
Abstract
Alteration in two-component systems (TCSs), which are signal transduction pathways in prokaryotes, can result in antibiotic resistance. Recently, it has been shown that the overexpression, using a multicopy cloning vector, of the dcuR, rcsB, and yehT genes, which code for the response regulator (RR) part of TCSs, enhanced the minimal inhibitory concentrations (MICs) of carbapenems in Escherichia coli K-12 derivative KAM3. Herein, the contribution to carbapenem resistance of the DcuS/DcuR, RcsC/RcsB, and YehU/YehT TCSs was assessed in E. coli K-12 derivative BW25113 (A phylogroup) and 536 (B2 phylogroup) recipient strains in combination with extended-spectrum β-lactamase that exhibit a weak carbapenemase activity. The genes encoding both the sensor kinase (SK) and the RR, on the one hand, and the genes encoding the SK only, on the other hand, of these regulating pathways were disrupted. Subsequently, the mutants and their parental strains were transformed by a recombinant plasmid encoding the CTX-M-15 gene, before testing their susceptibility to carbapenems and their fitness. Results showed a trade-off between enhanced MICs for ertapenem, which remained above the clinical resistance breakpoint, and decreased growth rate, specifically for the 536 strain SK mutants. In conclusion, mutations in dcuS/dcuR, rcsC/rcsB, and yehU/yehT genes may be a pivotal first-step event in the development of carbapenem resistance.
Introduction
Infections due to extended-spectrum β-lactamase (ESBL)-producing Escherichia coli are usually treated by carbapenems. Some isolates have developed carbapenem resistance, which resulted in limited options for the treatment of infections caused by these organisms. Decreased bacterial cell permeability due to loss or alteration of the outer membrane porins OmpF and OmpC in combination with ESBL production constitutes one of the main mechanisms of carbapenem resistance in ESBL-producing E. coli. Indeed, lack of permeability can magnify the hydrolytic activity of ESBL against substrates that are poorly hydrolyzed.1,2
Two-component systems (TCSs) consist of two types of signal transducers: a transmembrane sensor kinase (SK), which detects chemical and/or physical signals, and its cognate cytoplasmic response regulator (RR), which modulates gene transcription. 3 Signals are transduced from SK to RR through phosphotransfer reactions. Upon activation, SK converts the external stimulus into an adaptive signal by autophosphorylation at a conserved histidine residue. The phosphorylated histidine is the resource for phosphorylation of a conserved aspartic acid residue in the receiver domain of the RR. Unphosphorylated SK acts as a phosphatase that removes phosphoryl groups from RR, thereby maintaining the equilibrium between active and inactive state. 3
It has been shown that the alteration of some TCSs could affect the permeability, thus increasing the minimal inhibitory concentrations (MICs) of carbapenems. Indeed, inactivation of the EnvZ/OmpR TCS was responsible for ertapenem resistance in an ESBL-producing E. coli clinical isolate. 2 Moreover, Hirakawa et al. showed that the overexpression, using a multicopy cloning vector, of the dcuR, rcsB, and yehT genes, which code for the RR parts of TCSs, enhanced the MICs of carbapenems in E. coli K-12 derivative KAM3. 4
The aim of our study was to disrupt the chromosome-borne genes encoding the DcuS/DcuR, RcsC/RcsB, and YehU/YehT TCSs to assess their contribution to carbapenem resistance in E. coli in combination with the CTX-M-15 β-lactamase production, which is one of the most widespread ESBL, and to determine their impact on fitness.
The well-characterized E. coli K-12 derivative BW25113 and E. coli uropathogenic 536 strains, which belong to the phylogenetic groups A and B2, respectively, were used.5,6 TCS inactivation was performed by deleting simultaneously the genes encoding the SK and the RR, or by disrupting only the gene coding for the SK. Gene inactivation experiments were carried out using the technique described by Datsenko and Wanner, 7 with the primers listed in Supplementary Table S1.
It gave rise to the double mutants E. coli BW25113 ΔdcuSΔdcuR, E. coli BW25113 ΔrcsCΔrcsB, E. coli BW25113 ΔyehUΔyehT, E. coli 536 ΔdcuSΔdcuR, E. coli 536 ΔrcsCΔrcsB, and E. coli 536 ΔyehUΔyehT and to the single mutants E. coli BW25113 ΔdcuS, E. coli BW25113 ΔrcsC, E. coli BW25113 ΔyehU, E. coli 536 ΔdcuS, E. coli 536 ΔrcsC, and E. coli 536 ΔyehU, respectively. Polymerase chain reaction (PCR) amplification of the genetic environment surrounding the target genes and subsequent sequencing of the PCR products was carried out to confirm the gene disruption.
All the E. coli BW25113 and E. coli 536 mutants and their parental strains were subsequently transformed with the pCTX-M-15 recombinant plasmid, which was obtained by cloning the PCR product of the entire blaCTX-M-15 gene from E. coli ErtS using the Zero Blunt TOPO PCR cloning kit (Thermo Fisher Scientific, Courtaboeuf, France). 2 The orientation of the cloned insert was under the transcriptional control of the lacZ promoter flanking the cloning site, as confirmed by sequencing. MIC values of imipenem and ertapenem were determined using E-test (bioMérieux, Marcy-l'Etoile, France) and the Clinical and Laboratory Standards Institute (CLSI) agar dilution method. 8 A twofold increase in the MICs of the mutant strains as compared with their parental strain was considered significant.
The deletion of the gene encoding the SK in association with CTX-M-15 production increased significantly the MICs for ertapenem only in E. coli 536 recipient strain. However, the MICs remained above the clinical resistance breakpoint (Table 1). On the opposite, the deletion of the gene encoding the SK in the E. coli BW25113 recipient strain and the disruption of the genes encoding SK and RR in both strains failed to increase significantly the MICs for carbapenems even in combination with CTX-M-15 production (Table 1).
MIC Values for the Escherichia coli K-12 BW25113 and E. coli 536 Mutants, and Their Parental Strains, Expressing or not Expressing the CTX-M-15 Extended-Spectrum β-Lactamase
Bold values indicate MICs for parental strains.
Production of CTX-M-15 was confirmed by a positive synergy test between amoxicillin/clavulanic acid and extended-spectrum cephalosporin disks. A twofold increase in MIC as compared with the parental strain was considered significant. Significant modifications of MIC are underlined.
MIC, minimal inhibitory concentrations.
To assess the fitness cost induced by the change of TCS activity, the bacterial growth of the mutant strains and their parental isolates was measured in lysogeny broth (LB) (Difco; Fischer Scientific, Illkirch, France) medium by monitoring the optical density at 600 nm (Tecan microplate reader [Tecan, Lyon, France]), as described previously. 2 For each strain and condition, maximal growth rate (MGR) was measured in duplicate in five separate experiments and were compared by nonparametric tests (Mann–Whitney and Kruskal–Wallis) using the R software. Deletion of SK and RR-encoding genes decreased significantly the MGR in E. coli BW25113 (Fig. 1A), whereas deletion of SK-encoding genes decreased it in E. coli 536 (Fig. 1B). However, no competition assay was performed to screen for fine-scale variation in fitness of E. coli 536 mutants lacking both SK- and RR-encoding genes.
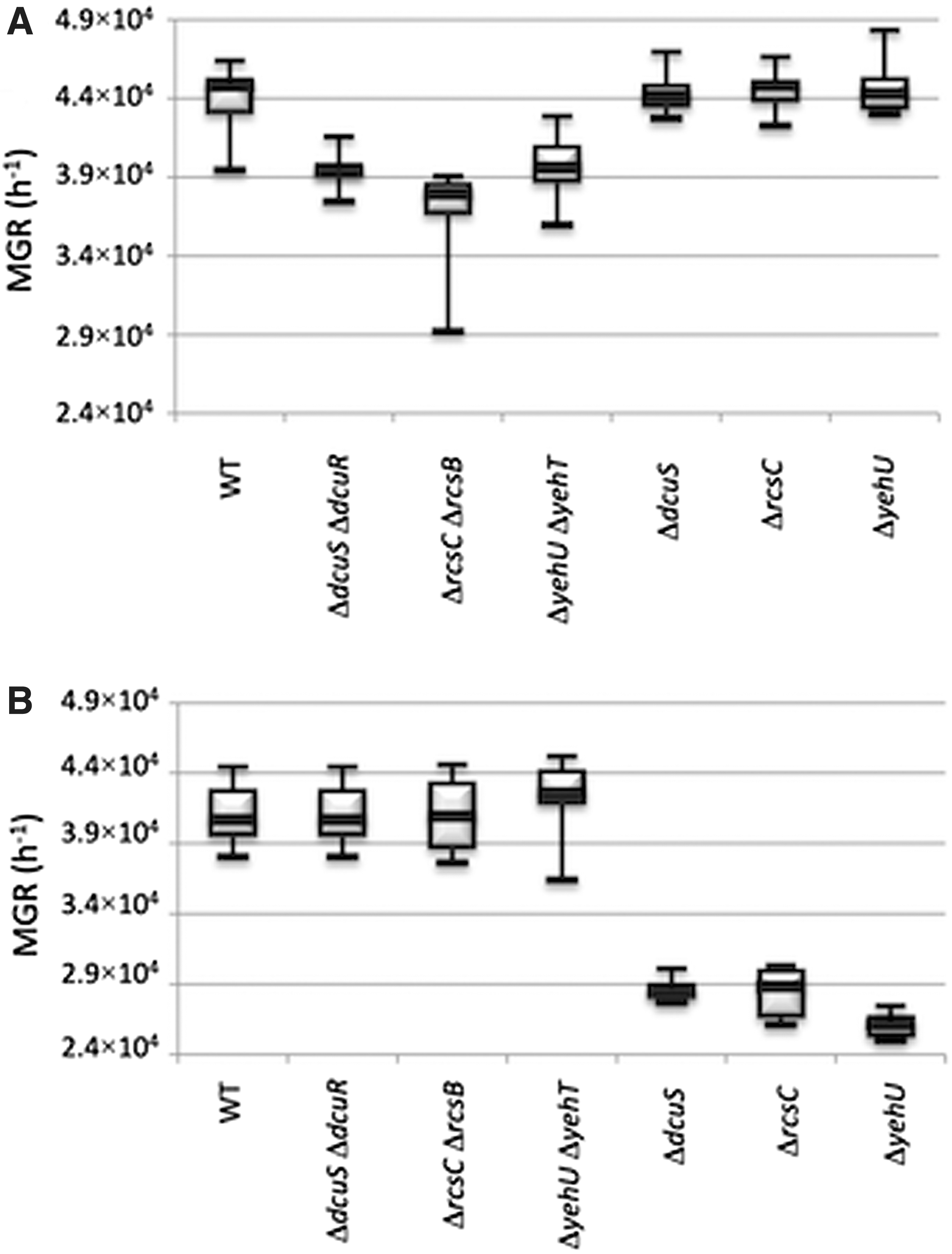
MGRs of E. coli K-12 BW25113
Alteration in some TCSs has been previously described as a source of antibiotic resistance. Changes in the expression of OmpR/EnvZ, CpxA/CpxR, and PhoP/PhoQ systems may confer carbapenem, 2 aminoglycosides, 3 and colistin resistance, 9 respectively. Herein, we showed that disruption of the dcuS, rcsC, and yehU genes accounted for a weak increase of the MICs for ertapenem in E. coli 536 (pCTX-M-15), whereas total inactivation of the Dcu, Rcs, and Yeh TCS in this strain, by deleting simultaneously SK- and RR-encoding genes, reduced MICs for ertapenem slightly. This effect could be ascribed to the loss of the phosphatase activity of the SK after specific disruption of the encoding gene. The absence of phosphatase activity might upregulate the TCS response owing to the unrestrained phosphorylation of RR by small-molecule phospho donors or heterologous kinases, as described previously for the CpxA/CpxR TCS. 3 Further experiments are warranted to ascertain this hypothesis.
It is also noteworthy that the disruption of the SK-encoding dcuS, rcsC, and yehU genes in E. coli 536 (pCTX-M-15) only weakly affected the MIC for ertapenem, whereas that for imipenem remained unchanged. In contrast, Hirakawa et al. showed that overexpression of RR-encoding dcuR, rcsB, and yehT genes, using an isopropyl β-D-1-thiogalactopyranoside (IPTG)-induced multicopy expression vector, led to a slight increase of the MIC of imipenem, which still remained below the clinical breakpoint. 4 This discrepancy could be attributable to the methodology used: we abolished the negative feedback activity of SK, thus mimicking the physiological upregulation of TCSs, whereas Hirakawa et al. transformed the E. coli K-12 derivative KAM3 by a multicopy expression vector harboring the RR-encoding genes before inducing their overexpression by IPTG. 4
In addition, the increased MICs for ertapenem in E. coli 536 (pCTX-M-15) was counterbalanced by a reduction of the growth rate. Such negative correlation between antibiotic resistance and fitness cost was already pointed out by Phan and Ferenci who showed that membrane permeability and competitive fitness are linked by a trade-off between self-preservation and nutritional competence. 10
Interestingly, the weak increase of MIC for ertapenem induced by SK-encoding gene disruption only occurred in E. coli 536 (pCTX-M-15) phylogroup B2 strain and not in E. coli BW25113 (pCTX-M-15) phylogroup A strain. This difference could be attributable to genome epistasis, which is a phenomenon where the effect of a mutation is dependent of the E. coli genetic background, as described by Tenaillon et al. 11 The contribution of epistasis to acquired resistance in E. coli had already been demonstrated in a study on the structure–activity relationship of chromosome-borne AmpC β-lactamases (cephalosporinase) of E. coli. It showed that structural alteration in the H-10 helix of the enzyme led to an important increase in the catalytic efficiency toward extended-spectrum cephalosporins in the AmpC β-lactamases of a phylogenetic group A strain only. 12
It is noteworthy that the increase of the MIC for ertapenem, which resulted from the alteration of the DcuS/DcuR, RcsC/RcsB, and YehU/YehT TCSs in combination with CTX-M-15 production, remained above the clinical resistance breakpoint, whereas the association between OmpR/EnvZ alteration and CTX-M-15 production was previously shown to confer ertapenem resistance. 2 This difference could result from the diverse mechanisms of resistance regulated by these transduction pathways. Although OmpR/EnvZ modulates the expression of ompF and ompC genes, which code for the main outer membrane porins in E. coli, 2 several studies have shown that DcuS/DcuR, RcsC/RcsB, and YehU/YehT play an important role in the remodeling of the bacterial surface during growth on a solid surface and biofilm formation. 13 The rcsB gene overexpression also increased the mRNA levels of the acrD and macA genes, which code for efflux transporters. 4 In addition, Hirakawa et al. stated that RcsB and DcuR should not repress ompF expression since these TCSs did not confer resistance to lipophilic compounds, which actually pass through OmpF porin. 4
In our study, the weak increase of the MICs for ertapenem is offset by a decreased fitness in LB. However, we should not overlook the mechanisms of low-level resistance in microorganisms, as they can serve as stepping-stones to develop high-level clinically relevant resistance. 14
In conclusion, although mutations in dcuS/dcuR, rcsC/rcsB, and yehU/yehT genes do not seem to confer clinical resistance to carbapenem treatment to ESBL-producing E. coli strains, they may be a pivotal first-step event in the development of carbapenem resistance during therapy, which subsequently may predispose to antibiotic treatment failure.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.