
Abstract
Nemonoxacin is a nonfluorinated quinolone with good bactericidal effects against quinolone-resistant Gram-positive microorganisms. The in vitro inducible resistance of nemonoxacin against clinically relevant Gram-positive pathogens was compared with ciprofloxacin, levofloxacin, and moxifloxacin. Three strains of each bacterial species, including Streptococcus pneumoniae, Staphylococcus aureus, Enterococcus faecium, and Enterococcus faecalis, were cultured. All clinical isolates had wild-type gyrA, gyrB and parC, parE before further in vitro test. DNA sequencing for the quinolone resistance determination region (QRDR) of gyrase and topoisomerase genes was performed. Nemonoxacin had the lowest minimum inhibitory concentrations (MICs) among all quinolones. During exposure to nemonoxacin, the MIC values did not increase for S. aureus, E. faecium, and E. faecalis, and revealed fourfold increase of S. pneumoniae over three cycles of a stepwise resistance selection. DNA sequencing did not show inducible QRDR resistance of nemonoxacin group. Compared to other fluoroquinolones, nemonoxacin has a low potential for inducing resistant pathogens.
Introduction
Pulmonary infection is a major infection causing the morbidities and mortality of human diseases.1,2 The widespread use of beta-lactams increased the prevalence and spread of resistant strains. Fluoroquinolones have been utilized as appropriate therapeutic substitutes, because of their good potency, oral bioavailability, pharmacokinetics, and pharmacodynamic profile. 3 However, resistance to fluoroquinolones is also on the rise. 4 Fluoroquinolone resistance in Streptococcus pneumoniae and Staphylococcus aureus is generally related to spontaneous point mutations in the quinolone resistance-determining regions (QRDRs) of the target genes, including parC and parE, which encode topoisomerase IV, and gyrA and gyrB, which encode DNA gyrase. 5
Mutations in parC and gyrA have been the key determinant of fluoroquinolone resistance in S. pneumoniae, and several stepwise mutations in all four of the target genes have made some of the most efficacious fluoroquinolones, such as levofloxacin and moxifloxacin, ineffective against resistant isolates of pneumococci. 6 The mutations selected by nemonoxacin were found to be limited to gyrA, gyrB, and parE, and spared parC, which is known to play an important role in resistance in clinical isolates of S. pneumoniae. 7
Nemonoxacin is a newly designed compound of C-8-methoxy nonfluorinated quinolone (NFQ). The combination of a methoxy group at the C-8 position facilitates nemonoxacin to target both topoisomerase IV and DNA gyrase, associated with an improved spectrum of activity and reduced mutant selection.7,8 Nemonoxacin has shown excellent potency against quinolone-resistant Gram-positive pathogens, including methicillin-resistant S. aureus (MRSA) and quinolone-resistant S. pneumoniae. This special structure resulted in the generation of NFQs, which did not show loss of potency due to hotspot mutations of QRDR in parC.8,9 In our study, a stepwise resistance selection assay with general resistant pathogens, such as S. pneumoniae, S. aureus, and Enterococcus, will be utilized to assess in vitro development of resistance to nemonoxacin, in comparison to ciprofloxacin, levofloxacin, and moxifloxacin.
Materials and Methods
Clinical bacterial strains
Three strains of each Gram-positive species, including S. pneumoniae (one penicillin-resistant S. pneumoniae strain), S. aureus (one MRSA strain), Enterococcus faecium (one vancomycin-resistant E. faecium strain), and Enterococcus faecalis (one ampicillin-resistant E. faecalis strain), were cultured from patient specimens for in vitro resistance selection studies at National Taiwan University Hospital, Taiwan. The genotypes of gyrA, gyrB and parC, parE in all clinical isolates were confirmed to be wild type before further in vitro laboratory examinations.
Stepwise resistance selection
Four respiratory quinolones, including nemonoxacin, levofloxacin, ciprofloxacin, and moxifloxacin, were compared in the stepwise resistance selection test. 10 Stepwise resistance selection studies were executed at 1 × , 2 × , and 4 × minimum inhibitory concentration (MIC) for each step of mutant selection in 100 mm Petri plates containing 20 mL Mueller-Hinton Agar II with 5% sheep blood. Agar plates were manipulated 24 hr before inoculation, and they cooled at room temperature with a mantle by aluminum foil to reduce illumination. Bacterial inoculum was generated by suspending 24-hr bacterial colonies, which were grown on Tryptic Soy Agar II agar, in saline to an optical density of 1.0 at 600 nm. Four plates of each concentration of tested antibiotics were inoculated with 250 μL of bacterial suspension, with a total number of cells ranging from 1 × 109–1 × 1010 CFU. The bacterial suspension was spread for confluent growth all over the agar surface and used to calculate the fold increase in MIC.
DNA sequencing
The QRDRs of the target genes, such as parC, parE, gyrA, and gyrB, were examined by DNA sequencing. The primers used in our study listed in Table 1 were obtained from Sigma (Sigma-Aldrich Co., Darmstadt, Germany). Platinum-PCR Supermix high fidelity and a low DNA mass ladder were gathered from Invitrogen. We used a PCR purification kit, QIAquick, from QIAgen. DNA sequencing was executed based on a previous report. 11 Bacterial DNA was prepared by suspending one colony of each isolate in 100 μL TE buffer (10 mM Tris HCl, pH 8.0, and 1 mM EDTA) for sequencing the QRDR of gyrase and topoisomerase IV genes. PCR amplification mixtures contained 90 μL high fidelity Super-mix, 4 μL of the reverse and forward primers, and 4 μL of DNA template. PCR steps included preheating at 94°C for 5 min, then 30 cycles of 94°C for 40 sec, then 55°C for 40 sec, and then 72°C for 1 min, and a final incubation of 72°C for 5 min.
Primers Used for Quinolone Resistance Determination Region in This Study
The references of primers used for quinolone resistance determination region in this study were Enterococcus faecium [11], Staphylococcus aureus [12], Enterococcus faecalis [13], and Streptococcus pneumoniae [14].
EM, E. faecium; SA, S. aureus; EL, E. faecalis; SP, S. pneumoniae.
+, sense primer; −, antisense primer.
Amplified DNA was purified by QIAgen QIAquick spin column method, and it was quantified by DNA electrophoresis beside a low mass ladder in a 1% agarose gel. Sequencing was done on an ABI Prism 3100 Genetic analyzer (Applied Biosystems). DNA sequencing of the QRDRs of the targeted genes were done on two isolates of each bacterial strain with elevated MICs after every step of selection. One isolate from the two genetically characterized isolates was chosen for repeat exposure to antibacterials in the second step of resistance selection. Isolates selected for further steps of resistance selection were chosen on the basis of having the most genotypic changes in the QRDR sequence. Similarly, an isolate after the second step of resistance selection was chosen for repeat exposure to antibacterials in step 3.
Ciprofloxacin was used as a control in all experiments. Sequence fragments were compared to the complete sequence through a BLAST search (BLAST; www.ncbi.nlm.nih.gov/BLAST). Mutations were identified visually and compared to hotspot mutations described in the literature.
Results
The in vitro antimicrobial susceptibility testing results of ciprofloxacin, levofloxacin, moxifloxacin, and nemonoxacin against each of these three different strains of S. aureus, E. faecium, E. faecalis, and S. pneumoniae are shown in Table 2. Nemonoxacin showed the lowest MICs among all quinolones. For instance, the MICs of three S. aureus strains were 0.03, 0.06, and 0.03 μg/mL for nemonoxacin, compared with 0.016, 0.125, and 0.047 μg/mL for moxifloxacin, 0.125, 0.19, and 0.19 μg/mL for levofloxacin, and 0.19, 0.38, and 0.25 μg/mL for ciprofloxacin, respectively.
Inducible In Vitro Antimicrobial Susceptibility Test of Ciprofloxacin, Levofloxacin, Moxifloxacin, and Nemonoxacin Against Three Strains Each of Staphylococcus aureus, Enterococcus faecium, Enterococcus faecalis, and Streptococcus pneumoniae
The alphabet numbers represent the MIC values in μg/mL of isolates.
NA, not available to get MIC data due to no growth.
Low resistance development of nemonoxacin was observed after increasing higher concentrations of quinolone exposure. The MICs of nemonoxacin did not increase for S. aureus, E. faecium, and E. faecalis and fourfold increase of S. pneumoniae after three cycles of selection. In contrast, the MIC of ciprofloxacin had 64-fold to more than 128-fold elevation after three selection cycles in S. aureus strains, followed by E. faecium strains (3–32-fold), and S. pneumoniae (40–64-fold increase). The MICs of ciprofloxacin against E. faecalis remained stationary. The antimicrobial susceptibility test revealed intermediate activities of increasing MIC levels in levofloxacin and moxifloxacin. In conclusion, our in vitro resistant selection studies revealed nemonoxacin had a low propensity for selecting resistant pathogens compared to other fluoroquinolones.
The results of QRDR of the target genes are shown in Tables 3–6. We could find several mutations in QRDR regions for the isolates after exposure to ciprofloxacin (Table 3). S378-G mutation could be noted in parE of E. faecium. For comparison, T414-M mutation could be identified in parE of S. aureus, and the mutations with S80-F in parC and S84-I/L in gyrA were also found. Among the resistant isolates of S. pneumoniae, the amino acid substitutions with S79-Y and C532-E were noted. The isolates of E. faecalis exposed to ciprofloxacin did not develop any QRDR mutations.
Mutations Identified in the Quinolone Resistance-Determining Regions and the Minimal Inhibitory Concentrations for Ciprofloxacin Selected Clinical Isolates
EF3 was vancomycin-resistant E. faecium strain. SA3 was methicillin-resistant S. aureus strain. EL3 was ampicillin-resistant E. faecalis strain. SP3 was penicillin-resistant S. pneumoniae strain.
QRDR, quinolone resistance-determining regions.
Mutations Identified in the Quinolone Resistance-Determining Regions and the Minimal Inhibitory Concentrations for Nemonoxacin Selected Clinical Isolates
EF3 was vancomycin-resistant E. faecium strain. SA3 was methicillin-resistant S. aureus strain. EL3 was ampicillin-resistant E. faecalis strain. SP3 was penicillin-resistant S. pneumoniae strain.
The MICs of the bacterial isolates were lower for levofloxacin than for ciprofloxacin. However, several induced mutations in QRDR were documented (Table 4). The mutations with H486-Y and D488-G in parE, and V450-I, R500-S, and Y504-D in gyrB were found in E. faecium. The mutations with S80-Y in parC and G433-E in gyrB were noted among two isolates of S. aureus with lower MICs against levofloxacin. However, E84-K was identified in the S. aureus isolate, which had a higher MIC (>32 μg/mL) for levofloxacin. Only an isolate of E. faecalis had an amino acid substitution of D40-V. Mutations in QRDR genes from S. aureus after exposure to levofloxacin did not occur alone. A combination of mutations on the same or different gene targets could be observed. Therefore, we cannot attribute the elevation of MIC to single mutation, but to the combination of those. There was no mutation for all isolates of S. pneumoniae against levofloxacin.
Mutations Identified in the Quinolone Resistance-Determining Regions and the Minimal Inhibitory Concentrations for Levofloxacin Selected Clinical Isolates
EF3 was vancomycin-resistant E. faecium strain. SA3 was methicillin-resistant S. aureus strain. EL3 was ampicillin-resistant E. faecalis strain. SP3 was penicillin-resistant S. pneumoniae strain.
The isolates after exposure to moxifloxacin had shown only mildly elevated MICs. There were some substitutions, such as A442-P, R457-P, A466-T, S467-T, E475-D, K501-Q, and V503-C, in gyrB for E. faecium (Table 5). Two mutations with A116-E and R159-K in parC were found in one strain of S. aureus, which had a higher MIC against moxifloxacin. There was no amino acid substitution under the selective pressure of moxifloxacin for E. faecalis and S. pneumoniae.
Mutations Identified in the Quinolone Resistance-Determining Regions and the Minimal Inhibitory Concentrations for Moxifloxacin Selected Clinical Isolates
EF3 was vancomycin-resistant E. faecium strain. SA3 was methicillin-resistant S. aureus strain. EL3 was ampicillin-resistant E. faecalis strain. SP3 was penicillin-resistant S. pneumoniae strain.
For nemonoxacin, we did not find any identifiable amino acid changes in the QRDR regions of the target genes (Table 6). To make a conclusion, nemonoxacin had the lowest possibility of inducing resistant QRDR mutations in the target genes (parE, parC, gyrA, and gyrB) among these four tested quinolones, and it as well as moxifloxacin maintained the lowest MICs for the strains that underwent the stepwise resistance selection (Fig. 1).
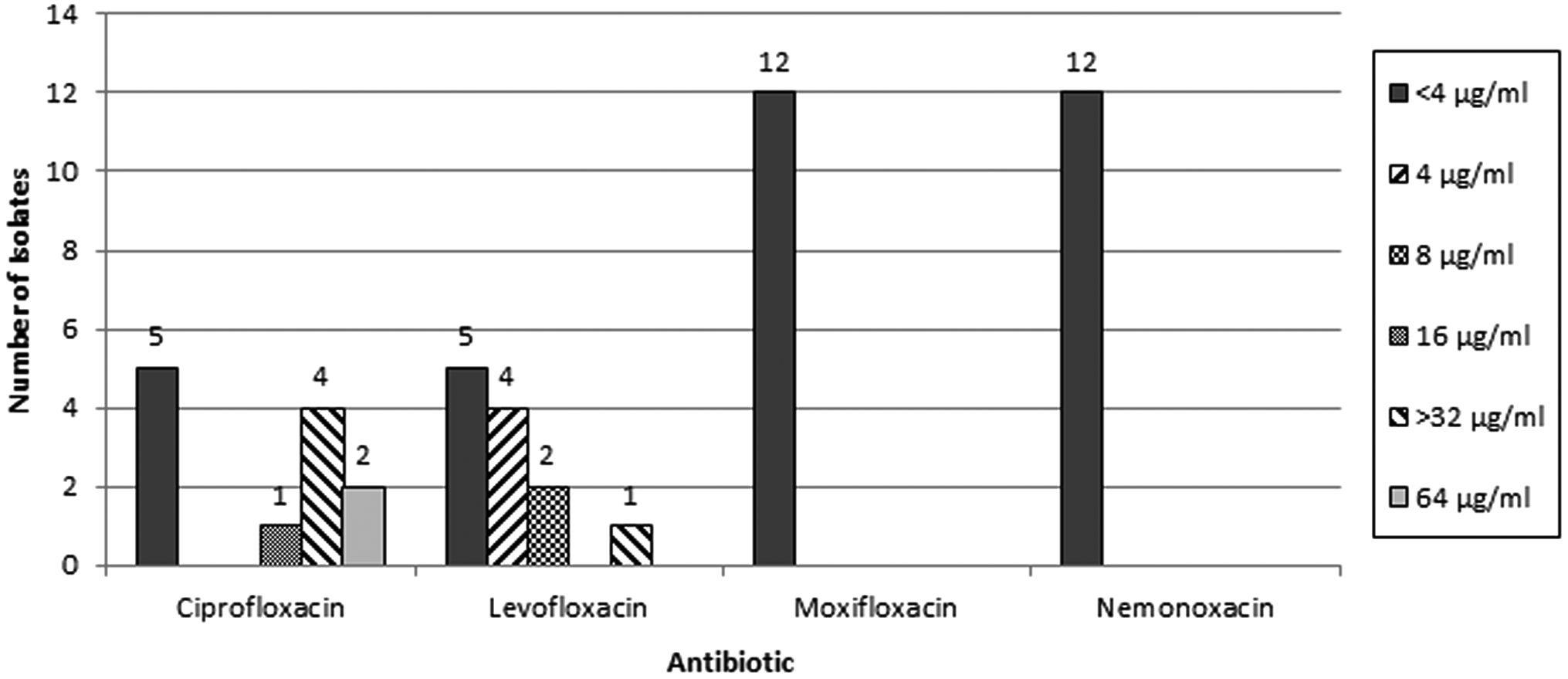
MIC values of selected antibiotics against the induced mutants isolated in this study. MIC, minimum inhibitory concentration.
Discussion
Fluoroquinolone resistance frequently emerges by changes of the target enzymes, such as DNA gyrase and topoisomerase IV, or by lowering intracellular accumulation of fluoroquinolones. 12 Fluoroquinolone resistance in E. faecalis and E. faecium is related with mutations in the QRDRs 13 of the gyrA or parC genes.14,15 High-level fluoroquinolone-resistant enterococci were found to have mutations in GyrA, GyrB, or ParC, while no amino acid substitution has been found in either of the three mentioned genes in the low-level resistant strains.15,16
Although some mutations in other QRDRs, such as parE, have been noted to induce the fluoroquinolone resistance of several bacteria, such as Escherichia coli, S. aureus, and S. pneumoniae, 12 there was no previous study investigating the association between these mutations and fluoroquinolone resistance in enterococci. In our study, a previously unreported amino acid substitution, S378-G in parE, was found, and it correlated with a significantly higher ciprofloxacin MIC for the E. faecium strain. This type of amino acid substitution was not found in the isolates exposed to levofloxacin, moxifloxacin, or nemonoxacin.
For the isolates of S. aureus in our study, we found that S80F or S80Y in parC and S84L or S84I in gyrA were associated with higher MICs for ciprofloxacin and levofloxacin, and the results were compatible with some previous studies.17,18 Otherwise, E84-K and A116-E in parC caused a high levofloxacin MIC for S. aureus, while A116-E and R159-K mutations in parC only mildly elevated MICs of moxifloxacin for S. aureus, as found previously. 19 Similar to a previous study in Taiwan, nemonoxacin retained good potency for many resistant Gram-positive pathogens, such as MRSA and S. pneumoniae. 20 Our mechanistic study further corroborated these observations.
In S. pneumoniae, either topoisomerase IV or DNA gyrase can be the primary target for quinolone resistance. A recent study revealed mutations were selected by nemonoxacin in clinical isolates of S. pneumoniae, including GyrB (S494-T), ParE (P454-S), and GyrA (S82-P) (20). However, our results failed to show the selection of QRDR mutations in different isolates against S. pneumoniae with nemonoxacin. Two of the three S. pneumoniae isolates were identified to have S79Y substitution in parC, and this resulted in a 64-fold elevation of MIC for ciprofloxacin.
S79Y or some other amino substitutions had been documented to induce such fluoroquinolone resistance. 21 Compared to ciprofloxacin, there were no other identifiable QRDR mutations for levofloxacin, moxifloxacin, or nemonoxacin. Mutations selected with nemonoxacin were often limited to gyrA, gyrB, and parE, sparing parC. The parC is known for its pivotal role on quinolone resistance among clinical isolates of S. pneumoniae. Our results are consistent with previous nonfluoroquinolones (NFQs) whose potencies were unaffected if there were no parC QRDR mutations in clinical isolates of S. pneumoniae. 22
Enterococci have caused more and more nosocomial infections over the past two decades. In a previous study, high-level ciprofloxacin resistance emerged from the point mutations in gyrA and parC, which were confined to a universal hospital-adapted clonal E. faecium strain. 23 Our study also demonstrated the amino acid substitutions with S378-G in parE, and this resulted in a significantly higher MIC of E. faecium for ciprofloxacin. However, we could not find QRDR mutations induced by nemonoxacin against Enterococcus species.
Our study had certain limitations that deserve to be mentioned. First, only limited numbers of bacterial strains and species were tested. However, our experiment exposed ∼1 × 109 to 1 × 1010 colony-forming units of bacteria, which should be an ample population for mutation frequencies that has been observed in selection of quinolone resistance. Second, the role of efflux pumps and multidrug transporters was not assessed in this study. 24 Therefore, the overall mechanisms for nemonoxacin to retain its antimicrobial potency could not be elucidated by this study. In one strain of our E. faecium (EM1), the MIC for levofloxacin and ciprofloxacin increased after drug exposure, but we found no QRDR mutations in the strain. Neither could other novel mechanisms of resistance development be excluded.
To make a conclusion, our results suggest that nemonoxacin is less vulnerable to inducible quinolone resistance in commonly relevant Gram-positive bacteria, including resistant strains, such as ampicillin-resistant enterococci, MRSA, and quinolone-resistant S. pneumoniae. It may thus be potentially reserved for the treatment of such resistant Gram-positive pathogens.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
The TaiGen Biotechnology Co. Ltd. provides financial support for laboratory labor and materials. The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.