
Abstract
Aim:
The aim of this study was to evaluate the effect of subinhibitory concentrations of trans-anethole on antibacterial and antibiofilm properties of mupirocin against mupirocin-resistant Staphylococcus aureus strains.
Methods:
Following parameters were examined: isolates susceptibility to antibiotics, minimum inhibitory concentration (MIC) of trans-anethole, antibacterial activity of mupirocin/trans-anethole combination, detection of ileS2 gene, genotypic relativity of isolates using pulsed-field gel electrophoresis method, and the influence of mupirocin/trans-anethole combination on S. aureus biofilm formation.
Results:
Our study revealed that trans-anethole combined with mupirocin increased the growth inhibition zone diameter around the mupirocin disk, independently on S. aureus strains susceptibility to this antibiotic. Moreover, combination of subinhibitory (MIC50) concentration of mupirocin and trans-anethole significantly decreased biofilm biomass.
Conclusions:
trans-Anethole appeared efficient in increasing susceptibility to mupirocin and decreasing biofilm formation in S. aureus strains used in this study. Reduction of biofilm formation can potentially protect against S. aureus recolonization. Moreover, use of trans-anethole in combination with mupirocin can increase the mupirocin activity against methicillin-resistant and mupirocin-resistant S. aureus strains.
Introduction
Methicillin-resistant Staphylococcus aureus (MRSA) strain represents a major challenge for hospital treatment. These pathogens are outstandingly common nosocomial strains potentially easily transmitted in hospital environment. S. aureus similar to other staphylococci, reside on the human skin and in the nasal cavity. These staphylococci properties make proper prophylaxis the major tool in combating MRSA nosocomial infection. Prevention of MRSA infection is implemented by strict epidemiological surveillance for MRSA carriage among medical staff as well as certain groups of patients, for example, cardiosurgical or the hemodialyzed patients. According to the international standards, nasal vestibule smear is used to assess MRSA carriage. 1 Mupirocin administration to nasal cavity is recommended for MRSA positive patients in order to eradicate MRSA carriage and to interrupt the transmission pathway. The exact data on MRSA eradication effectiveness are limited; however, it is estimated that the incidence of recolonization level as 67% after 6 months. 2
Mupirocin is the topical antibiotic that exhibits high activity against staphylococci. It was initially isolated in 1971 from Pseudomonas fluorescens and was introduced into clinical practice in 1985 in the United Kingdom. 3 Available data have shown that 5 days intranasal application eliminates MRSA as well as methicillin-susceptible S. aureus (MSSA). 1 Nevertheless, the incidence of MRSA recolonization is probably underestimated and, in addition, due to the widespread use of mupirocin, increasing number of resistant strains appear.
There are two mupirocin-resistance phenotypes among S. aureus: low- and high-level resistance. First is the result of mutational change in the chromosomally encoded ileS gene, whereas the second mechanism is based on the acquisition of transferable plasmid containing the ileS2 gene encoding additional isoleucyl-tRNA synthetase.4,5
Mupirocin and chlorhexidine are widely used to decolonize MRSA carriers in medical facilities around the world, but literature data report their ineffectiveness. The combined low-level mupirocin and chlorhexidine resistance was confirmed in the case–control study on MRSA carriers who received decolonization therapy. 6 Liu et al. 7 showed that resistance to biocides was correlated with tolerance to chlorhexidine in 90.6% out of 53 of the tested high-level mupirocin-resistant MRSA isolates and was correlated with the presence of the plasmid-borne genes: qacA/B, smr as well. In addition, overexpression of chromosome-mediated genes norA and norB was observed. Therefore, it is necessary to investigate alternative methods of MRSA eradication in asymptomatic carriers.
Increasing interest is observed in plant-derived substances, potentially synergistically acting with conventional antibiotics, in which trans-anethole represents the major example. This is an organic compound formed in plants as a secondary product of terpene synthesis, one of the main components of anise, cinnamon, clove, and thyme essential oils. 8 trans-Anethole is widely used in the food and liquor industries, such as candies, chewing gum, and alcoholic beverages.9–11 According to the available data, trans-anethole has been shown presenting antibacterial activity.12,13
The aim of the study
The aim of this study was to evaluate the effect of subinhibitory concentrations of trans-anethole on antibacterial and antibiofilm activity of mupirocin against mupirocin-resistant S. aureus strains.
Materials and Methods
Bacterial strains and growth condition
The study included eight S. aureus strains belonging to Chair of Microbiology, Immunology and Laboratory Medicine, Pomeranian Medical University in Szczecin; isolated from nasal carriers (n = 4) and patients with superficial ambulatory-managed wound infections (n = 4). Specimen samples were cultured on Columbia agar with 5% sheep blood (bioMérieux, Poland) and incubated overnight at 37°C in aerobic atmosphere. Isolates were identified by colony morphology, Staph-Kit test (bioMérieux), biochemical test with use of GP-card on VITEK 2 Compact (bioMérieux), and amplification of nuc gene, which encodes the thermostable nuclease of S. aureus (A&A Biotechnology, Poland). S. aureus ATCC 29213 was used as a control strain in this study.
Susceptibility testing
The S. aureus susceptibility to mupirocin (200 μg) and cefoxitin (30 μg) was determined using disk diffusion method according to European Committee on Antimicrobial Susceptibility Testing. 14 Cefoxitin-resistant strains were considered as MRSA.
The prevalence of resistance to macrolides-lincosamides-streptogramin B (MLSB) antibiotics was determined by D-test using erythromycin (15 μg) and clindamycin (2 μg) disks antibiotics (Diag-Med, Poland). S. aureus strains were considered to have constitutive MLSB (cMLSB), inducible MLSB (iMLSB), and MSB resistance phenotypes when interpreted according to Clinical and Laboratory Standards Institute (CLSI) recommendations. 15
S. aureus susceptibility to trans-anethole
The agar dilution method was used according to the CLSI 16 with the following modification: a final concentration of 1% Tween 80 (v/v) (Sigma-Aldrich, Germany) was incorporated into MHA after autoclaving to enhance trans-anethole (99.0% purity was obtained from Sigma-Aldrich) solubility. trans-Anethole in various concentrations (1.0%, 2.0%, 4.0%, 6.0%, 8.0%, 10.0%, 12.0%, and 14.0%) (v/v) was transferred into MHA with 1% Tween 80 (v/v). Plates were dried at 37°C for 60 min before inoculation with suspension containing ∼104 CFU/mL of each S. aureus isolate, using a multichannel pipette. MHA with 1% Tween 80 (v/v) without trans-anethole was used as a positive growth control for S. aureus. Minimum inhibitory concentration (MIC) of each tested ingredient was determined after 24 hr as the lowest concentration of chemical inhibiting the visible growth of S. aureus on MHA plate (according to CLSI recommendation one or two colonies were ignored).
Antibacterial activity of mupirocin in combination with trans-anethole
Antibacterial activity of mupirocin in combination with trans-anethole was examined using agar dilution method and disk diffusion method. Following concentrations of trans-anethole were used to for the screening of antibacterial activity of mupirocin placed on the surface of MHA with 1% Tween 80 (v/v): 0.5%, 1%, 1.5%, and 2%. Plates were dried for 60 min at 37°C before inoculation with 108 CFU/mL of bacterial suspension using sterile cotton swab. The activity of antibiotic was studied with disk containing mupirocin (200 μg) on the surface of MHA plates (with and without chemicals—control). The plates were incubated for 18 hr at 37°C in aerobic conditions. After incubation period, inhibition zone around the disk was measured.
DNA extraction
The S. aureus strains were thawed and sown on Columbia agar with 5% sheep blood (bioMérieux), and incubated at 37°C for 18 hr. Next, one colony was harvested from the pure culture, suspended in 3 mL of tryptic soy broth (TSB; Sigma-Aldrich) and incubated for 18 hr at 37°C with shaking (100 rpm). Then, 1 mL of the culture was centrifuged for 5 min at 10,000 rpm, and the pellet was used for DNA extraction using GeneMatrix Bacterial & Yeast Genomic DNA Purification Kit (EURx, Poland) and lysostaphin (400 U/mL; DNA Gdansk, Poland) according to the manufacturer's procedure.
Polymerase chain reaction amplification
The following primers were used in this study: nuc1 (5′-GCG ATT GAT GGT GAT ACG GTT-3′) and nuc2 (5′-AGC CAA GCC TTG ACG AAC TAA AGC-3′) for nuc, ileS2-1 (5′-TAT ATT ATG CGA TGG AAG GTT GG-3′) and ileS2-2 (5′-AAT AAA ATC AGC TGG AAA GTG TTG-3′) for ileS2 (IBB-PAN, Poland). 17 The following reaction mixture was added to each sample: 1 μL of supernatant containing DNA, 150–400 nM each primer, 12.5 μL StartWarm HS-PCR Mix mixture consisting of 1 × reaction buffer, 2.5 mM MgCl2, 0.5 mM dNTPs, 0.1 U/μL Taq DNA polymerase, red dye, and loading buffer (A&A Biotechnology). The mixture was completed to 25 μL volume by distilled H2O. Amplification was performed using an Applied Biosystems Veriti 96 Well Thermal Cycler (Applied Biosystems). Initial denaturation at 95°C for 4 min was followed by 35 cycles of amplification (denaturation at 95°C for 15 sec, annealing at 53°C for 30 sec and extension at 72°C for 60 sec) and ending with final extension at 72°C for 7 min. All polymerase chain reaction (PCR) assay runs incorporated a reagent control (without template DNA) and with S. aureus ATCC 29213. After PCR amplification, 10 μL was removed and subjected to agarose gel electrophoresis (1.5% agarose, 1 × Tris/borate/EDTA—TBE, 100 V, 60 min) to estimate the sizes of the amplification products by comparison with a 100-bp molecular size standard ladder (DNA Gdansk). The gel was stained with 3 μL ethidium bromide (Sigma-Aldrich), and the amplicons were visualized using a gel image system GelDoc-It2 Imager (Upland, CA).
Molecular typing
All of the isolates were characterized by using pulsed-field gel electrophoresis (PFGE). Preparation of bacterial DNA was performed according to manufacturer of GenePath Group 6 Reagent Kit Instruction Manual (Bio-Rad, France) protocols using CHEF Bacterial Genomic DNA Plug Kits (Bio-Rad). Digestion of DNA was conducted using SmaI (ThermoScientific) according to procedure described by manufacturer. PFGE was performed with CHEF DR III apparatus (Bio-Rad) in TBE buffer; the conditions were 6 V/cm at 14°C, with an initial switching time of 2.2 sec and a final switching time of 54.2 sec. 1.2% agarose gel was run for 22 hr. SmaI digested DNA from S. aureus ATCC 29213 was included as a normalization standard on gel. Gels were stained with ethidium bromide and photographed in image system GelDoc-It2 Imager.
Restriction profiles were analyzed using the FPQuest program (Bio-Rad). The classification of individual restriction patterns for particular genetic profiles was made using the UPGMA (unweighted pair group method with arithmetic mean) method [SAB (average similarity value) value = 47.3%] and the Dice coefficient (2.0%). The results are presented in the form of a dendrogram.
Analysis of influence of mupirocin and trans-anethole on S. aureus biofilm
Biofilm was formed according to Skogman et al. 18 with minor modifications. First, TSB supplemented with 0.25% of D-+-glucose (TSBGlc; Sigma-Aldrich) was inoculated and incubated overnight. Inoculum was 20-fold diluted in TSBGlc and a total 100 μL was added to 96-well microtiter plate and incubated stationary for 24 hr at 37°C to form biofilm. After incubation, medium was removed, and formed biofilm was rinsed with PBS buffer. To rinsed biofilm TSBGlc supplemented with MIC50 of mupirocin or/and with 2% of trans-anethole was added. All samples were incubated for 12 hr. Afterward, medium was removed, and biofilm was rinsed with use of PBS. Biofilm biomass was quantified with use of crystal violet staining according to Skogman et al. 18 with minor modifications. Biofilm was washed and incubated for 20 min with the 200 μL of methanol (Sigma-Aldrich) for fixation. After removal of methanol, plates were air dried at room temperature. Wells were stained for 20 min with crystal violet 1% (w/v) (Sigma-Aldrich) and washed by sterile water. Afterward, ethanol:acetone mixture (in 8:2 ratio; Sigma-Aldrich) was added to each well. Absorbance indicating the biofilm biomass was measured at 595 nm wavelength (Perkin Elmer). All the experiments were conducted in triplicate.
Results
Antimicrobial susceptibility assay
The antimicrobial susceptibility resistance profile of S. aureus strains is summarized in Table 1. It was noted that all isolates were resistant to mupirocin, whereas 37.5% were resistant to cefoxitin (MRSA phenotype). iMLSB and MSB resistance phenotype were observed among 50.0% and 12.5% isolates, respectively. No cMLSB phenotype based on the D-test has been detected.
Characteristic of Staphylococcus aureus Strains Used in This Study
CC, clindamycin; E, erythromycin; FOX, cefoxitin; iMLSB, inducible macrolides-lincosamides-streptogramin B resistance phenotype; MIC, minimum inhibitory concentration; MRSA, methicillin-resistant Staphylococcus aureus; MSB, macrolides-streptogramin B resistance phenotype; MSSA, methicillin-susceptible S. aureus; MUP, mupirocin; R, resistant; S, susceptible.
Determination of MIC for trans-anethole
According to the agar dilution method, all strains were susceptible to trans-anethole (Table 1). The minimum concentrations of trans-anethole inhibiting growth of S. aureus strains ranged from 6.0% to 10.0%, but the major number of strains (n = 4) was inhibited in the concentration of 6.0%.
Polymerase chain reaction
The PCR results showed that all S. aureus strains possessed nuc gene encoding thermostable nuclease characteristic to S. aureus genus. Moreover, in all isolates ileS2 gene was detected (Table 1).
Pulsed-field gel electrophoresis
The genetic analysis revealed two different PFGE banding patterns (patterns A and B) and two clusters (clusters 1 and 2) among the eight mupirocin-resistant isolates of S. aureus (Table 1 and Fig. 1).

Dendrogram of the PFGE patterns of the Staphylococcus aureus isolates resistant to mupirocin. PFGE, pulsed-field gel electrophoresis.
The effect of trans-anethole on antibacterial activity of mupirocin
Our study revealed that trans-anethole (0.5–2%) combined with mupirocin increased the growth inhibition zone diameter around the disk, when S. aureus strains were susceptible (reference strain) or resistant (isolates) to this antibiotic, tested without trans-anethole by disk diffusion method (Fig. 2). It was shown that the concentration of trans-anethole >2% reduced the growth of S. aureus strains and inhibition zone diameter was not measurable. Moreover, it has been also observed that 1% Tween 80 addition to the culture medium had no impact on inhibition zone diameter of strains.

Growth inhibition zone of mupirocin against S. aureus strains evaluated on MHA supplemented with 1% Tween 80 (control) and with 1% Tween 80 and various concentrations (0.5–2%) of trans-anethole. Inhibition zone diameter around mupirocin disk for strain no. 6 on MHA supplemented with 1% Tween 80
trans-Anethole (0.5–2%) added to MHA resulted in the increase of growth inhibition zones ranging from 30.5 ± 0.7 to 53.5 ± 0.7 mm and from 7.1 ± 8.9 to 20.5 ± 14.0 mm, respectively, for S. aureus ATCC 29213 (susceptible to mupirocin) and S. aureus isolates (resistant to mupirocin). It was also noted that two isolates (strains no. 3 and 8) were intermediately resistant to mupirocin, whereas one isolate (strain no. 6) was completely susceptible to this antibiotic when treated with trans-anethole at concentration of 2%.
The effect of trans-anethole (0.5–2%) on growth inhibition zone diameter was observed for the following isolates: 1 (6.0 ± 0.0 to 9.5 ± 0.7), 2 (6.0 ± 0.0 to 12.5 ± 0.7), 3 (6.0 ± 0.0 to 18.5 ± 0.7), 4 (6.0 ± 0.0 to 15.5 ± 0.7), 5 (6.0 ± 0.0 to 16.5 ± 0.7), 6 (10.5 ± 0.7 to 53.5 ± 0.7) (Fig. 2A, B), 7 (6.0 ± 0.0 to 14.5 ± 0.7), and 8 (9.5 ± 0.7 to 23.5 ± 0.7).
trans-Anethole enhances the mupirocin efficacy against S. aureus
Biofilm biomass reduction assay showed that in case of 75.0% isolates (1, 2, 4, 5, 6, and 8) the use of mupirocin at a MIC50 value increased biofilm biomass. Furthermore, the use of trans-anethole does not significantly influence formed biofilm biomass. Combination of a mupirocin MIC50 value and 2% of trans-anethole significantly decreased biofilm biomass. Highest reduction was observed for strains 2 (from 0.792 ± 0.079 to 0.154 ± 0.061), 3 (1.102 ± 0.43 to 0.3 ± 0.89), and 8 (0.987 ± 0.082 to 0.245 ± 0.078). Therefore, effect of this combination in case of reference mupirocin-susceptible strain (0.676 ± 0.098 to 0.630 ± 0.069) showed minimal changes in decrease of zone diameter. The results of biofilm formation are presented in Fig. 3.
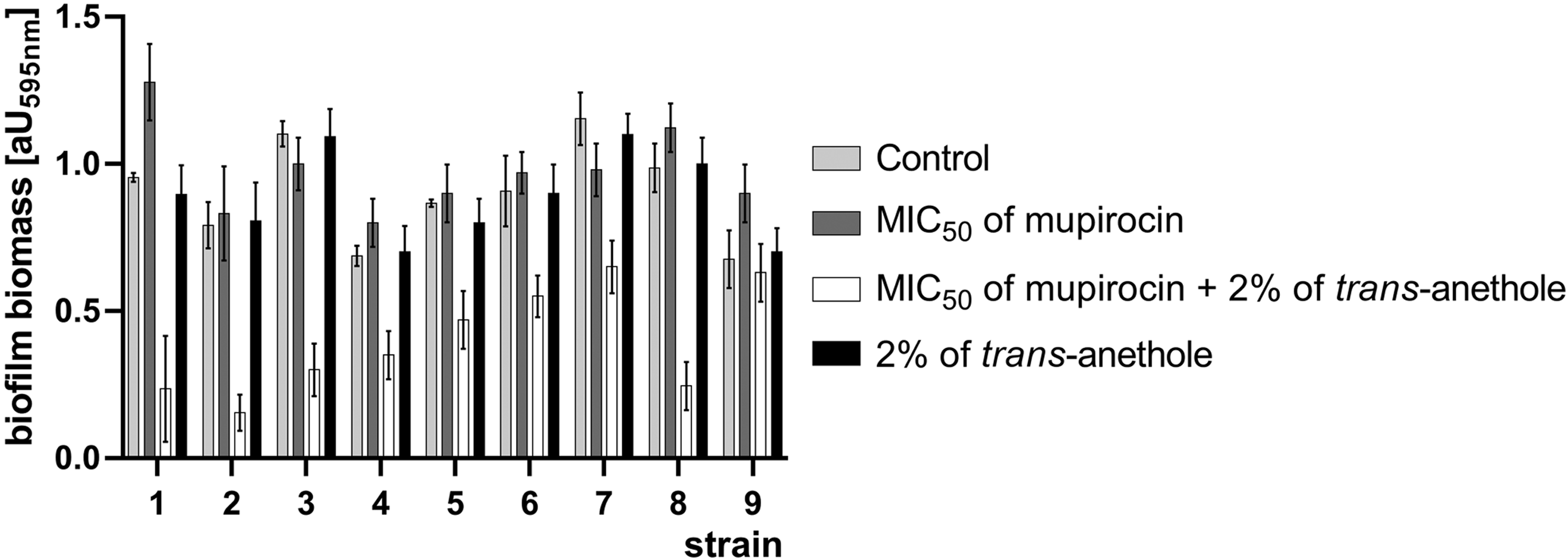
Effect of mupirocin and/or trans-anethole on biofilm biomass reduction.
Discussion
This study shows that combination of mupirocin with trans-anethole increases effectiveness of this commonly used antibiotic against S. aureus clinical mupirocin-resistant isolates. The use of disk diffusion method showed significant difference in inhibition zones around mupirocin disk. Trombetta et al. 19 proposed a mechanism of action of phenolic and alcoholic terpene. These lipophilic terpenes possibly migrate in the extracellular medium and interact with lipids of bacterial cell wall. These interactions increase cell membrane permeability and change the concentration of intracellular products. In addition, mupirocin is a passive-diffusion hydrophilic antibiotic so that intracellular concentration is considered to be directly correlated with membrane permeability. On the basis of the proposed mechanism we hypothesized that mupirocin and trans-anethole act in concert against mupirocin-resistant S. aureus.
Biofilm production, especially biofilm-related overproduction of exopolysaccharides, protects staphylococci in biofilm community against mupirocin activity. It should be noted that in most cases (strains 1, 2, 4, 5, 6, and 8) the use of mupirocin itself increased biofilm biomass. Sritharadol et al. 20 mentioned that subinhibitory concentration of mupirocin is able to induce the biofilm production rate. In contrast, trans-anethole used alone does not show significant biofilm eradication potency. Moreover, some terpenes, for example, thymol and menthol reduce biofilm of S. aureus. Kifer et al. 21 noted that two- to sixfold higher concentration than MIC is able to reduce almost 90% of produced staphylococci biofilm. The same authors showed that 1,8-cineole also exhibits weak antibiofilm properties. Nevertheless, terpenes antibiofilm properties are still poorly investigated and the use of mupirocin and 2% trans-anethole combination showed synergistic mode of action in terms of biofilm biomass reduction. Referring to the mentioned mode of action, terpenes could interact with the S. aureus surface protein, this interaction could disturb the quorum sensing system.22,23 Not all terpenes exhibit antibiofilm properties Kifer et al. 21 showed that mupirocin–menthol combination did not present antibiofilm activity but exerted antagonism. Authors speculate that this combination formed some kind of complex with exopolysaccharide-matrix of biofilm.
Mupirocin still remains the major effective tool in fighting MRSA carriage in community and nosocomial environment. Growing data on S. aureus physiology and its interaction with mupirocin enable the step forward in prediction of the potential weak points in existing strategies that target MRSA transmission. The same refers to the role of natural compounds that act synergistically with antibiotics. To the authors' opinion biofilm production appears an evident example of bacterial activity highly prone to be responsible for relatively high risk of MRSA recolonization. At the same time this is the novel topic that has not been intensively addressed yet.
This study indicated that the potential application of trans-anethole in combination with mupirocin can increase the mupirocin activity against MRSA and mupirocin-resistant strains. In addition, such combination is potentially capable to reduce biofilm formation and decrease the risk of MRSA recolonization.
Footnotes
Acknowledgments
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Disclosure Statement
No competing financial interests exist.