
Abstract
Staphylococcus epidermidis is a commensal inhabitant of human skin and mucosa, and a common nosocomial pathogen in immunocompromised patients. S. epidermidis strains were isolated from places with precisely defined and controlled air quality and regular cleaning and disinfection regimes—cleanrooms. These strains were adapted to increasing concentrations of the quaternary ammonium disinfectant didecyldimethylammonium chloride (DDAC). Compared to nonadapted strains, these strains became adapted to up to 180-fold higher concentrations of DDAC, as seen by their increased minimal inhibitory concentrations. Examination of the stability of adaptation showed that three strains became permanently adapted to DDAC and named as resistant strains, and four strains were temporarily adapted to DDAC and named as strains with higher tolerance to DDAC. Some adapted strains showed cross-resistance to benzalkonium chloride and/or antibiotics. The adaptation response mechanisms of these DDAC-adapted strains were also investigated. The majority of adapted strains showed modifications to cell size and fatty acid composition. Some of the adapted strains showed changes in biofilm formation and overexpression of efflux pumps. Three adapted strains also showed altered growth rates. In this first report of adaptation of S. epidermidis strains to DDAC, the fatty acid profiling showed that the majority of strains had reduced ratio of saturated to unsaturated fatty acids and decreased content of straight-chain fatty acids, at the expense of the anteiso-branched fatty acids. We can conclude that S. epidermidis strains can adapt or become resistant to DDAC. We have revealed several adaptive response mechanisms that can be targeted for control and inhibition of S. epidermidis in cleanrooms and other clean processing environments.
Introduction
The aim of any disinfection protocol is to control contamination, which is an important part of the quality aspects of cleanrooms. Cleanrooms are rooms with defined and controlled air quality (e.g., generally using high-efficiency particulate air filtration systems), number of particles in the air, temperature, and humidity. They regularly undergo cleaning and disinfection to assure minimal levels of microbial contamination. Cleanrooms are designed to ensure the quality and safety of many production areas, where microbial and particle contamination are critical factors, such as for surgical units, manufacturing of pharmaceuticals, food production, and biosafety laboratories.1–3
Quaternary ammonium compounds (QACs) are cationic substances that are often used as disinfectants, surfactants, dyes, and neuromuscular blocking agents. 4 Didecyldimethylammonium chloride (DDAC) is a QAC that is often used as a disinfectant, as it can increase cell membrane porosity, and therefore promote leakage of cellular contents. 5 Indeed, DDAC is currently one of the most frequently used QACs in cleanroom environments. Regular disinfection also leaves residues of QACs that help to maintain the cleanroom environment. However, through being exposed to and surviving such residual subinhibitory concentrations in their environment, bacteria can develop adaptive response mechanisms. 6
Under stress conditions, staphylococci can rapidly change their metabolic pathways to enhance their fitness to promote survival and persistence in the environment. 7 Staphylococcus epidermidis is a coagulase-negative staphylococcus, and it is the predominant bacterial species on the human skin and mucosa, from where it can be transferred into cleanrooms and other clean processing environments.8,9 S. epidermidis is an important nosocomial pathogen, and can adapt to different environmental conditions, not only in terms of physical stress but also in terms of presence of antimicrobials, such as disinfectants, preservatives, and antibiotics.4,10
As reported previously, S. epidermidis is currently more tolerant to the disinfectant triclosan than isolates from the 1960s were, 11 which indicates that they have become adapted and can survive in the presence of higher concentrations of this disinfectant. Adaptation is a process, which allows cells to tolerate higher concentrations of antimicrobial substances. However, this is usually a reversible process and higher tolerance to antimicrobial substance can be lost after the removal of the antimicrobial. In contrast, resistance is a permanent change and genetically transferred, which enables the cells to grow under higher concentrations of an antimicrobial substance as their counterpart nonresistant cells. 4
Adaptation can be modulated through several cell properties, such as modifications of metabolic processes that can result in changed growth rates, fatty acid content, biofilm formation, and overexpression of efflux systems, or modifications of cell membranes.7,12,13 Adaptations to QACs can also lead to cross-resistance to other antimicrobials.14–16 However, there have been limited studies related to cleanroom isolates of S. epidermidis, with most of them related to S. epidermidis strains from clinical environments. Furthermore, alterations to biofilm formation represent an adaptation response mechanism. 5
S. epidermidis has the potential to form biofilms, where the cells can undergo long-term survival, resist antimicrobials, and overexpress certain genes. 17 Efflux pumps and their overexpression are usually the most widespread response mechanism in bacterial resistance. 18 Nearly 30 different drug efflux pumps have been identified in S. epidermidis, and some of these might export QACs out of the cells. 19 Changes in the fatty acid profile of cell membranes can help to maintain cell functionality in terms of cell shape and turgor pressure, and can have an influence on cell division, energy production, and regulation of cell permeability.7,20 Formation of branched fatty acids can change the proportion of iso-branched and anteiso-branched fatty acids, and changes in the saturated to unsaturated fatty acid ratio might occur after cells are exposed to antimicrobials or other environmental stress.21–24 Adaptation can also result in changes to the morphological properties of cells.6,10 As a consequence of such adaptation response mechanisms, the metabolic pathways in the cells can be modified to maximize the fitness of a population and to ensure the survival and persistence of the bacteria.
The aim of this study was first to investigate S. epidermidis strains isolated from cleanrooms in terms of their potential adaption to increasing levels of the QAC disinfectant DDAC. Furthermore, the resulting DDAC-adapted strains were characterized in terms of their cross-resistance to benzalkonium chloride and several antibiotics (i.e., ampicillin, erythromycin, gentamicin, tetracycline, chloramphenicol, and ciprofloxacin). Moreover, the adaptation response mechanisms that are crucial for S. epidermidis adaptation to DDAC were investigated, which included modulation of cell growth rate, biofilm formation, efflux pump activity, fatty acid profile, and cell size.
Materials and Methods
S. epidermidis strains
Six S. epidermidis strains were isolated from cleanrooms 9 and used along with S. epidermidis #60 (ATCC 51625). The strains were stored in 30% glycerol (Oleon, Ertvelde, Belgium) at −75°C. For experimental use, the suspensions (100 μL) were transferred into 4 mL Tryptic Soya Broth (TSB; Oxoid, Hampshire, United Kingdom) and incubated for 24 h at 37°C. The standardized inocula were prepared by adding 100 μL of these overnight cultures to 10 mL TSB without and with DDAC (Merck, Darmstadt, Germany). Optical density measurements at 600 nm (OD600) were used to estimate the numbers of S. epidermidis to 105 CFU/mL (colony forming unit [CFU]), although the precise numbers were determined using the plate count method.
Adaptation of S. epidermidis strains to DDAC
Adaptation of S. epidermidis strains to DDAC was performed as previously described, 25 with some modifications. The minimal inhibitory concentrations (MICs) for each strain were determined before their adaptation to DDAC, using the broth microdilution method in 96-well microtiter plates (Eppendorf, Hamburg, Germany). 26 For the DDAC adaptation, the S. epidermidis strains were exposed to a starting DDAC concentration of 0.25 × MIC in 10 mL TSB, with incubation for 24 h at 37°C. If the media were not turbid after 24 h, the incubations were prolonged up to 7 days. Each strain was inoculated in parallel. The TSB turbidity, which indicated the growth of the strains, was checked after 24 h and up to 7 days of incubation. For the cultures where growth was detected, 100 μL was then transferred to the next higher concentration of DDAC. These DDAC concentrations in TSB were increased by a factor of two for each transfer, passing from 0.25 × MIC to 0.5 × MIC, to 1 × MIC, to 2 × MIC, and up to 180 × MIC. If no growth was seen after 7 days of incubation, the samples were centrifuged (11,714 g, 2 min), and the pellets resuspended in 100 μL TSB and spread on Tryptic Soya Agar (TSA; Oxoid) that included the corresponding concentration of DDAC, to determine whether there were any adapted cells in the sample. These TSA samples were incubated for 24 h or up to 7 days at 37°C, and the growth of colonies was monitored. One colony from each plate was further transferred to TSB with the next higher concentration of DDAC. If there was no growth on the TSA plate after 7 days, the previous concentration of DDAC was defined as the maximum concentration to which the strain had become adapted.
During the adaptation to DDAC, 1 mL aliquots of the cultures from each step of the DDAC adaptation were stored at −75°C in 30% glycerol suspension for further analysis. In addition, for each strain, a control sample was prepared that contained the S. epidermidis strain in TSB with no DDAC. Growth of these control incubations was monitored using the same protocol for the strains undergoing adaptation.
To determine the stability of the adaptation to DDAC, the strains were then passaged 10 times (one passage every 24 h) in TSB with no DDAC.5,13 The MIC to DDAC for each strain was determined every day after each passage. After these 10 postadaptation passages, if the MIC remained as it had been initially for the DDAC-adapted strain, the strain was defined as resistant to DDAC. If the MICs of the strains following the 10 postadaptation passages showed full or partial reversal toward the MIC before DDAC adaptation, the strain was defined as DDAC adapted, rather than DDAC resistant.
Cross-resistance of adapted S. epidermidis strains
The MICs of these nonadapted and DDAC-adapted S. epidermidis strains were determined for benzalkonium chloride, ampicillin, erythromycin (all from Sigma-Aldrich, St. Louis, Missouri), gentamicin sulfate (Glentham, Life Science, Corsham, United Kingdom), ciprofloxacin, tetracycline (both from Fluka, Chemie GmbH, Buchs, Switzerland), and chloramphenicol (Merck). The stock solutions of the antibiotics were at 64 μg/mL, and the stock solution for benzalkonium chloride was 500 μg/mL. The antibiotic breakpoints were the concentrations defined by the Clinical and Laboratory Standards Institute (CLSI) and the European Committee on Antimicrobial Susceptibility Testing (EUCAST) breakpoints.26,27 Cross-resistance was defined as increased MICs to benzalkonium chloride or to the antibiotics after DDAC adaptation.
Growth of the S. epidermidis strains
The nonadapted S. epidermidis strains and the DDAC-adapted S. epidermidis strains were prepared in TSB alone or with the appropriate DDAC concentration, respectively, as indicated in Table 1, to the standardized OD600 of 0.1. Fifty microliter samples of each strain were transferred into microtiter plates, and following replacement of the plastic covers, they were incubated for 20 h at 37°C in a microtiter plate reader (Varioscan Lux, ThermoScientific, Vantaa, Finland). The absorbance at 600 nm (A600) was measured every 20 min for 20 h. The growth rate for each strain was calculated as the slope of Eq. (1) (using MS Office Excel):
Staphylococcus epidermidis Strains and Their Adaptation to Didecyldimethylammonium Chloride, and Their Cross-Resistance to Benzalkonium Chloride and Antibiotics
Strains showing adaptation to DDAC that were stable for 10 postadaptation passages in TSB, defined in this study as strains resistant to DDAC.
Ratio of MIC of adapted strain/MIC of nonadapted strain; MIC increase <1 indicates lower MIC after adaptation.
Following 10 postadaptation passages in TSB.
Upper limit for QAC, above which the strain is resistant. 32
Antibiotic breakpoint according to the Clinical and Laboratory Standards Institute. 33
Antibiotic breakpoint according to the European Committee on Antimicrobial Susceptibility Testing. 27
/, no fold difference determined.
AMP, ampicillin; BC, benzalkonium chloride; CHL, chloramphenicol; CIP, ciprofloxacin; DDAC, didecyldimethylammonium chloride; ERY, erythromycin; GEN, gentamicin; MIC, minimal inhibitory concentration; ND, not done; QAC, quaternary ammonium compound; TET, tetracycline; TSB, Tryptic Soya Broth.
where μ is the growth rate, and A1 and A2 are A600 at incubation times of t1 and t2.
Biofilm formation
Protocol and cutoff values to determine biofilm formation were determined as described before, with some modifications. 28 Briefly, overnight cultures were freshly diluted in TSB, and 200 μL was transferred into 96-well microtiter plates and incubated for 24 h at 37°C. After this incubation, the unattached cells were washed and the remaining attached cells were fixed (10 min, 60°C, and dyed with 1% crystal violet [Merck]). Then, the crystal violet solution was removed, and 96% ethanol (Hungrana, Ipartelep, Hungary) was added to each well. The absorbances were then measured at 584 nm (A584) using a microtiter plate reader. 9 The results were determined according to the previously described cutoff values. A cutoff absorbance (Ac) was fixed as three standard deviations above the mean absorbance of a pure TSB medium and the final absorbance of the tested strain was calculated as the mean absorbance for the strain minus Ac, and expressed as ΔA. Each strain was classified as not biofilm former (ΔA < Ac), weak biofilm former (Ac < ΔA < 2 × Ac), medium biofilm former (2 × Ac < ΔA < 4 × Ac), and strong biofilm former (ΔA > 4 × Ac).28,29
Efflux pump activity
The involvement of efflux in adaptation of S. epidermidis to DDAC was evaluated using the acridine orange (Thermo Fisher) uptake assay, 30 with some modifications. Briefly, the S. epidermidis strains were grown in TSB for 20 h at 37°C. The nonadapted and DDAC-adapted strains were swabbed in four parallel lines from the center to the edge of TSA plates that contained different concentrations of acridine orange (i.e., 5, 10, 20 μg/mL). After an incubation for 16 h at 37°C, the fluorescence was observed under UV at 366 nm. Increased efflux pump activity was seen as fluorescence at higher concentrations of acridine orange, and the fluorescence of the DDAC-adapted strain was compared to that of the nonadapted strains.
Scanning electron microscopy
The cell sizes of S. epidermidis cells were determined using scanning electron microscopy and measuring the cell diameters. The nonadapted and DDAC-adapted S. epidermidis strains were grown on TSA and TSA with the relevant DDAC concentration, respectively, for 18–24 h at 37°C. One colony for each strain was smeared onto a metal holder covered in carbon foil. These samples were air dried, sputtered with carbon dust (ACE600; Leica, Vienna, Austria), and analyzed using scanning electron microscopy (Carl Zeiss Ultra Plus+ EDS Oxford X-Max; Carl Zeiss AG, Oberkochen, Germany) with magnification of 50,000 times. The cell diameters were measured using ImageJ 1.51 of the National Institutes of Health (Bethesda, MD; https://imagej.nih.gov/ij/), three times on at least three cells in the scanning electron microscopy field. The cell diameters of the nonadapted and DDAC-adapted strains were compared.
Fatty acid composition of S. epidermidis strains
The fatty acid compositions of the nonadapted and DDAC-adapted strains were determined by gas chromatography using in situ transesterification, as previously described. 31 The fatty acid methyl esters (FAMEs) were determined by capillary gas chromatography (GC 6890; Agilent Technologies, Santa Clara, CA) with a flame ionization detector and capillary column (HP-5MS; 25 m × 0.2 mm × 0.33 μm; Agilent Technologies). Separation and detection were performed under the following temperature conditions: 150°C held for 4 min, raised at a rate of 4°C/min to 250°C, and then held for 5 min. The total analysis time was 34 min. The injector and detector temperatures were 240°C and 280°C, respectively. The carrier gas was helium, at a flow rate of 0.8 mL/min. The injected volume was 1 μL, and the injector split ratio was 1:30. Nitrogen was used as the makeup gas, at a flow rate of 45 mL/min; the detector gases were hydrogen and synthetic air (21% O2), at a rate of 40 and 450 mL/min, respectively.
The FAMEs were determined through their retention times in comparison to the relevant standard mixtures used: Bacterial Acid Methyl Ester Mix (47080-U); 37 Components FAME Mix (Supelco, Bellefonte); and Nu-Chek standards GLC-68D, GLC-85, and GLC-411g (Nu-Chek, Minnesota). The GLC-68D and GLC-85 standard mixtures were used to determine the response factors for each fatty acid. The weight of each fatty acid in each sample was determined using the response factor and the transformation factor of the fatty acid content from the FAME content. The samples were analyzed in duplicate. The FAMEs were expressed as weight percentages of the total fatty acid content.
Statistics
The data are expressed as mean log (CFU/mL) ± standard deviation (SD) within all of the sample groups. The mean and SD were calculated using analysis of variance. In addition, the IBM SPSS Statistics package (IBM Corp., USA) was used for comparisons of the MIC changes, using Mann–Whitney U-tests, and comparisons of the biofilm assays, using t-tests and analysis of variance. p-Values less than 0.05 was taken to define significance.
Results
Adaptation of the S. epidermidis strains to DDAC
In the stepwise adaptation of S. epidermidis strains to DDAC, all of the seven tested strains showed increased MICs of DDAC. After adaptation to DDAC, MICs increased from the lowest 2-fold to the highest of 180-fold change (Table 1). Five of the DDAC-adapted S. epidermidis strains (i.e., S. epidermidis #4, #11, #18, #46, #60) exceeded the previously published upper expected limit for commercial QACs of 5.0 μg/mL. 32 Interestingly, the lower MICs in the strains before adaptation resulted in greater fold changes after adaptation to DDAC. This was seen for S. epidermidis #4, #18, #46, and #49, which adapted with a minimal 15-fold change and a maximal 180-fold change. In contrast, the lower fold change was seen in strains that have higher MICs of DDAC as nonadapted strain such as for S. epidermidis #1 and #11 (i.e., twofold and fourfold).
After the DDAC-adapted strains had undergone 10 postadaptation passages in pure TSB, the MICs for three of these strains remained constant, as S. epidermidis #1, #18, and #49, which indicated that they had become resistant to DDAC, through irreversible cell changes (Table 1). The MIC of DDAC-adapted S. epidermidis #11 decreased to that for the nonadapted strain after these 10 postadaptation passages. The MICs of DDAC-adapted S. epidermidis #46, #60, and #4 also decreased during these postadaptation passages, which indicated that they had undergone temporary changes to their cellular adaptation mechanisms.
Cross-resistance of the DDAC-adapted S. epidermidis strains to benzalkonium chloride and antibiotics
The DDAC-adapted S. epidermidis strains were screened for cross-resistance to benzalkonium chloride and six antibiotics according to the reference concentrations (Table 1).27,32,33 DDAC-adapted S. epidermidis #60 was cross-resistant to benzalkonium chloride, and DDAC-adapted S. epidermidis #4 and #49 showed fourfold increased MICs for benzalkonium chloride.
According to the antibiotic breakpoints defined by the CLSI 33 and the EUCAST, 27 some of the nonadapted S. epidermidis strains used in this study were already resistant to the antibiotics tested. The majority of these strains were resistant to ampicillin (n = 6/7) and gentamicin (n = 5/7), and some strains also to chloramphenicol (n = 2/7), erythromycin (n = 1/7), and tetracycline (n = 1/7) (Table 1). Thus, the cross-resistance of the DDAC-adapted S. epidermidis strains to these different antibiotics was evaluated only on the strains that were not resistant to these antibiotics in terms of their nonadapted strains (according to the CLSI and EUCAST breakpoints).27,33 Nonadapted S. epidermidis #11 was the only strain that was sensitive to ampicillin, for which its MIC increased by more than twofold for DDAC-adapted S. epidermidis #11. Cross-resistance to gentamicin, erythromycin, ciprofloxacin, and chloramphenicol was shown for DDAC-adapted S. epidermidis #60 (i.e., ATCC 51625), which became highly resistant to all of these antibiotics, with the exception of tetracycline. Cross-resistance to ciprofloxacin and tetracycline was also seen for DDAC-adapted S. epidermidis #49, and to chloramphenicol and tetracycline for DDAC-adapted S. epidermidis #1. The other S. epidermidis strains that were tested did not show any cross-resistance to these antibiotics (Table 1).
Adaptation response mechanisms of DDAC-adapted S. epidermidis
The DDAC-adapted S. epidermidis strains were characterized by evaluation of their (1) growth rate; (2) biofilm formation; (3) efflux pump activity; (4) cell size; and (5) fatty acid composition.
Growth rate
The growth rates for these S. epidermidis strains were calculated from the curves of the exponential growth phase. There were no significant changes in the growth rates of DDAC-adapted S. epidermidis #4, #18, #46, and #49 (Table 2). However, DDAC-adapted S. epidermidis #11 and #60 increased their growth rates in comparison to their nonadapted strains. In addition, DDAC-adapted S. epidermidis #1 showed a lower growth rate in comparison to its nonadapted strain.
Characteristics of the Didecyldimethylammonium Chloride-Adapted Staphylococcus epidermidis Strains
Degree of fluorescence: −, no fluorescence; +, slight fluorescence; ++, intense fluorescence; +++, very intense fluorescence.
Might be involved in adaptation response mechanism.
Significantly different versus respective nonadapted strain (p < 0.05).
SD, standard deviation.
Biofilm formation
Biofilm formation was determined by biomass determination using the crystal violet assay. DDAC-adapted S. epidermidis #1, #11, and #49 were stronger biofilm formers than their nonadapted counterparts, while DDAC-adapted S. epidermidis #4 and #60 were weaker biofilm formers than their nonadapted counterparts (Table 2). It appears that stronger biofilm formation is a DDAC adaptation response mechanism for strains that have initially low biofilm formation, as nonadapted S. epidermidis #11 and #49 were weak biofilm formers and became stronger biofilm formers with DDAC adaptation. Decreased biofilm formation upon DDAC adaptation was also seen in some strains that were initially (i.e., when nonadapted) strong biofilm producers. No significant changes in biofilm formation after DDAC adaptation were seen for DDAC-adapted S. epidermidis #18 and #46.
Efflux pump activity
Efflux pump activity was determined by the intensity of fluorescence after uptake of different concentrations of acridine orange. Five of the DDAC-adapted S. epidermidis strains showed increased efflux pump activity compared to their nonadapted strains: S. epidermidis #1, #4, #11, #18, and #46. For DDAC-adapted S. epidermidis #49 and #60, no changes in efflux pump activities were seen (Table 2).
Cell size
The nonadapted and DDAC-adapted S. epidermidis strains were screened for their cell sizes using scanning electron microscopy. Three to five micrographs were taken for each strain and the cell diameters were measured three times on at least three cells within each micrograph frame. Table 2 indicates the calculated mean cell diameters for each nonadapted and DDAC-adapted S. epidermidis strain. In this study, the cells of the nonadapted and DDAC-adapted S. epidermidis strains showed different sizes. The mean cell diameter for the nonadapted S. epidermidis strains was 841 ± 49 μm. Reductions in cell diameters in comparison to the nonadapted strains was seen for six of the DDAC-adapted strains: S. epidermidis #1, #4, #11, #18, #46, and #60 (Table 2). For these DDAC-adapted strains, the mean cell diameter was 735.5 ± 44 μm. For DDAC-adapted S. epidermidis #49, there was no change in cell size. Figure 1 shows images of the S. epidermidis #4, #11, and #49 cells, without and with DDAC adaptation.

Representative scanning electron microscopy images of the nonadapted and DDAC-adapted Staphylococcus epidermidis strains. DDAC, didecyldimethylammonium chloride.
Fatty acid composition
Fatty acid analysis of the nonadapted and DDAC-adapted strains was carried out using gas spectrometry following modified in situ transesterification. The highest content was for anteiso-C15:0 branched fatty acids, followed by anteiso-C17:0, C18:0, C20:0, and iso-C17:0. All of these investigated S. epidermidis strains changed their fatty acid profiles upon DDAC adaptation (Table 3). The changes in the contents of straight-chained fatty acids and branched fatty acids are shown in Fig. 2. Six of these DDAC-adapted strains showed decreased content of straight-chained fatty acids: S. epidermidis #1, #4, #11, #18, #49, and #60. The iso-branched fatty acids were reduced in six of these DDAC-adapted strains: S. epidermidis #4, #11, #18, #46, #49, and #60. Also, the content of the anteiso-branched fatty acids was increased in five of the DDAC-adapted strains: S. epidermidis #1, #4, #11, #18, and #60. Finally, the ratio of saturated to unsaturated fatty acids decreased in five of these DDAC-adapted strains: S. epidermidis #11, #18, #46, #49, and #60, as shown in Fig. 3.
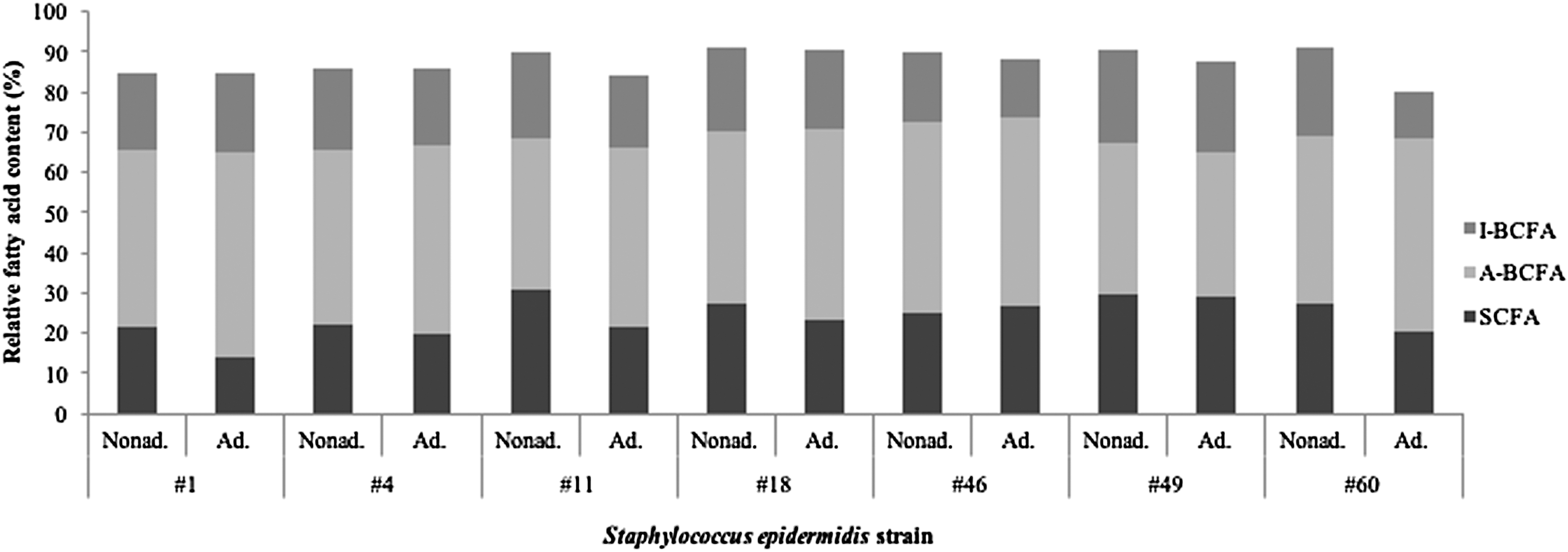
Content of I-BCFA and A-BCFA and SCFA in the nonadapted and DDAC-adapted Staphylococcus epidermidis strains. A-BCFA, anteiso-branched fatty acids; Ad., adapted; I-BCFA, iso-branched branched fatty acids; Nonad., nonadapted; SCFA, straight-chain fatty acids.
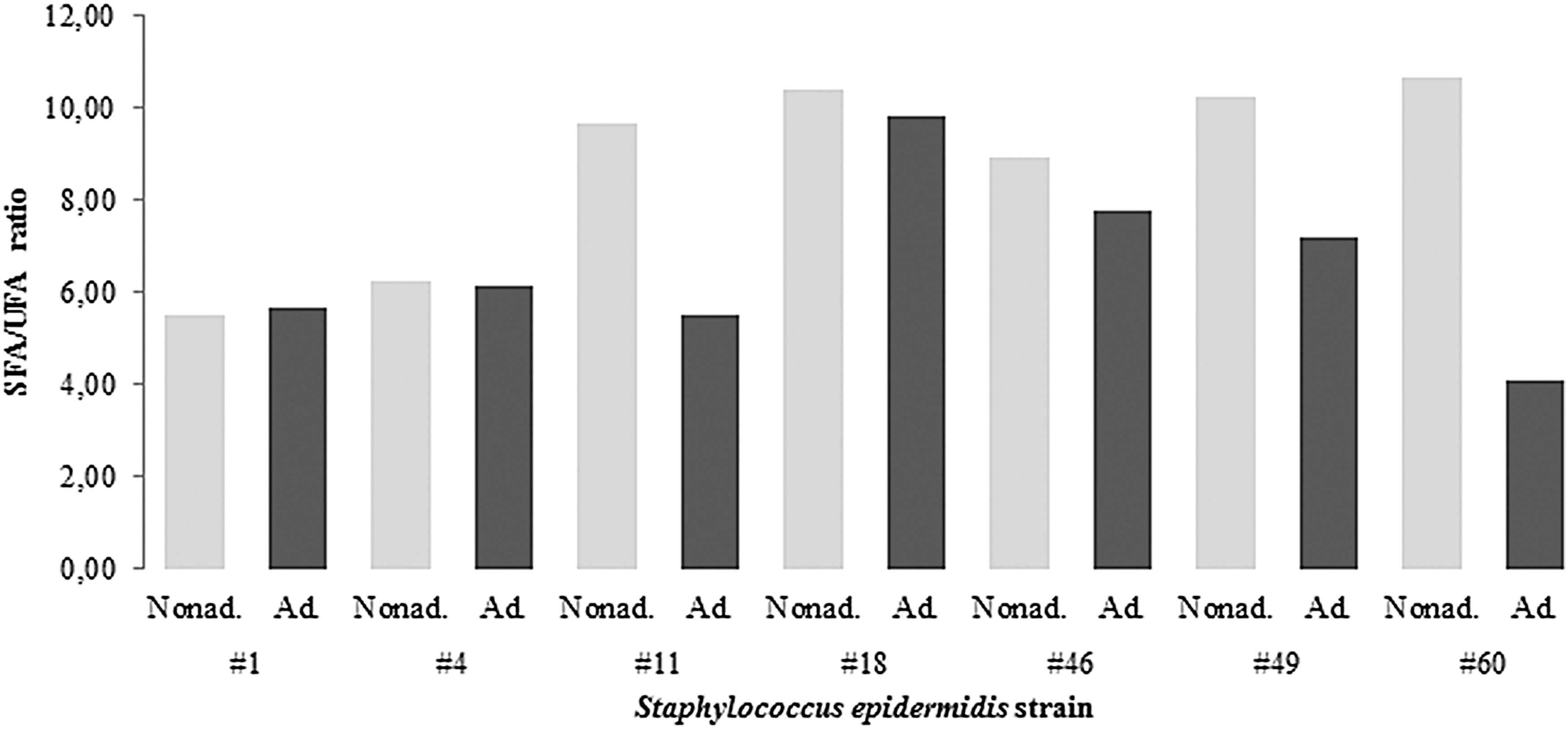
Ratio of SFA to UFA in the nonadapted and DDAC-adapted Staphylococcus epidermidis strains. SFA, saturated fatty acids; UFA, unsaturated fatty acids.
Fatty Acid Methyl Ester Levels in the Nonadapted and Didecyldimethylammonium Chloride-Adapted Staphylococcus epidermidis Strains
Nonidentified fatty acid.
Discussion
S. epidermidis is the most common bacterial contaminant for cleanrooms and clean processing environments where human operators are involved. Their primary source are the cleanroom operators themselves, from their skin and mucosa. 34 Thus staphylococci are frequently exposed to disinfectants such as DDAC, which is one of the most frequently used disinfectants. 4 Adaptation and/or resistance to DDAC of S. epidermidis cleanroom isolates has not yet been widely examined, although S. epidermidis frequently shows the potential to adapt to, or even become resistant to, different antimicrobials, such as antibiotics and disinfectants. 11 Based on a review of the literature available, primary S. epidermidis isolates from clinical environments have been studied.5,10,11 Therefore, we investigated S. epidermidis cleanroom isolates 9 and their adaptation to DDAC, and we looked for their adaptation response mechanisms.
S. epidermidis strains of human origin previously showed more than fourfold increased MICs for the QAC cetrimide. 5 We have shown in this study that S. epidermidis cleanroom isolates can also adapt to DDAC. Furthermore, three of these strains even became DDAC resistant, where this resistance is a permanent and irreversible change after adaptation and the later removal of the antimicrobial. 4
The QAC disinfectants DDAC and benzalkonium chloride both target the cell membrane. According to previous published upper expected limits for commercial QACs (5.0 μg/mL), 32 in this study, only one DDAC-adapted strain became cross-resistant to benzalkonium chloride, although some of the other DDAC-adapted S. epidermidis strains also had higher MICs for benzalkonium chloride. The data from this study also indicate that resistance to one QAC does not ultimately indicate resistance to other QACs, although resistance to one QAC can lead to higher tolerance or cross-resistance to other QACs.
Resistance of staphylococci to antibiotics such as penicillins and gentamicin has been frequently reported.8,11,32 This was also confirmed in these nonadapted S. epidermidis strains tested. Adaptation to DDAC resulted in cross-resistance to different antibiotics for four of these S. epidermidis strains, which indicated that cross-resistance among QACs and antibiotics can manifest in S. epidermidis cleanroom isolates.
As previously reported,5,10,13 different adaptation response mechanisms can be detected in such adapted S. epidermidis strains. We characterized these DDAC-adapted S. epidermidis strains in terms of changes in growth rate, biofilm formation, efflux pump activity, cell size, and fatty acid composition, as possible adaptation response mechanisms.
Compared to the growth rates of the nonadapted strains, changes were seen for three of these DDAC-adapted strains. Two strains increased their growth rates after DDAC adaptation, similar to what has been seen elsewhere. 13 Increased growth rates might be attributed to adaptation of ribosomes and increased protein synthesis rates. 35 One of these DDAC-adapted S. epidermidis strains decreased its growth rate following adaptation. As reported before, decreased growth rates and higher biofilm formation are correlated, 36 and this was seen in this study for DDAC-adapted S. epidermidis #1; this strain showed a decreased growth rate, but became a stronger biofilm former.
One of the adaptation response mechanisms in S. epidermidis strains is thus a change in growth rate, with the selection of isolates with metabolic modifications that result in altered growth rates and fitness.7,13,37
Adaptation to QACs can also enhance biofilm formation, as previously reported. 13 Survival of cells in biofilms under stress can be prolonged. This was possibly the case in the three adapted S. epidermidis strains in this study that showed increased biofilm formation, S. epidermidis #1, S. epidermidis #11, and S. epidermidis #49. In contrast, two S. epidermidis strains, S. epidermidis #4 and S. epidermidis #60, decreased biofilm formation. These data support previous findings, 5 where both increases and decreases in biofilm formation were seen among staphylococci after multiple exposure to antimicrobials. The mechanisms proposed relate to the selection of isolates with alterations in factors involved in biofilm formation, such as alterations in factors involved in cell adhesion and biofilm maturation, or selection of isolates with metabolic changes that result in growth rate and fitness, which can affect biofilm formation.5,7
Expression and/or overexpression of efflux pumps can enable cells to remove toxic compounds from the cells, and this is usually seen as one of the most important adaptation response mechanisms.38,39 QACs can induce expression of the QAC efflux pumps that are encoded by different qac genes, and in this study, overexpression of the efflux pumps was seen for five of the DDAC-adapted S. epidermidis strains, each of which carried at least one qac gene. S. epidermidis #60 was the only strain missing all of the qac genes, and as expected, this strain showed no changes in efflux pump activity. This accompanies previous findings, 38 where adaptation of qac-negative staphylococci to QAC was possible. Cells with overexpressed efflux pumps can remove toxic compounds from the cells more efficiently, and this was an important adaptation response mechanism in these S. epidermidis strains.
Exposure of S. epidermidis to antimicrobials might also alter cell morphology and size, 10 which might represent an adaptation response mechanism as a result of modifications to metabolic pathways for cell wall or membrane components. 40 According to measurements of the S. epidermidis cell diameters before and after DDAC exposure, reduction in cell size was the most common adaptation response mechanism for these S. epidermidis strains, as previously reported. 10 The adaptation response mechanism might also be coexistent with the S. epidermidis small-colony variants (SCVs) phenotype. SCV S. epidermidis colonies are characterized as bacterial subpopulations with atypical growth features, such as smaller colonies and loss of pigmentation.10,41 We did not observe such colonies for these S. epidermidis strains either before adaptation or after DDAC adaptation. The reductions in cell sizes seen are most likely due to the altered metabolism induced by DDAC adaptation, and not the persistence of SCVs.
Fatty-acid modifications promote maintenance of membrane integrity. For the first time, we have shown, in this study, differences in the relative proportions of different fatty acids after DDAC adaptation of these S. epidermidis strains. The presence of an antimicrobial can result in a shift in the proportion of anteiso-branched fatty acids and a reduction in the proportion of iso-branched fatty acids and straight-chain fatty acids, 15 and this corresponds to our findings in four strains. Furthermore, a decreased ratio of saturated to unsaturated fatty acids affects membrane fluidity, as we have shown for five of these S. epidermidis strains. Indeed, the modification of this ratio was one of the main adaptation response mechanisms in these S. epidermidis strains. Changing the fatty acid contents, the proportions of straight-chain to branched-chain fatty acids, and the saturated to unsaturated fatty acid ratio regulates the fluidity of cell membranes, for the maintenance of their functions. 7
Bad disinfection practices (i.e., missing cleaning prestep, incorrect disinfectant preparation, contaminated disinfection equipment, and overcontaminated surfaces in cleanrooms) can allow the survival of cleanroom S. epidermidis strains and their multiple exposure to low concentrations of DDAC. This can result in adaptation and resistance to DDAC, and even cross-resistance to other antimicrobials. Our study confirmed that S. epidermidis can adapt and even become resistant to DDAC. Adapted S. epidermidis strains can also become cross-resistant to other disinfectants and antibiotics. Allowing the spread of these strains to the most critical area for microbial contamination (i.e., surgical wounds, biological preparations, and food and drug products) is concerning. Therefore, correct disinfection measures together with a monitoring program should be provided in cleanrooms to prevent such adaptation, resistance, and cross-resistance phenomena, as stated before. 8
Several adaptation response mechanisms can be induced during this process of DDAC adaptation. The predominant changes revealed in this study were for cell size and fatty acid profile. Changes in growth rate and biofilm formation, and overexpression of efflux pumps, were also seen. However, in particular, and shown for the first time, the membranes of these S. epidermidis cleanroom isolates are an important target, with modifications to the fatty acid composition, in terms of the proportions of straight-chain to branched-chain fatty acids, and modifications to the saturated to unsaturated fatty acid ratio. The consequence of these changes will be modifications to the fluidity of the cell membranes. Knowledge of these mechanisms will now serve in the continued search for novel targets for successful inhibition and elimination of these contaminating microbiota from cleanrooms, and to indirectly monitor the effectiveness of disinfection programs.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was co-funded by the Slovenian Research Agency (Program P4-0116).