
Abstract
Acinetobacter baumannii has emerged as a problematic multidrug-resistant (MDR) pathogen worldwide. The aim of this study was to characterize clinical isolates of A. baumannii from Jeddah, a coastal city in the western region of Saudi Arabia that has an ∼50% expatriate population. In this study, 135 nonrepetitive clinical isolates of A. baumannii were identified through MALDI-TOF and susceptibility was determined with VITEK-2 systems and broth microdilution method. Genotypic characterization of the isolates was performed by using M13 primer typing and polymerase chain reaction screening was performed for carbapenemase genes, insertion sequences, metallo-β-lactamases, and cephalosporinase genes. The isolates were recovered from heterogeneous clinical specimens, and the majority of the cases of A. baumannii infection were acquired in the hospital and predominantly involved patients who were older than 50 years. Total, 58.5% of the isolates were MDR, and 55.6% isolates were resistant to carbapenem antibiotics. Approximately half of the isolates were resistant to ceftazidime, and cefepime among the β-lactam antibiotics and ciprofloxacin from the quinolone group. The blaOXA-23-like gene and ISAba1 upstream of blaOXA-23-like were detected in 92% of the carbapenem-resistant isolates, while all carbapenem-resistant isolates were found to carry blaOXA-51-like, and blaADC-type cephalosporinase gene. The blaIMP gene was detected in 84% isolates, and two isolates carried the blaNDM-1 gene. Data demonstrate the coexistence of multiple carbapenem resistance determinants in A. baumannii from the western region of Saudi Arabia.
Introduction
Antimicrobial resistance (AMR) represents a focal point among emerging global health challenges, and experts have expressed concern that we may be entering the postantibiotics era. 1 The phenomenal therapeutic advantages offered by antibiotics are being severely threatened by the emergence of increasingly resistant strains of bacterial pathogens, and the extensive use of antibiotics both within and beyond the medical field plays a critical role in the problem. 2 Addressing AMR requires a multipronged approach that incorporates basic research on how microbes develop resistance as well as clinical trials that extend research findings to potential treatments. Resistance to therapeutically available antibiotics from all classes has been discovered in bacteria,3,4 and resistance is most frequently acquired through horizontal gene transfer. 5 Overall, the molecular basis of AMR is well studied, but its prevention and control present difficult challenges to the medical community.6,7 The phenomenon of pan-drug resistance has raised the specter of untreatable infections.
The increasing prevalence of antimicrobial-resistant Gram-negative bacteria is one of the prime threats to modern medicine. Among these bacteria, Acinetobacter baumannii, has become prominent due to the global dissemination of multidrug-resistant (MDR) lineages resistant to the carbapenem antibiotics. 8 A. baumannii is a Gram-negative, nonmotile, nonfermentative, oxidase-negative, and aerobic bacillus, which is one of the common opportunistic pathogens affecting human health. Owing to their capacity to persist on dry surfaces and their relative resistance to disinfectants, these species are able to survive well in the hospital environment. 9 Infection with A. baumannii is associated with high mortality and morbidity, including disorders such as pneumonia, bacteremia, and urinary tract, soft tissue, and skin infections, especially in patients with severe illness. 10 Acinetobacter spp. in general and A. baumannii in particular are emerging as a serious cause of health care-associated infections (HAI), especially in intensive care units. 11
The mounting number of MDR Acinetobacter species have limited the therapeutic choices for infection control. 12 In addition to its intrinsic resistance to many commonly used antibiotics, this pathogen can rapidly gain additional resistance to new broad-spectrum antibiotics.13,14 The rise of multidrug resistance, extensive drug resistance, and even pan-drug resistance is common among A. baumannii isolates. 4 More importantly, pan-drug-resistant isolates have arisen worldwide. 4 Colistin (polymyxin E) and tigecycline are frequently the only remaining antibiotics for treating MDR A. baumannii infections. 15 However, extensive resistance to the majority of antibiotics and resistance against colistin have been reported in clinical situations throughout the world. 9
The incidence of carbapenem-resistant A. baumannii has been reported within countries of the Gulf Cooperation Council, such as Saudi Arabia, Kuwait, and Bahrain.1,7 However, there is little information available on the local epidemiology, phenotypic and molecular characterization of A. baumannii from the western region of Saudi Arabia. The aim of this study was to determine antimicrobial susceptibility and investigate carbapenemase-associated resistance genes in the A. baumannii strains isolated from clinical specimens at a health care facility in western Saudi Arabia.
Materials and Methods
Sample collection
This study was performed at King Abdulaziz University Hospital (KAUH), an 845-bed major territory care hospital in Jeddah. It is the largest city in western Saudi Arabia with a population of around four million people. Approximately 50% of its residents are expatriates. We collected 135 nonduplicate A. baumannii isolates recovered from clinical specimens of the patients who visited KAUH during 2015–2016. We chose the first positive isolated sample with A. baumannii, except for two cases with a second positive bloodstream culture due to its clinical importance. If two isolates from two different sites at the same date of collection of the samples were collected, we chose the most probable source of infection. Demographic information, hospital stay, and clinical data were obtained from patients' electronic medical records following the guidelines of the ethics committee. Centers for Disease Control and Prevention/National Healthcare Safety Network (CDC/NHSN) guidelines (2013) were used to define HAI. 16 This study was approved by the Ethics Research Committee of the Faculty of Medicine at King Abdulaziz University under the reference number (235-15).
Identification and antimicrobial susceptibility screening
The purified isolates were fresh cultured on blood agar plates at 37°C for 18–20 hours using a biosafety level-2 cabinet. The identity of purified isolates was determined by Matrix Assisted Laser Desorption/Ionization Time-of-Flight (MALDI-TOF) (MALDI Biotyper Bruker Daltonics) according to the manufacturer's instructions as previously described. 17 The calibration was performed using standard Escherichia coli ATCC 8739 to validate the run. All isolates were run in duplicate and an identity score of ≥1.9 with database spectra was used to define correct identification. All isolates were tested for antimicrobial susceptibility using automated VITEK-2 (bioMérieux) system with specific AST-N291 card for Gram-negative bacteria. Antimicrobial susceptibility of A. baumannii isolates to colistin was reconfirmed by broth microdilution method using serial dilution of colistin ranging from 0.25 to 8 μg/mL. Minimum Inhibitory Concentration (MIC) results were interpreted based on the Clinical and Laboratory Standards Institute guidelines. 18
Genotyping and molecular characterization of AMR genes
Bacterial genomic DNA was extracted from A. baumannii isolates using the Wizard Genomic DNA Purification Kit (Promega) following the manufacturer's guidelines. The study isolates were typed by optimized polymerase chain reaction (PCR) with M13 primers to determine genetic relatedness as described previously. 19 Briefly, M13 PCR was performed in 25 μL volumes containing 5 μL of primer (5 pmol/μL), 12.5 μL of Go Taq Green Master Mix (Promega), and 2 μL of DNA template following previously described PCR conditions. 19 Gel electrophoresis was performed on 2% agarose gel with an initial run time of 5 minutes at 100 V in 1% TBE buffer, followed by 30 minutes at 40 V and a final run time of 70 minutes at 100 V.
Multiplex PCR was done using primers for the blaOXA-51-like, blaOXA-23-like, blaOXA-143-like, blaOXA-58-like, and blaOXA-24/40-like as described by Woodford et al. 20 The frequency of ISAba1 and ISAba4 elements upstream of blaOXA-23-like and blaOXA-51-like genes was assessed using a set of primers referred to as ISAba1F/OXA-23R, ISAba4F/OXA-23R, and ISAba1F/OXA-51R. 21 The blaADC gene, the IS element (ISAba1), and the location of the IS element in the promoter (ISAba1-ADC) of the blaADC gene were detected by PCR using the primers in Supplementary Table S1. 22 The blaIMP, blaVIM, blaSPM, blaGIM, blaSIM, blaKPC, blaNDM, blaAIM, and blaBIC genes were screened using the primers described by Poirel et al. 23 For the detection of blaNDM-1, NDM-F-38, and NDM-R-344 primers were used. 24 Screening of class 1, 2, 3 integrons was performed following the procedure described previously.25,26 The ESBL genes blaSHV, blaCTX-M, blaTEM, and blaVEB were amplified following the previously published protocol and primers sets (Supplementary Table S1).27–29 Gel-purified amplified PCR products were sequenced with ABI prism sequencer 3730 (Applied Biosystems). NCBI nucleotide BLAST was used to confirm the amplification of the respective resistance genes.
Statistical analysis
BioNumerics software (V 7.6.0) from Applied Maths was used to analyze the M13 PCR fingerprinting results, and a dendrogram was constructed using unweighted pair group method with arithmetic mean. The default parameters of 1% tolerance and 70% similarity index were considered for clustering of the isolates based on fingerprinting data.
Results
Demographic and clinical data analysis
In this study, 135 clinical isolates of A. baumannii identified by MALDI-TOF were recovered from patients visiting KAUH during 2015–2016. The average age of the patients was 47.2 ± 25.4 years (range, 0.04–93 years) with a median age of 55 years (Table 1). Relatively higher percentage of isolates were obtained from patients older than 50 years (n = 72, 53.3%) (Table 1). The strains were mainly isolated from Saudi patients (n = 63, 46.7%) and from expatriates mainly from Yemen (n = 37, 27.4%) and Palestine (n = 7, 5.2%). Twenty-eight other strains (20.7%) were obtained from patients of 14 other nationalities from Africa, Middle East, South Asia, and Southeast Asia. The strains were cultured from heterogeneous clinical specimens mostly from tracheal aspirate (n = 29, 21.5%) and blood (n = 28, 20.7%) followed by wound swab (n = 19, 14.1%) and urine midstream (n = 19, 14.1%) (Table 1). Majority of the A. baumannii infections were acquired in the hospital (n = 91, 67.4%) after 2 days patients' admission. HAI were mainly found among older patients >50 years of age (n = 56, 61.5%) and were identified mainly in tracheal aspirate (n = 24, 26.4%), blood (n = 19, 20.9%), and wound swab (n = 15, 16.5%) specimens (Table 1).
Distribution of Acinetobacter baumannii Isolates According to Demographic and Clinical Data
Others (nationality): Afghanistan, Bangladesh, Chad, Egypt, Eritrea, Ethiopia, Indonesia, Mali, Myanmar, Nigeria, Pakistan, Somalia, Sudan, Syria. Others (specimens type): eye, ear, gastric aspirate, nasopharyngeal swab, sputum, tissue, umbilical cord. Others (clinical background): anemia, arthritis, central nervous system, diabetes, fever, fracture, osteomyelitis, wound.
CAI, community-acquired infections; HAI, health care-associated infections.
A. baumannii strains were mainly isolated from HAI types of sepsis (n = 23, 25.3%), pneumonia (n = 14, 15.4%), surgical site infections (n = 8, 8.8%), and urinary tract infection (n = 8, 8.8%) (Supplementary Fig. S1). Importantly, MDR strains of A. baumannii were isolated from two cases of endocarditis. A. baumannii were recovered from patients with different clinical backgrounds that were broadly grouped into cancer, respiratory, renal, and kidney diseases (Table 1).
Antimicrobial-susceptibility analysis and genotyping
Total, 58.5% of the A. baumannii isolates tested in this study were found to be MDR, but none of them was pan-drug resistant. In total, 54.1% isolates were resistant to ≥10 tested antibiotics. Among them, 36 isolates were resistant to ≥13 tested antibiotics. The isolates, Ab15 and Ab39, recovered from blood and wound samples of Saudi and Sudanese patients, respectively, were resistant to 15 tested antibiotics. Importantly, 75 (55.6%) isolates were resistant to both meropenem and imipenem (MIC ≥8 μg/mL) from the carbapenem group of antibiotics, including one isolate that had intermediate resistance of imipenem (MIC ≥4 μg/mL) (Fig. 1). More than 60% of isolates were resistant to aztreonam, ampicillin, and ceftazidime, including isolates with intermediate resistance (Fig. 1). Furthermore, 56.3% of isolates were resistant to cefepime from the fourth-generation cephalosporin group. In the quinolone group, 57% of isolates were resistant to ciprofloxacin and 31.1% were resistant to levofloxacin, whereas 39.3% of isolates were resistant to gentamicin. The least resistance was observed against tigecycline, minocycline, tobramycin, nitrofurantoin, and ceftriaxone, with 38 isolates having intermediate resistance to tigecycline (MIC ≥4 μg/mL) and 20 isolates having intermediate resistance to minocycline (MIC ≥8 μg/mL). All isolates were sensitive to colistin (Fig. 1).
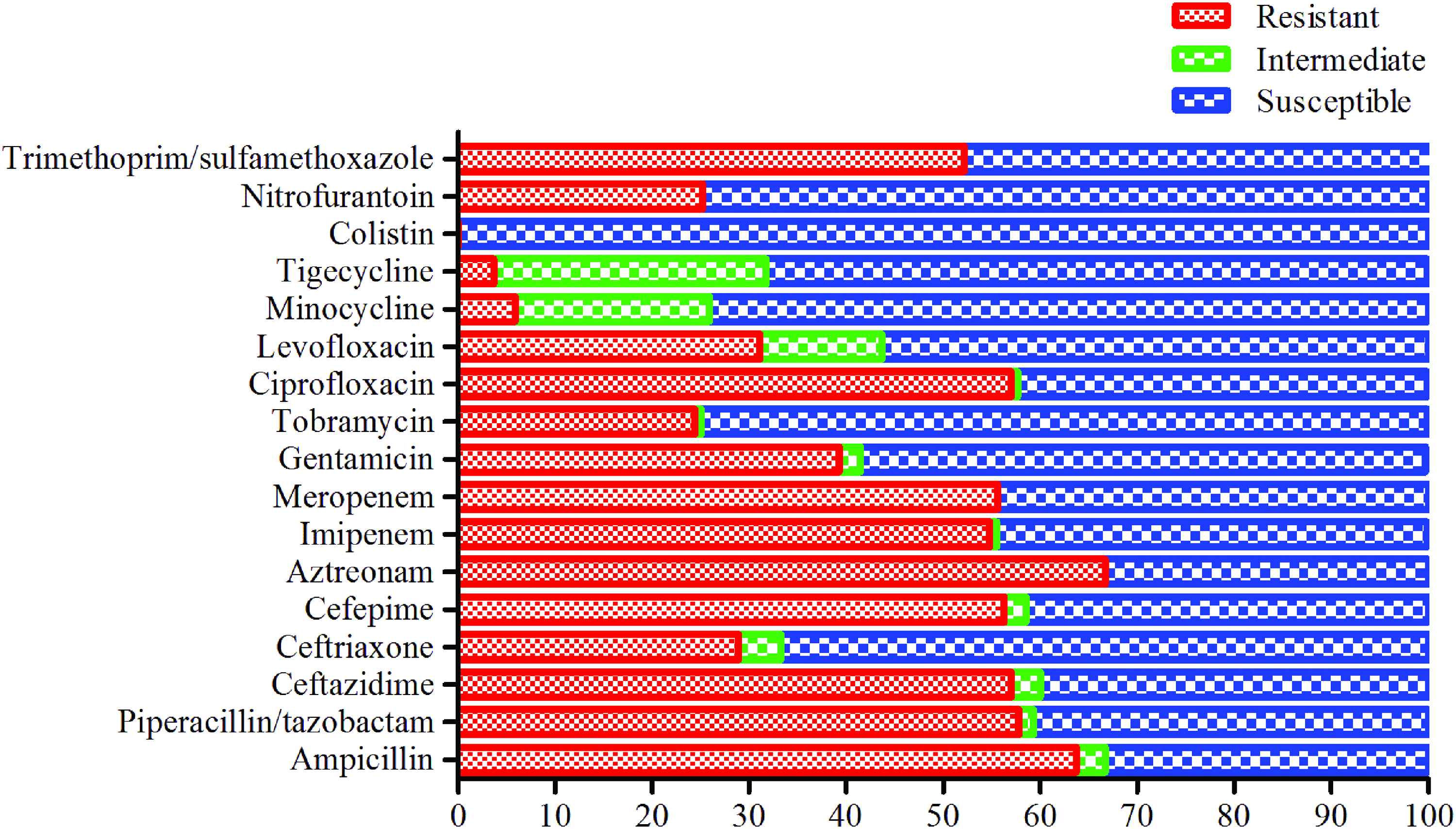
Antimicrobial resistance and susceptibility profile of the tested antibiotics among the studied Acinetobacter baumannii isolates. The x-axis values are expressed in percentage. Color images are available online.
In total, 43 different patterns (P) of antibiotic resistance were observed in 135 isolates, with resistance, including 16 group patterns and 27 singular patterns (Fig. 2). The dominant pattern (P1) of single resistance to aztreonam was identified in 26 A. baumannii isolates (Fig. 2). The second pattern of antibiotype (P2) was identified in 22 isolates, which showed resistance to 14 antibiotics. Antibiotype pattern P3 was identified in 13 isolates that harbored resistance to three antibiotics (ampicillin, ceftriaxone, and nitrofurantoin). In addition, antibiotic resistance patterns to 11 (P4) and 10 (P5) antibiotics were observed in eight isolates (Fig. 2).

Heat map of antimicrobial resistance and susceptibility patterns among 135 A. baumannii isolates. Color images are available online.
M13-PCR gave varying amplification products according to molecular weight by gel electrophoresis. Depending on the isolates, 5–20 bands were observed, and 5 isolates were not typeable by M13 primer (Fig. 3). The carbapenem-resistant isolates were divided into 10 clusters composed of 2–14 isolates at 70% similarity index, whereas 15 isolates made distinct clades in the dendrogram. Comparatively more genotypic diversity was observed in the carbapenem-sensitive isolates, which were divided into 10 clusters composed of 2–7 isolates, whereas 22 isolates made distinct clades.

UPGMA dendrogram from the bands pattern of M13-PCR that revealed the genetic relationships among the carbapenem-resistant A. baumannii isolates, along with the country (Ctry), specimen source, antibiotic resistance information, and ARGs. ARGs, antimicrobial resistance genes; CSF, cerebrospinal fluid; CSU, catheter specimen of urine; PCR, polymerase chain reaction; SPU, sputum; SW, swab; TAS, tracheal aspirate; UC, urine-catheter; UM, urine-midstream; UPGMA, unweighted pair group method with arithmetic mean.
Molecular detection of resistance genes
Most of the A. baumannii isolates carried blaOXA-type genes. All the carbapenem-resistant isolates carried blaOXA-51-like, whereas ISAba1 was found upstream of blaOXA-51-like gene in 68 (90.7%) imipenem–meropenem-resistant isolates (Figs. 3 and 4A). Whereas blaOXA-51-like was detected in 50 (83.3%) susceptible isolates, and 43 (71.7%) of them carried ISAba1 (Fig. 4B). The blaOXA-23-like gene and ISAba1 upstream of blaOXA-23-like were detected in 69 (92%) carbapenem-resistant isolates and found in five (8.3%) and three (5%) carbapenem-susceptible isolates, respectively (Fig. 4A, B). The blaOXA-24/40-like gene was detected in five isolates, including four carbapenem-resistant isolates. However, carbapenem resistance associated with blaOXA-58-like and blaOXA-143-like genes was not detected in this study.
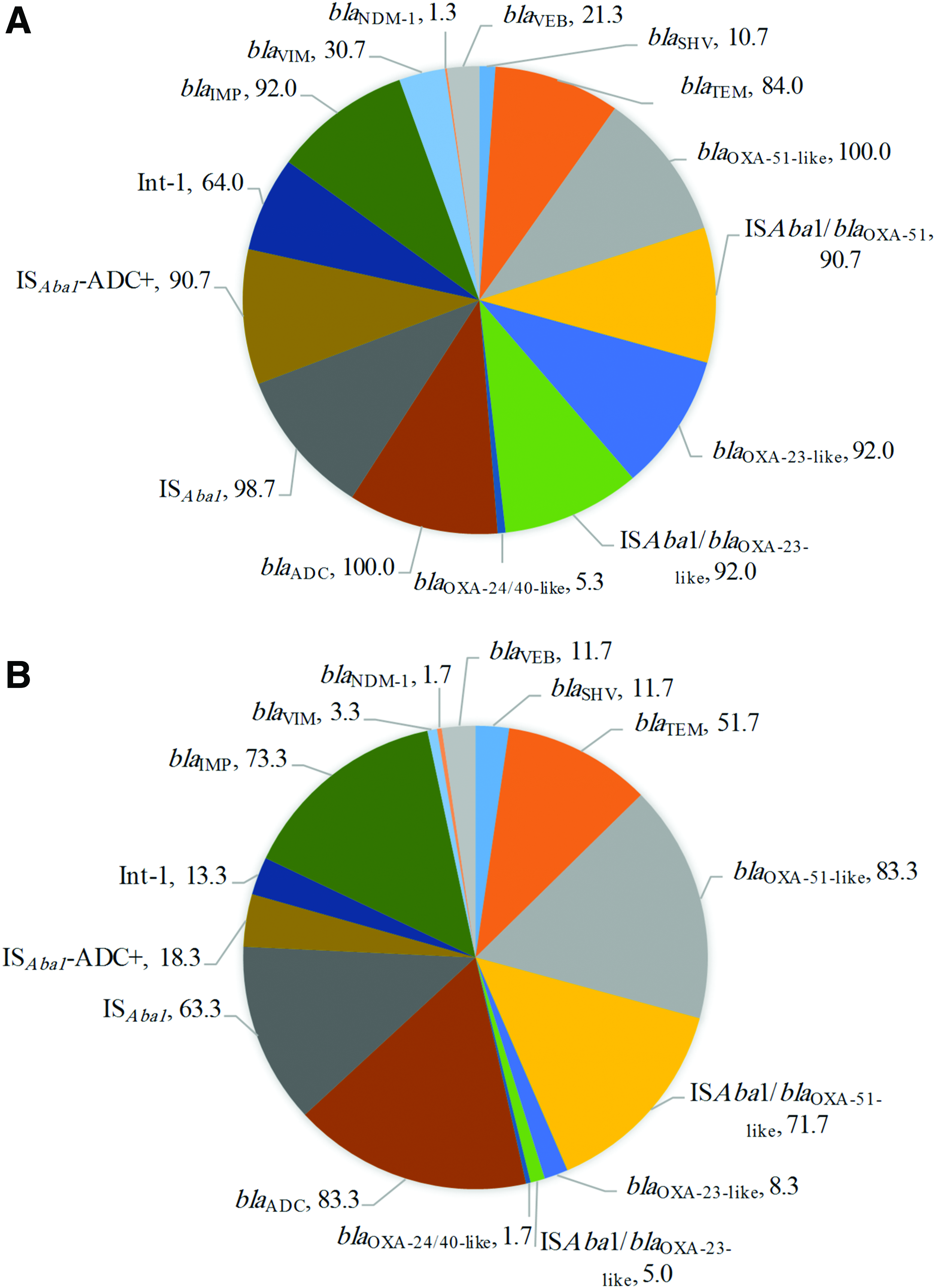
Percentage distribution of the ARGs in the
The blaADC-type cephalosporinase gene was detected in all carbapenem-resistant isolates, including 74 (99%) that were carrying IS element (ISAba1). Sixty-eight (91%) out of 75 isolates had the IS element located in the promoter region (IS Aba1 -ADC) of the blaADC gene (Fig. 4A). Fifty (83%) carbapenem-susceptible isolates carried the blaADC gene, and ISAba1 and IS Aba1 -ADC genes were found in 38 (63%) and 11 (18%) isolates, respectively (Fig. 4B). Among the acquired carbapenem-resistant genes, blaIMP was detected in 113 (84%) isolates, blaVIM in 25 (18.5%), and blaNDM-1 in 2 (1.5%). However, all isolates were negative for the following genes: blaKPC, blaSPM, blaAIM, blaGIM, blaBIC, blaSIM, and blaDIM. Of 135 A. baumannii isolates, 94 isolates (70%) harbored the blaTEM gene and 15 (10%) carried blaSHV. All isolates were negative for blaCTX-M, and blaVEB was found in 23 (17%) isolates. The presence or absence of class 1, 2, and 3 integrons among all 135 A. baumannii isolates was determined by PCR using specific primers. Our results revealed that 56 (41%) of the isolates had a class 1 integron, while class 2 and 3 integrons were not detected.
Comparative analysis of AMR genes revealed relatively high percentage of blaOXA-23-like (n = 62, 68.1%) in isolates from HAI than community-acquired infections (CAI; n = 12, 27.3%). The blaVIM and blaVEB genes were detected at relatively higher abundance of 22% (n = 20) and 20.9% (n = 19) in HAI isolates, respectively, compared with CAI isolates. Class 1 integron was detected in 46.2% (n = 42) HAI isolates and 29.5% (n = 13) CAI isolates. Supplementary Table S2 summarizes the results.
Discussion
This study assessed the antimicrobial susceptibility mechanisms of resistance to carbapenem antibiotics and local epidemiology of A. baumannii at a territory care hospital in Jeddah, Saudi Arabia. In total, 58.5% isolates were MDR and 55.6% isolates were resistant to carbapenem antibiotics that were carrying mainly blaOXA-23-like gene. We observed a high incidence of HAI caused by A. baumannii in KAUH, which is in line with previous reports.5,30 Moreover, infected patients in our study were generally older (>50 years) and may have had compromised immunity.
In this study, most of the carbapenem-resistant isolates were concurrently resistant to β-lactams other than ceftriaxone, including ampicillin, piperacillin/tazobactam, and cefepime. In addition, these isolates were more resistant to ciprofloxacin and trimethoprim/sulfamethoxazole compared with carbapenem-susceptible A. baumannii isolates. In two recent Saudi Arabian studies, the resistance rates of A. baumannii to imipenem and meropenem were 62% and 67%, respectively.14,31 In a study from China, carbapenem resistance increased from 15% for imipenem and 23% for meropenem in 2008 to 90% and 92% in 2011, respectively. 32 The prevalence of imipenem resistance in Taiwan increased from 3% in 2002 to 59% in 2010. 33 According to the SENTRY program, the resistance to imipenem ranged from 32.8% in North America to 51.7% in Latin America. 34
Consistent with a previous study from Saudi Arabia and other geographical regions, colistin was the most active antimicrobial agent against MDR isolates of A. baumannii. 35 Similarly, in studies from Bosnia and Herzegovina, all isolates were susceptible to colistin, as were 80% of isolates from Egypt.6,36 Colistin remains the last option for treating MDR A. baumannii infections. Although it has effective in vitro activity against several Gram-negative bacteria, colistin has a narrow clinical efficacy compared with β-lactam, quinolone, and aminoglycoside antibiotics because of significant neurotoxicity and nephrotoxicity. 15 Widespread use of colistin and tigecycline in treating infections caused by MDR A. baumannii may lead to increased resistance rates, exacerbating an already difficult situation.31,37 Polymyxins arose as alternative antimicrobials against A. baumannii, and the resistance rate to polymyxin B has been found to range from 2% in North America to 0.9% in Europe. 34
Generally, blaOXA-51-like and blaADC genes are the most prevalent resistance genes, and these genes were detected in around 97% of our A. baumannii isolates, irrespective of the carbapenem susceptibility or resistance. The detection of blaOXA-51-like genes in 94% of isolates in this study is consistent with previous reports that they are intrinsic to A. baumannii.13,20 The blaOXA-51-like genes not being found in 6% of the A. baumannii isolates may be due to variation in sequence at the primer binding site or to the presence of other variants of blaOXA-51-like in those isolates. 38 ISAba1 has been suggested to provide the promoter for bla genes, and it is associated with carbapenem resistance. 38 The results of this study revealed the presence of ISAba1 upstream of blaOXA-51-like among susceptible and nonsusceptible A. baumannii isolates, indicating that this IS element may not influence the regulation of blaOXA-51-like gene.
The blaOXA-23-like gene was the first carbapenemase OXA detected in A. baumannii. 39 In this study and other studies from the eastern region of Saudi Arabia, blaOXA-23-like was found in 55%–94% of A. baumannii isolates.7,8,31 In a study from China, Fu et al. found dissemination of blaOXA-23-like in carbapenem-positive A. baumannii isolates in multiple cities.40,41 Our study showed that the presence of ISAba1-blaOXA-23 was sufficient to confer resistance even without the backing of other oxacillinase genes. However, detection of ISAba1-blaOXA-23 in some isolates with carbapenem susceptibility might be due to the downregulation of the IS element in those isolates or gene truncation. 42 Production of OXA-23 is the most commonly encountered mechanism of carbapenem resistance in A. baumannii globally. 43 The OXA-23-like enzymes that contribute to carbapenem resistance are encoded by blaOXA-23-like genes present on plasmids or chromosomes and are associated with the presence of ISAba1. The carbapenemases originated on plasmid are easily spread to other bacterial species and can occur in the hospital environments, which highlights the importance of developing control strategies to prevent infections caused by carbapenem-resistant A. baumannii.
Detection of blaVIM in 19% isolates described that metallo-β-lactamases were not the dominant β-lactamases in the clinical A. baumannii isolates from KAUH, but 84% of A. baumannii strains contained blaIMP and 70% carried blaTEM genes. Screening for the blaVEB genotype revealed that 17% of our isolates carried this gene, which was previously reported in Iran (10%) and the United States (46.71%). 44 Our results indicate that more than 80% of A. baumannii isolates harboring ISAba1 and blaADC were phenotypically resistant to ceftazidime and cefepime. However, several A. baumannii isolates with ISAba1-blaADC remained susceptible to ceftriaxone from among the third-generation cephalosporins. Different A. baumannii strains likely have differences in transcription-level regulation of blaADC that may affect the cephalosporin susceptibility. 45 In this study, blaOXA-58-like and blaOXA-143-like genes were absent in our isolates, in contrast to previous reports from Saudi Arabia that found a low prevalence of the respective genes.7,8,31,46 In Saudi Arabia, the blaOXA-24/40-like gene was detected at a rate of 4%-45% in A. baumannii isolates. 7 Similarly, in this study five isolates were blaOXA-24/40-like positive. Importantly, two isolates were found to be blaNDM-1 positive. In 2012, an A. baumannii isolate carrying blaNDM-1 was found in Buraidah, located in the north-central region of Saudi Arabia. 47 El-Mahdy et al. reported blaNDM-1 isolates in the eastern region of Saudi Arabia in 2014. 48 These data about the possible prevalence of this successful carbapenemase in A. baumannii isolates in Saudi Arabia are alarming. The Indian subcontinent is the main reservoir for NDM producers, and recent studies suggest that the Middle East region might be a secondary reservoir. 3
Conclusions
This study highlighted the high prevalence of carbapenem resistance among A. baumannii isolates in the health care facilities of Saudi Arabia. The risk of swift dissemination of carbapenem-resistance mechanisms in our study might be through IS-OXA-23-like carbapenemase in the A. baumannii isolates. However, variation exists in the prevalence and distribution of carbapenem-associated genes in the different regions of Saudi Arabia. A national surveillance program is needed to monitor the rapid dissemination of carbapenem-resistant A. baumannii and the associated risk factors in health care facilities to adopt effective policies for control measures.
Footnotes
Acknowledgments
This project was funded by the National Plan for Science, Technology, and Innovation (MAARIFAH)—King Abdulaziz City for Science and Technology—the Kingdom of Saudi Arabia—award number (12MED3108-03). The authors also acknowledge with thanks the Science and Technology Unit, King Abdulaziz University for their technical support.
Authors' Contributions
M.W.S., and M.Y. contributed to the experimental work, data analysis, and writing of the article. M.F. and S.B.A. contributed to the experimental work. M.A. and D.E. contributed to the sample collection. A.A.J.-F. and E.I.A. contributed to the study design and reviewed the article.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.