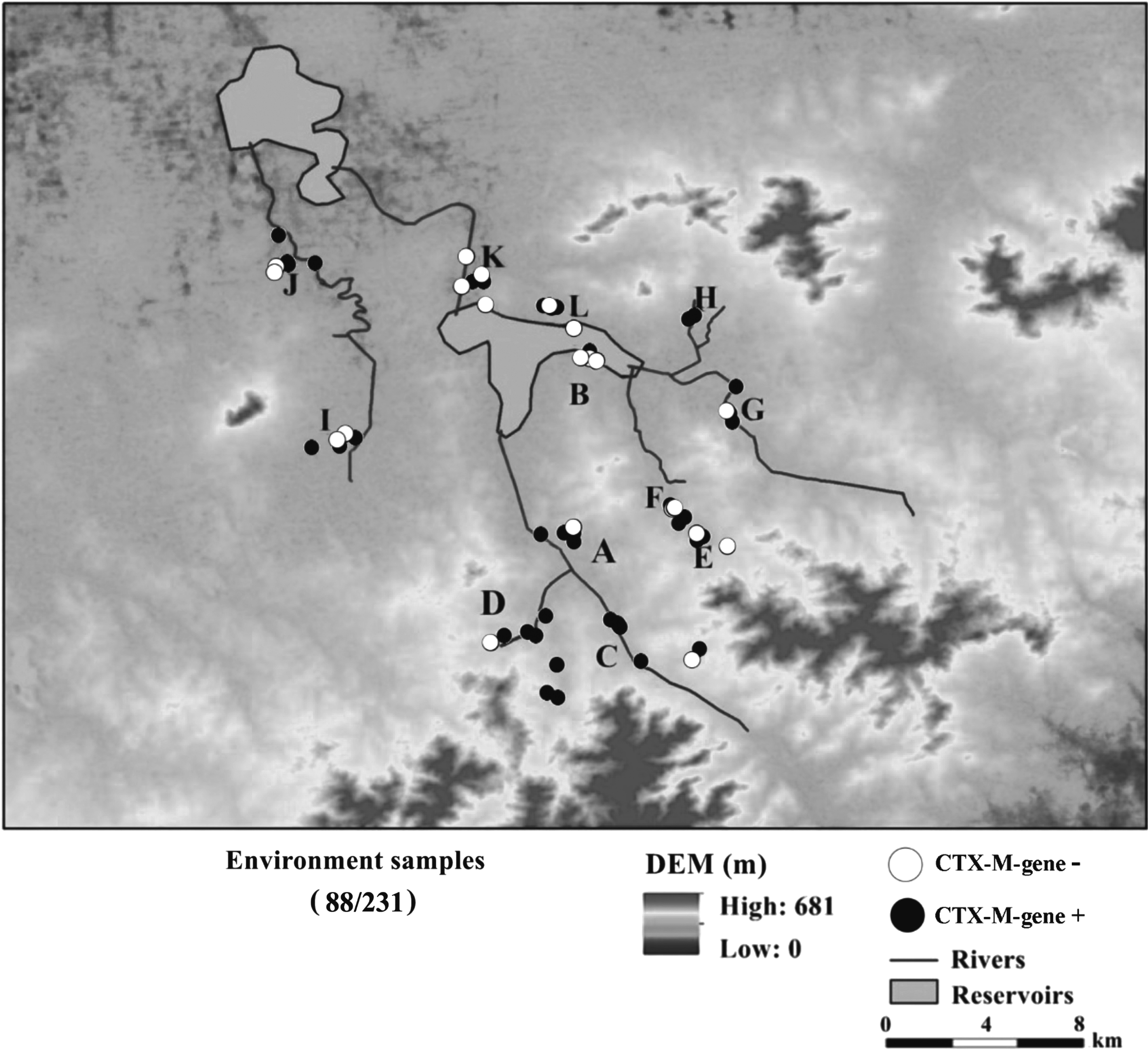
The sampling distribution of CTX-M-producing Escherichia coli among different environmental samples from a rural area in Shandong province, China.
Samples from 10 Different Sources Were Collected from 12 Villages in Shandong Province, China, in 2015
The number of samples positive for CTX-M-producing Escherichia coli are denoted in the table with the total number of samples collected given in parentheses.
All samples were put in cool boxes with ice packs (4–8°C) upon collection, and transported to the local CDC laboratory at the end of the day. Water samples were subjected to filtration through 0.45 μm sterile membrane filters (Millipore, Billerica). After filtration, membranes were transferred into ESwabs and stored at −80°C until cultivation. Sediments (2.0 g) were homogenized in a fivefold volume of sterile brain heart infusion broth (Oxoid, Basingstoke, UK). The vegetable samples were not washed, and the inedible or damaged parts were removed using a sterilized knife. The surface was sampled using the ESwab collection and transport system, and stored until analysis as described hereunder.
Screening of samples for ESBL-producing E. coli
Each sample was inoculated in brain heart infusion broth (Oxoid) and cultured at 37°C overnight. The enriched solutions were plated on ChromID ESBL agar (bioMérieux, Marcy l'Etoile, France) for 18–24 hours at 37°C to isolate potential ESBL-producing isolates. Screening was performed as previously described. 22 Species confirmation of presumptive E. coli isolates was performed by using matrix-assisted laser desorption ionization time-of-flight mass spectrometry. ESBL production was confirmed by using the double disk diffusion method, using cefotaxime, ceftazidime, each alone and in combination with clavulanic acid according to the manufacturer's protocol (Oxoid). E. coli ATCC 25922 was used as control strain.
Genetic characterization of CTX-M genes
DNA was extracted from the ESBL isolates using Gentra Puregene Yeast/Bact. Kit (QIAGEN, Germany), following the manufacturer's instructions. The presence of CTX-M genes was confirmed by using a multiplex PCR as previously described. 23 Four blaCTX-M subgroups (group 1, 2, 8, and 9) were discriminated by multiplex PCR assay as previously described. 24 PCR products were verified by sequencing (BGI Company, Beijing, China).
Antibiotic susceptibility testing
The confirmed CTX-M-producing isolates were cultured on blood agar, incubated overnight at 37°C, and then subjected to antibiotic susceptibility testing using agar dilution with Müller–Hinton agar (Oxoid), according to the Clinical & Laboratory Standards Institute (CLSI, 2016), except for colistin, for which broth microdilution was used. The results were evaluated in accordance with the European Committee on Antimicrobial Susceptibility Testing (EUCAST) breakpoints version 8.1,a except for tetracycline and nitrofurantoin, for which the CLSI breakpoints version 2017 was used, and florfenicol, for which the EUCAST epidemiological cutoff value was used. Multidrug resistance was defined as acquired nonsusceptibility to at least one agent in three or more antimicrobial categories. 25
Molecular typing and determination of putative transmission routes
For multilocus sequence typing of E. coli isolates, internal fragments of the seven housekeeping genes (adk, fumC, gyrB, icd, mdh, purA, and recA) were amplified by PCR from DNA, as previously described, 26 and subsequently sequenced (BGI Company). Sequences were imported into the E. coli Multilocus sequence typing (MLST) database websiteb to determine sequence types (STs). A phylogenetic tree based on the concatenated MLST sequences was constructed by using Molecular Evolutionary Genetics Analysis 5. Annotation of the phylogenetic tree was performed with Interactive Tree of Life.c
Preparation of DNA for pulsed-field gel electrophoresis (PFGE) was performed as previously described.27,28 A dendrogram with the PFGE results was generated using BioNumerics 6.1 (Applied Maths, Sint-Martens-Latem, Belgium). According to the Unweighted Pair group Method using Arithmetic average (UPGMA) cluster analysis, isolates were defined as being of a related clone if dice similarity indices of the PFGE fingerprints were >80%. Putative transmission routes of CTX-M-producing E. coli were determined by identifying related clones isolated from different sources.
Results
A total of 88 CTX-M-producing E. coli were isolated from 231 samples (38%); CTX-M-producing E. coli was detected among all sample types. The detection rate was highest in pig feces (73%), followed by river water (64%), river sediment (52%), human waste (50%), wastewater (31%), drinking water (23%), outlet sediment (21%), soil (17%), and vegetables (4.4%) (Table 1). In addition, five CTX-M-producing E. coli were isolated from three samples of bird feces (two samples were isolated with four different CTX-M-producing E. coli).
Genes of the CTX-M-9 group were most frequently detected (n = 64), and blaCTX-M-14 (n = 35) was the predominant genotype, followed by blaCTX-M-65 (n = 20), blaCTX-M-55 (n = 10), blaCTX-M-15 (n = 9), blaCTX-M-27 (n = 9), and blaCTX-M-3 (n = 5). blaCTX-M-14 was the most common CTX-M gene found in pig feces as well as some of the environmental sources sampled (human waste, bird feces, river water, river sediment, and vegetables), whereas blaCTX-M-65 was the most common found in drinking water, soil, and outlet sediment, and blaCTX-M-55 was the most common CTX-M gene in E. coli from wastewater samples. The distribution of CTX-M genes in different matrices and different villages is shown in Fig. 2. blaCTX-M-14 was found in all isolates except for those from villages F and L. blaCTX-M-65 was predominant in villages E, F, and J, whereas blaCTX-M-27 was the main genotype in villages D and L.

Distribution of CTX-M genes types among CTX-M-producing E. coli from a rural area in Shandong province, China.
All CTX-M-producing E. coli isolates were cefotaxime resistant, and the majority (>80%) of these isolates were also resistant to tetracycline, trimethoprim–sulfamethoxazole, and florfenicol (Table 2). Fifty percent to 75% of the isolates were resistant to fosfomycin, piperacillin–tazobactam, ciprofloxacin, gentamicin, and amoxicillin–clavulanic acid; 43% was resistant to nitrofurantoin, and <25% of the isolates were resistant to colistin, ceftazidime, cefoxitin, and amikacin. Only one isolate was resistant to meropenem, whereas none of the isolates were resistant to tigecycline. All isolates were multidrug resistant.
Antibiotic Resistance Rates of CTX-M-Producing E. coli Isolated from Different Sources in a Rural Area of Shandong Province, China, in 2015
CTX, cefotaxime; TE, tetracycline; SXT, trimethoprim/sulfamethoxazole; FOF, fosfomycin; FFC, florfenicol; TZP, piperacillin/tazobactam; CIP, ciprofloxacin; AMC, amoxicillin/clavulanate; GEN, gentamicin; F, nitrofurantoin; CL, colistin; CAZ, ceftazidime; FOX, cefoxitin; AMK, amikacin; MEM, meropenem; TGC, tigecycline.
A total of 57 STs were detected, including 2 new STs (Supplementary Fig. S1). The CTX-M-9 producers were distributed among 43 ST types (n = 64), whereas the CTX-M-1-producers were distributed among 18 ST types (n = 24). The most prevalent STs were ST10 (n = 10), ST48 (n = 7), ST3489 (n = 4), ST34 (n = 3), and ST1638 (n = 3). ST10 was the most common ST overall, and was found in pig feces, wastewater, drinking water, river water, and river sediment. Five isolates from wild birds belonged to ST216, ST542, ST1011, ST2807, and ST4633. In addition, two isolates (both belonging to ST2614) from pig waste belonged to phylogroup B2, whereas three isolates (belonging to ST38, ST174, and ST2309, respectively) from river sediment, wastewater, and drinking water, respectively, belonged to phylogroup D. The predominant STs were distributed over several villages; ST10 isolates were found in villages D (n = 4), L (n = 3), H (n = 1), I (n = 1), and J (n = 1); ST48 isolates were found in villages C (n = 3), G (n = 2), F (n = 1), and H (n = 1); and ST3489-isolates were distributed in villages B (n = 3) and G (n = 1). Isolates sharing STs were found in villages A, B, C, D, G, J, and L (Supplementary Fig. S2).
The PFGE analysis revealed a high clonal diversity among the isolates, nevertheless, genetically related isolates could be determined based on the PFGE dendrogram (Supplementary Fig. S3). Twenty-three groups (consisting of two or more isolates) of related isolates were identified, with 18 of the groups containing isolates originating from different environmental sources (Table 3). From all the villages, isolates genetically related to isolates from other villages could be determined.
Based on Genetic Relatedness as Determined by the Pulsed-Field Gel Electrophoresis Analysis (≥80% Homogeneity), 23 Groups of Genetically Related Isolates were Determined, 18 Out of Which (VI–XXIII) Consisted of Two or More Isolates Originating from Different Environmental Sources
BF, bird feces; DW, drinking water; F, pig feces; HW, human waste; OS, outlet sediment; RS, river sediment; RW, river water; S, soil; WW, wastewater.
Discussion
The highest detection rate of CTX-M-producing E. coli among all sample types was among pig fecal samples (73%), indicating that pig feces can be a source of wider dissemination of CTX-M-producing E. coli. A previous study on CTX-M-producing E. coli isolated from fecal samples in 21 pig farms in China reported a prevalence of 43%. 29 The detection rate of CTX-M-producing E. coli in river water in Taian city, China, was 53% in a previous study. 30 Compared with these studies, the prevalence of CTX-M-producing E. coli in pig feces and river water (64%) was higher in the study area of this study. In the rural area of Laiwu, in the Shandong province, China, the prevalence of CTX-M-producing E. coli in well water has been reported as 23%, 31 which was similar to the results in this study. In rural areas, well water is essential in daily life for activities such as tooth brushing, bathing, and as drinking water for humans (usually after boiling) and pigs. Such activities could potentially increase the risk of dissemination of CTX-M-producing E. coli from the environment to humans and animals. Notably, CTX-M-producing E. coli was also detected in vegetables (4.4%). The vegetable samples were collected from edible parts that can be eaten fresh without preparation, such as cucumbers and pepper. Through vegetables, humans and animals can be exposed to CTX-M-producing E. coli in daily life. Three isolates of CTX-M-producing E. coli (50%) originated from six human waste samples, and five isolates of CTX-M-producing E. coli originated from three samples of bird feces (two samples were isolated with four different CTX-M-producing E. coli). Results similar to ours have been reported in a study in Sweden in which CTX-M-producing E. coli could be isolated from 47% of fecal samples from wild birds. 32 Birds often live close to human settings and are exposed to our bacteria and antibiotic residues through, for example, garbage dumps, river, and agricultural areas. Thus, wild birds may be an important source and facilitator of ARB transmission.
Notably, all of the CTX-M-producing E. coli isolated in this study (n = 88) displayed the multidrug resistance phenotype, with rates of resistance to tetracycline, trimethoprim–sulfamethoxazole, and florfenicol being particularly high (94%, 82%, and 81% respectively). blaCTX-M genes may also be selected by environmental exposure to non-β-lactam antibiotics, either by being physically linked on the same plasmid as other ARGs or by being carried by ARB resistant to other antibiotics. 33 A previous study in the sampling area, conducted during the same period, showed that the various environmental compartments had levels of antibiotic contamination that may pose a risk for selection of ARB and ARGs, further emphasizing the risk of dissemination of CTX-M-producing E. coli in the local area. 34
CTX-M enzymes are the most widely disseminated ESBLs worldwide, and among clinical isolates, CTX-M-14 is the most common.35–37 CTX-M-65 has been frequently reported in E. coli isolated from humans and food-producing animals in China.38,39 In this study, 20 isolates carrying the blaCTX-M-65 gene were detected in samples from pig feces, wastewater, soil, drinking water, and river water, implying that blaCTX-M-65 has been disseminated in pigs as well as in various environmental matrices inside and outside pig farms. blaCTX-M-55 is not frequently reported in environmental isolates from China, nevertheless, the gene was detected in this study among isolates belonging to six different STs (ST88, ST206, ST1844, ST2526, ST2973, and ST5020), showing that blaCTX-M-55 has disseminated among multiple E. coli lineages in this region.
ST10 has been described from different sources worldwide, including river water, wastewater, pigs, chicken, and migratory birds.35,40–42 One study reported on the zoonotic potential of avian pathogenic isolates of E. coli belonging to the ST10 clonal complex (includes ST10 and ST48), which were associated with poultry and human cases of urinary tract infections and sepsis. 43 In this study, one isolate from wild birds belonged to ST1011. E. coli belonging to ST1011 have previously been reported both as extraintestinal pathogenic E. coli isolated from humans and as avian pathogenic E. coli isolated from birds, suggesting the zoonotic potential through wild birds of CTX-M-producing E. coli. 44 Isolates belonging to phylogroup B2 (ST2614) and phylogroup D (ST38, ST174, and ST2309) were also detected in this study. E. coli of phylogroups B2 and D can possess toxin functions and are commonly causative agents of urinary tract infections. 45 The presence of E. coli strains capable of causing urinary tract infections in the environment increases the exposure risk for humans.
CTX-M genes are a form of environmental pollution resulting from anthropogenic activities, either by direct exposure of the environment to ARGs and ARB through waste or through emergence through environmental exposure to antibiotics of subtherapeutic levels. 46 Thus, CTX-M-producing E. coli is potentially of use as an indicator to track contamination sources and dissemination of ARB and ARGs. In this study, genetic relatedness of isolates was determined by using the PFGE dendrogram (Supplementary Fig. S2), and putative dissemination routes were explored based on this phylogeny. The results showed that from all the sampled villages, isolates could be determined that were genetically related to isolates from other villages, indicating dissemination in the local geographical region (Table 3). Furthermore, 18 groups consisting of genetically related isolates of CTX-M-producing E. coli from different environmental sources were identified (Table 3). A large number of isolates with close relatives isolated from different environmental sources were identified, suggesting a widespread dissemination of CTX-M-producing E. coli between environmental matrices in the local area. Pig feces and environmental sources such as bird feces, drinking water, river sediment, river water, and wastewater had a particularly high number of isolates, which could be determined to have close relatives in other environmental matrices; from all these sources, isolates genetically related to isolates from each other of the aforementioned sources were identified (excepting that no isolates from bird feces was found that were closely related to any isolate from drinking water). These results suggest that these environmental matrices may serve as nodes through which dissemination of ARGs and ARB can occur. Although it is likely that transmission can occur in any direction from any of these environmental matrices, the methods employed in this study cannot establish the direction of transmission. Nevertheless, it is possible to speculate as to the likely routes and mechanisms of transmission (Supplementary Fig. S4).
Anthropogenic activities might modulate the environmental resistance gene pool and promote antibiotic resistance dissemination. 46 Pig farms are often flush cleaned with water, and pig fecal material can be discharged into the environment through the run-off water from the cleaning. 47 In addition, owing to the lack of management measures for livestock manure, the animal manure was directly piled up by the river in the study area. During rain, wastewater from the manure piles can be leached directly into the river, introducing large number of bacteria carrying different types of ARGs into the river water. 48 For example, a high degree of homology was observed between isolates EC6 (river water, village A), EC8 (river water, village A), and EC41 (feces, village C). These results indicated that ARB from animal feces can spread not only to environmental matrices in the surrounding aquatic environment in the same village but also may spread through the aquatic route across villages.
Rivers can likely function as vectors for ARGs and ARB transmission, either allowing the dissemination along the river to villages along its length, or to drinking water (well water) through leaching into groundwater. Previous studies have shown that environmental factors such as river direction and aerosols can affect the dissemination of ESBL genes.49–52 Another study found that presence of CTX-M-producing E. coli in river water was found to be higher in regions with large number of animals being raised than in regions with lower number of animals being raised. 53 In this study, genetically homologous isolates from river water were also observed in drinking water and wastewater of different villages, suggesting the possibility of dissemination through the river to other aquatic environments in the local area. For example, EC1 (drinking water, village A), EC22 (drinking water, village B), and EC76 (river water, village H) showed high homology in the PFGE analysis, indicating that dissemination may have occurred between river water and drinking water (well water) between different villages.
ARGs and ARB can also disseminate from river water to soil. For example, CTX-M-producing E. coli isolated from soil shared high homology with isolates collected from aquatic environments, which indicates that irrigation may facilitate dissemination of ARB between different environmental matrices (Supplementary Fig. S5). ARGs may migrate to deeper soil layers and contaminate groundwater through vertical transport. 14 Animal manure is often applied to soil as fertilizer, thus contaminating the soil with ARB present in the animal manure. 54 However, in this study, no dissemination of CTX-M-producing E. coli from pig feces to soil was indicated by the homology analysis. A possible explanation for this is that the soil and pig manure samples were collected during autumn, whereas the fields are generally fertilized during spring. A failure to observe homologous isolates in the two sample types could indicate either that the microbiota of the pigs had changed or that contaminating soil through animal manure only gives rise to a transient presence of the ARB. A previous study investigated ARGs in soils from areas along the Pearl River irrigated with wastewater, and concluded that wastewater was the primary source of antibiotics in the soil. 55
Wild birds have been hypothesized to be reservoirs for ARB, and facilitators of antibiotic resistance dissemination.35,56 In this study, CTX-M-producing E. coli isolated from wild bird feces were found to be related to isolates from animal waste, human waste, aquatic environments, and the soil, indicating that wild birds may be responsible for disseminating CTX-M-producing E. coli to these environmental matrices. Furthermore, isolates genetically related to isolates from wild birds were observed in different villages. For example, EC18, isolated from bird feces in village A, was closely related to EC36 (wastewater, village C) and EC87 (river sediment, village I). Village I is remotely located from both villages A and C, and all three villages lack a connection through river (Fig. 1). A possibility is that isolates may have spread between villages A, C, and I through birds; it had been previously shown that birds can carry CTX-M-producing E. coli from long distances while migrating and that an extensive dissemination between individual birds can allow ARB to persist in dense bird populations. 57 In addition, the isolate EC17 carrying blaCTX-M-14 gene from wild bird showed a significant genetic homology to seven isolates carrying blaCTX-M-14 gene from different matrices (Supplementary Fig. S3). This indicates that both plasmid and clonal expansion may be important modes of dissemination for CTX-M genes in the area.
Conclusion
In this study, CTX-M-producing E. coli were detected in all sampled environmental matrices, including drinking water and vegetables. The prevalence of CTX-M-producing E. coli was relatively higher in pig feces and river water than shown in previous studies. Through drinking water and vegetables from edible parts, humans and animals can be exposed to CTX-M-producing E. coli in daily life. Positive samples were distributed over several villages. Dissemination of CTX-M-producing E. coli to different environmental matrices was evaluated by analyzing the genetic relatedness of isolates from different environmental sources, and putative transmission routes through bird feces, pig feces, drinking water, river sediment, river water, and wastewater were hypothesized. Dissemination through these routes is likely facilitated by anthropogenic activities and environmental factors. Wild birds as potential vectors for dissemination of CTX-M-producing E. coli have the capacity to spread ARB across long distances. The PFGE analysis indicated that both plasmid and clonal expansion may be important modes of dissemination for CTX-M genes in the area. Regional dissemination between different environmental matrices of CTX-M-producing E. coli increases the exposure risk of humans and animals in the area.
Footnotes
Acknowledgments
The authors thank the IMPACT Consortium for its efforts in this project. This project was supported by the National Natural Science Foundation of China (Grant Nos. 41771499, 81361138021), the Fundamental Research Funds of Shandong University (Grant No. 2018JC102), the Swedish Research Council, Public Health Agency of Sweden (Grant No. D0879801), and the Swedish Research Council Formas (Grant No. 2016-00640).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.