
Abstract
This study investigated the occurrence of virulence and antimicrobial resistance properties in Vibrio spp. isolated from hard-shelled mussel (Mytilus coruscus) marketed in Korea. A total of 32 Vibrio spp. isolates including Vibrio diabolicus (n = 16), Vibrio alginolyticus (n = 13), Vibrio parahaemolyticus (n = 2), and Vibrio harveyi (n = 1) were identified by gyrB gene sequencing. Every isolate grouped with their respective reference sequence excluding V. diabolicus and V. alginolyticus that were intermixed in a neighbor-joining phylogenetic tree. Every Vibrio spp. showed DNase and gelatinase activities. Phospholipase, slime, caseinase, lipase, β-hemolysis, and α-hemolysis were positive in 31 (96.8%), 31 (96.8%), 29 (93.5%), 28 (87.5%), 17 (53.1%), and 9 (28.1%) isolates, respectively. The PCR amplification of virulence genes displayed that V. alginolyticus-specific (toxR, tlh, VAC, and VPI) genes were detected in 23 (71.9%), 18 (56.3%), 21 (65.6%), and 26 (81.3%) isolates, respectively. V. parahaemolyticus-specific (toxR and tlh) genes were harbored by 2 (6.3%) and 2 (6.3%) isolates, respectively. The other virulence genes including ctxAB and hupO genes were observed in 23 (71.9%) and 16 (50.0%) isolates, respectively. The antimicrobial resistance was prevalent for amoxicillin (100.0%), ampicillin (100.0%), and streptomycin (37.5%) in disk diffusion test. Multiple antimicrobial resistance index ranged from 0.09 to 0.22. The extended-spectrum β-lactamase (ESBL) genes, blaCTX, blaTEM, blaSHV, and blaOXA were detected in 28 (87.5%), 13 (40.6%), 7 (21.8%), and 1 (3.1%) isolates, respectively. Non-ESBLs such as streptomycin resistance (strA.B), kanamycin resistance (aphA-IAB), and tetracycline resistance (tetE) genes were found in 5 (15.6%), 3 (9.4%), and 1 (3.1%) isolates. Class 1 integron-related Integrase 1 (intI1) gene was found in 8 (25.0%) isolates. Our results reveal that the consumption of raw mussels may cause a potential public health risks owing to the virulent and antimicrobial-resistant Vibrio spp.
Introduction
Seafood is a nutrient-rich healthy diet and source of vitamins, high-quality proteins, minerals, trace elements, polyunsaturated fatty acids, and other nutrients. 1 Among various kinds of seafood, mollusks comprise a large proportion owing to the taste preference and nutritional value. Mollusks are the second most marine aquaculture product in Korea. The country's self-sufficiency ratio of mollusks is 84.6% and a large portion of this consists of domestically produced mussels. 2 Hard-shelled mussel (Mytilus coruscus) (also known as Korean mussel) is one of the popular mussel species cultured in Korea. Koreans intake up to 251 g of mussels per day and they enjoy dishes of stir-fried, grilled, smoked, boiled, hot stew, and raw mussels.
Mollusks, especially bivalves can harbor different types of microorganisms from surrounding water because of their filter-feeding strategy. 3 Similar to other bivalve species, mussels are known as a mediator of many pathogenic bacteria. 4 Indigenous bacteria of marine environment particularly belong to the Vibrionaceae family and nonindigenous pathogenic bacteria (e.g., Shigella spp. and Salmonella spp.) are introduced into marine water by infected animals and humans. The ingestion of contaminated bivalve mollusks has been reported as a cause of many bacterial disease outbreaks that constitute a potential risk for consumers. 5
Vibrios are halophilic Gram-negative bacteria that are indigenous to the marine and brakishwater environments. 6 Up to date, there are ∼100 Vibrio spp. identified and most of these are considered as nonpathogenic species. However, several species are known to cause diseases in humans and other animals, and at least 13 species are known as pathogenic to human among these. 7 Vibrio cholerae, Vibrio vulnificus, and V. parahaemolyticus have been associated with food or waterborne diseases. 8 However, a few reports of infections caused by other Vibrio spp. such as V. alginolyticus, Vibrio diabolicus, and V. harveyi have also been described.9,10 Vibrios account for different types of human infections caused as a result of the consumption of raw or incompetently cooked or improperly processed seafood. 11
The pathogenicity of Vibrio spp. has been widely studied because of the presence of an assortment of virulence determinates. 12 The virulence factors are mainly related to toxin and hemolysin production. Thermostable direct-related hemolysin (TRH) and thermostable direct hemolysin (TDH) are considered as the most unique virulence factors of V. parahaemolyticus that are responsible for hemolytic activities. 13 In addition, ToxR helps them to colonize in the host's intestine and produces toxins. Thermolabile hemolysin (TLH) shows phospholipase activity and can lyse human erythrocytes. 14 Non-V. parahaemolyticus vibrios such as V. alginolyticus and V. diabolicus can harbor a variety of virulence determinants such as tlh, tdh, outer membrane protein, collagenase, and toxR. 15 Besides, there are other virulence-related extracellular enzymes such as lipase, gelatinase, protease, amylase, and DNase that can be observed in Vibrio spp. 16
Antimicrobial drugs are generally used to prevent bacterial infections in plants, humans, and other animals. However, the intensive use or misuse of antimicrobials in human, veterinary, agriculture, and aquaculture have caused antimicrobial contamination in the coastal environments. 17 The dissemination of antimicrobial resistance in a bacterial population results through mutation or resistance genes on mobile genetic elements (plasmid, transposon, and integron). 18 Of these, integrons play an important role in acquiring antimicrobial resistance as they can capture antimicrobial resistance gene(s) in their cassette region that contribute to multiple antimicrobial resistance.
Previously, several studies reported the occurrence and characterization of Vibrio spp. in bivalve mollusks including oysters, scallops, clams, and cockles, and so on.19–22 Only a few studies reported about the virulence properties and antimicrobial resistance patterns of Vibrio spp. from mussels in Korea.23,24 However, no study has been extensively carried out to characterize the virulence and antimicrobial resistance genes of different Vibrio spp. isolated from mussels in Korea. Thus, the aim of our study was to investigate the occurrence of virulent and multidrug-resistant Vibrio spp. isolated from mussels to raise public awareness about the potential public health risks related to the consumption of mussels.
Materials and Methods
Sampling of mussels
A total of 275 fresh mussels harvested in the western and southern coast of Korea were purchased from nine different retail shops during January to December 2018. Mussels were brought to the laboratory in sterilized containers and processed immediately. Each mussel was shucked and the flesh was transferred to the blender using sterile spatula. The flesh was blended in a sterile jar to make a homogenate that was used to isolate Vibrio spp. The sterile gloves were used and changed each time an individual was handled.
Bacterial isolation and biochemical identification
One gram of each homogenate was enriched in 9 mL of alkaline peptone water overnight at 37°C. One loopful of each enrichment was streaked on thiosulfate citrate bile salts sucrose (TCBS) agar (MB Cell, Los Angeles, CA) and incubated at 37°C for 24 h. Green or yellow color colonies in TCBS were presumed as Vibrio spp. and streaked on triple sugar iron agar (TSI; MB Cell). The positive isolates in TSI test (slant; alkaline or acidic and butt; acidic) were chosen for oxidase test. Oxidase-positive isolates were checked for the sensitivity by means of DD15 0129 (150 μg; Oxoid, Hampshire, United Kingdom) in vibriostatic disk diffusion test. Finally, the biochemically identified Vibrio spp. were selected for molecular identification.
Molecular identification and phylogenetic analysis
The genomic DNA of each biochemically identified Vibrio spp. was extracted by means of Exgene Cell SV kit (GeneAll, Seoul, Korea) according to the manufacturer's protocol. The species was identified by conventional PCR using MultiGene OptiMax Thermal Cycler (Labnet International, Inc., Edison, NJ). The gyrB housekeeping gene primers (F: AGCCAAACNAAAGAYAARYT and R: CGYARYTTRTCYGGRTTRTRYTC) were used in this study. 25 A total volume of 30 μL PCR mixture contained 3 μL of 10 × Taq reaction buffer (15 mM Mg2+) (Toyobo, Osaka, Japan), 1 μL of each forward and reverse primer (10 μM), 3 μL of dNTP (2 mmol/L), 0.3 μL Taq polymerase (5 U/μL), 20.7 μL of PCR water, and 1 μL (50 ng/μL) of template DNA. The amplified PCR products were checked by agarose gel electrophoresis on 1.5% (w/v) agarose gels. PCR products were sequenced after purification by Exgene PCR SV (GeneAll) kit. The sequences were analyzed by BLAST comparison in NCBI database.
A neighbor-joining phylogenetic tree was constructed by multiple sequence alignment with ClustalW function in the MEGA 7.0 sequence analysis software program. The gyrB sequences of mussel-borne Vibrio spp. and the reference sequences such as V. alginolyticus (AF007288), V. diabolicus (KF899126), V. harveyi (KF899127), V. parahaemolyticus (DS267854), and Escherichia coli KCTC 2441 (EU014649) as the outgroup taxa were used to construct the tree. The phylogenetic tree was created by neighbor-joining method and 1,000 bootstrap replicates were used to calculate tree topology robustness.
Phenotypic pathogenicity tests
Thirty-two identified Vibrio spp. were assayed for seven phenotypic pathogenicity tests. Every isolate was maintained in tryptic soy agar (TSA; MB Cell) with 1% (w/v) NaCl used as a supplement for all phenotypic pathogenicity tests. The isolates were streaked on DNase agar (MB Cell) to observe DNase activity and 1 N HCl was added on to the colonies on the agar plate (MB Cell). The formation of a halo effect around the colonies was observed as positive for DNase production. 26 TSA was supplemented with 0.5% (w/v) skim milk and incubation was carried out at 37°C for 48 h for detecting caseinase production. The formation of clear zones around the colonies was selected as positive for caseinase activity. 27 Gelatinase activity was tested using gelatin medium with the addition of gelatin (120 g/L), peptone (5 g/L), and beef extract (3 g/L) in test tubes. After incubating at 37°C for 48 h, test tubes were refrigerated at 4°C for 30 min, the isolates that have hydrolyzed the gelatin medium was considered positive. 28 TSA was supplemented with 5% (v/v) egg yolk emulsion and 1% (v/v) Tween 80 to detect phospholipase and lipase activities, respectively. The presence of opaque halo effect around the colonies was considered positive for phospholipase and lipase activities. 29 Slime production was assessed by inoculating colonies on Congo red agar. The changes of the medium color and black colonies were considered a positive reaction for slime production. 30 Blood agar (MB Cell) with sheep blood supplement was used to observe hemolysis activity. The existence of a greenish coloration zone around the colonies demonstrated the α-hemolysis and clear zones around the colonies were considered as β-hemolytic activity. 31
Antimicrobial resistance patterns and multiple antimicrobial resistance index
Disk diffusion test was carried out using 22 antimicrobials belonging to 11 antimicrobial classes as follows: penicillins: ampicillin (10 μg), amoxicillin (30 μg), piperacillin (100 μg); cephalosporins: cephalothin (30 μg), cefoxitin (30 μg), ceftriaxone (30 μg), cefotaxime (30 μg); carbapenems: imipenem (10 μg); quinolones: nalidixic acid (30 μg), ciprofloxacin (5 μg), ofloxacin (5 μg); lipopeptides: colistin sulfate (10 μg); macrolides: erythromycin (15 μg); aminoglycosides: streptomycin (10 μg), gentamicin (10 μg), kanamycin (30 μg), amikacin (30 μg); tetracyclines: oxytetracycline (30 μg), tetracycline (30 μg); folate pathway inhibitors: trimethoprim–sulfamethoxazole (25 μg); phenicols: chloramphenicol (30 μg); and ansamycins: rifampicin (5 μg).
The antimicrobial susceptibility testing was carried out according to the standards of the Clinical and Laboratory Standards Institute. 32 The multiple antimicrobial resistance (MAR) index value was calculated (MAR index = a/b, where “a” designates the number of antimicrobials the isolate was resistant to, and “b” designates the total number of antimicrobials the isolate was tested against). 33
Detection of virulence genes, antimicrobial resistance genes, and class 1 integron
The occurrence of 10 virulence genes, 15 antimicrobial resistance genes, and integron-related genes was observed by conventional PCR with MultiGene OptiMax Thermal Cycler (Labnet International, Inc.) (Tables 1 and 2). A total volume of 30 μL PCR mixture contained 3 μL of 10 × Taq reaction buffer (15 mM Mg2+; Toyobo), 3 μL of dNTP (2 mmol/L), 0.3 μL Taq polymerase (5 U/μL), 1 μL of each forward and reverse primer (10 μM), 1 μL of template DNA (50 ng/μL), and 20.7 μL of PCR water. PCR products were checked on 1.5% (w/v) agarose gels by agarose gel electrophoresis.
Oligonucleotide Primers Used to Detect Virulence Genes of Vibrio spp.
Oligonucleotide Primers Used to Amplify Antimicrobial Resistance Genes of Vibrio spp.
Results
Identification of Vibrio spp. and phylogenetic analysis
According to the gyrB gene sequencing, the Vibrio isolates were identified as four species, including V. diabolicus (n = 16), V. alginolyticus (n = 13), V. parahaemolyticus (n = 2), and V. harveyi (n = 1).
A neighbor-joining phylogenetic tree that was created using the sequences showed two major clusters (Fig. 1). One cluster included V. diabolicus, V. alginolyticus, and V. harveyi, whereas the other cluster included only V. parahaemolyticus. Every isolate grouped with their respective reference sequence excluding V. diabolicus and V. alginolyticus that appear intermixed in a single clade.

Neighbor joining phylogenetic tree based on GYRB gene sequences of Vibrio spp. Isolated from mussels. VA, Vibrio alginolyticus; VD, Vibrio diabolicus; VH, Vibrio harveyi; VP, Vibrio parahaemolyticus, and reference sequences have been obtained from the GenBank database.
Pathogenic virulence factors
Phenotypic virulence factors were analyzed among the tested isolates by seven pathogenicity tests including DNase, caseinase, gelatinase, lipase, phospholipase, slime formation, and hemolysis activity (Table 3).
Phenotypic Pathogenicity Profile of Vibrio spp. Isolated from Mussels (Mytilus coruscus)
All isolates showed 100% positivity for DNase and gelatinase activities. Of interest, except one isolate, all Vibrio spp. isolates were positive for phospholipase and slime production. Caseinase and lipase activities were observed in 29 (93.5%) and 28 (87.5%) isolates, respectively. In the case of hemolysis activity, β-hemolysis and α-hemolysis were seen in 17 (53.1%) and 9 (28.1%) isolates, respectively.
Antimicrobial susceptibility patterns and MAR index
The antimicrobial susceptibility patterns of Vibrio spp. are given in Fig. 2. All isolates were resistant to at least two antimicrobials belonging to ≥1 structural class.
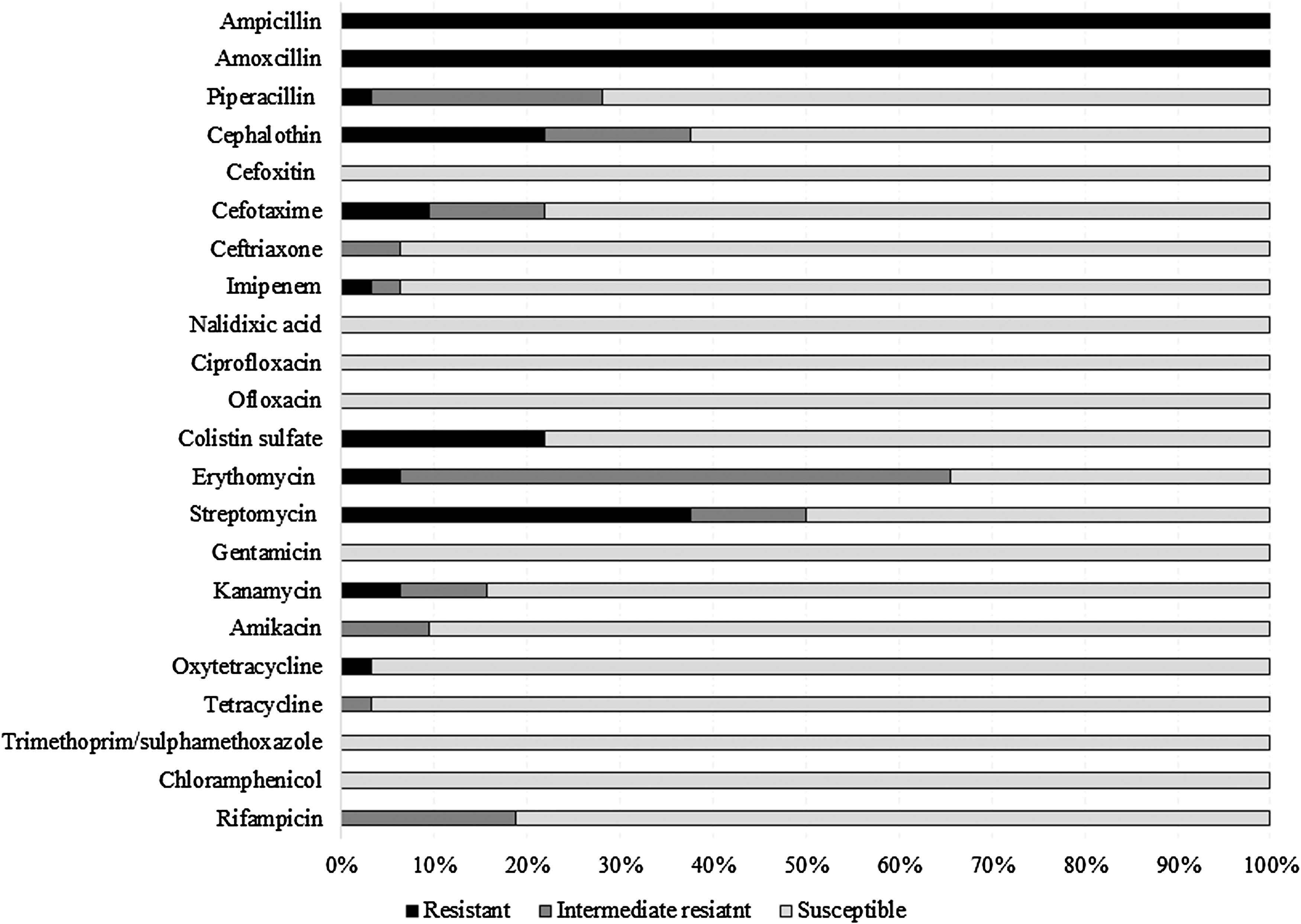
Antimicrobial susceptibility profile of Vibrio spp. isolated from mussels.
The resistance was prevalent for amoxicillin (100.0%), ampicillin (100.0%), and streptomycin (37.5%). The resistance to cephalothin, colistin sulfate, cefotaxime, erythromycin, kanamycin, oxytetracycline, imipenem, and piperacillin were 21.9%, 21.9%, 9.8%, 6.3%, 6.3%, 3.1%, 3.1%, and 3.1%, respectively. The isolates showed intermediate resistance to piperacillin (25.0%), cephalothin (15.6%), cefotaxime (12.5%), ceftriaxone (6.25%), imipenem (3.1%), erythromycin (59.4%), streptomycin (12.5%), kanamycin (9.4%), amikacin (9.4%), tetracycline (3.1%), and rifampicin (18.8%). All isolates were susceptible to cefoxitin, nalidixic acid, ciprofloxacin, ofloxacin, gentamicin, trimethoprim–sulfamethoxazole, and chloramphenicol. The MAR index values ranged from 0.09 to 0.22 (Table 4).
Antimicrobial Resistance Determinants of Vibrio spp. Isolated from Mussels
AMOX, amoxicillin; AMP, ampicillin; CEF, ceftriaxone; COL, colistin sulfate; E, erythromycin; ESBL, extended-spectrum β-lactamase; IMP, imipenem; KAN, kanamycin; KF, cephalothin; MAR, multiple antimicrobial resistance; OT, oxytetracycline; PIP, piperacillin; STR, streptomycin.
Prevalence of virulence and antimicrobial resistance genes
The occurrence of virulence-related genes of Vibrio isolates is presented in Table 5. V. alginolyticus-specific (toxR, tlh, VPI, and VAC) genes were observed in 23 (71.9%), 18 (56.3%), 21 (65.6%), and 26 (81.3%) isolates by PCR amplification. V. parahaemolyticus-specific (toxR and tlh) genes were amplified by 2 (6.3%) and 2 (6.3%) isolates, respectively. The other virulence genes including ctxAB and hupO genes were observed in 23 (71.9%) and 16 (50.0%) isolates, respectively. However, none of the Vibrio spp. isolates gave positive amplicons for the tdh and trh.
Prevalence of Virulence-Related Genes Among Vibrio spp. Isolated from Mussels
The extended-spectrum β-lactamase (ESBL) genes, blaCTX, blaTEM, blaSHV, and blaOXA were detected in 28 (87.5%), 13 (40.6%), 7 (21.8%), and 1 (3.1%) isolates, respectively (Table 4). The streptomycin resistance gene (strA.B) and kanamycin resistance (apha-IAB) genes were present in 3 (9.4%) and 5 (15.6%) isolates, respectively. Tetracycline resistance gene, tetE was detected only in 1 (3.1%) isolate. Class 1 integron-related integrase 1 gene was found in 8 (25.0%) isolates.
Discussion
Mollusks are considered important proportion of global seafood consumption. Mussel is one of the popular commercial seafood species worldwide, and in Korea. 47 There is a high market demand for mollusks such as scallops, oysters, cockles, mussels, and abalones. However, they are highly prone to natural occurring bacteria that are abundantly found in the aquatic environment. Therefore, this study characterized the Vibrio spp. in marketed mussels. As the phenotypic pathogenicity and antimicrobial susceptibility patterns were revealed, virulence and antimicrobial resistance determinants were distinctive among the Vibrio isolates.
V. diabolicus was the most prevalent species followed by V. alginolyticus among the isolates. V. diabolicus and V. alginolyticus are two closely related species that often share the same phylogenetic characteristics. The predominance of V. alginolyticus was observed in green mussels (Perna viridis) and oyster (Crassostrea gigas). 19 In addition, V. alginolyticus was frequently isolated from the coastal environment of Korea in a previous study. 48
The secretion of extracellular enzymes of Vibrio spp. has been extensively studied because of their active involvement in potential virulence.28,49 The extracellular enzymes are known as indicators of health risks related to Vibrio spp. isolated from clinical, environmental, and food sources. 50 The production of extracellular enzymes was also reported in our study. DNase and gelatinase activities were observed in every isolate and caseinase, phospholipase, and lipase activities were detected by most of the mussel-borne Vibrio spp. isolates. The extracellular DNase enzyme contributes to DNA hydrolysis and propagation of the bacterial strains. Gelatinase and caseinase enzymes have been associated with gelatin and protein degradation, respectively. 51 Lipase plays an important role in nutrient acquisition by the degradation of lipid membrane of host cells, whereas the phospholipase entails in hemolysis and lecithinase activities.52,53
Slime production is related to bacterial adherence in the specific host that acts against phagocytosis, antimicrobial action, and chemotaxis. 54 The slime production was observed in every species except V. parahaemolyticus. Similar to our study, Ben Abdallah et al. also detected nonslime-producing V. parahaemolyticus strains. 55 Hemolysis activity is one of the important virulent factors that helps in the intercellular growth of pathogenic bacteria. 56 α-Hemolysis or partial hemolysis reduces the host resistance by the inhibition of the phagocytic activity, whereas β-hemolysis or complete hemolysis causes bacterial infection by depleting iron from red blood cells and makes it able for the growth of bacteria.57,58 More than 50.0% of the Vibrio isolates were β-hemolytic. In contrast, ∼15.8% of the V. parahaemolyticus strains isolated from mussels and oysters showed β-hemolytic activity. 59
Collagenase plays an essential role in the wound infection as it influences the distribution of bacteria by the digestion of protein components of the extracellular matrix. 60 The V. alginolyticus-related collagenase gene, VAC was the most prevalent among the mussel-borne isolates. The VAC gene was highly present in Vibrio spp. from Yesso scallop (Patinopecten yessoensis). 20 In another study, the existence of collagenase was found in 71.4% of the V. alginolyticus isolated from green mussel (Perna viridis). 61
The continuous and widespread use of antimicrobials in aquaculture favors the growth of a variety of antimicrobial-resistant isolates and the distribution of resistance genes within the bacterial population. 62 The antimicrobials that have been used in this study are the antimicrobials used for the treatment of Vibrio infections, including cefotaxime, amikacin, gentamicin, tetracycline and trimethoprim–sulfamethoxazole.63,64 Some of these antimicrobials are extensively used in aquaculture including chloramphenicol and oxytetracycline. 65
The resistance to β-lactam antimicrobials was exhibited by the mussel-borne isolates, which is in agreement with previous studies that noted the resistance to β-lactam antimicrobials among the Vibrio spp. from seafood. 19 The ESBL gene, blaCTX was recorded as the most prevalent (87.5%) among the antimicrobial resistance genes. The production of ESBL facilitates the isolates to be resistant against β-lactam antimicrobials. ESBLs are one of the beta-lactamase enzyme groups that are known to hydrolyze penicillins, extended-spectrum cephalosporins and carbapenems. The ESBL production is commonly associated with Gram-negative bacteria. 66 ESBLs are frequently identified in Vibrio spp. isolated from different sources such as clinical, environmental, food sources. 23 Zanetti et al. found that 88.9% of the Vibrio isolates from marine or coastal environments showed resistance to β-lactams. 67 However, the tenacious exposure of bacterial isolates to β-lactams has induced dynamic production and mutation of β-lactamases among the bacterial population, expanding their action even to the newly developed β-lactam antimicrobials. 68
The strAB and aphA-IAB genes are non-β-lactamase genes that encode streptomycin and kanamycin resistance mechanisms. The isolates that were positive for strA.B and aphAI-IAB genes showed susceptibility to streptomycin and kanamycin in the disk diffusion test. This discrepancy might be attributed owing to the phenotypic and genotypic expressions of antimicrobial resistance characteristics. 69
Integrons are considered as the mobile genetic elements that can cause the dissemination of antimicrobial resistance genes among the bacteria. 62 Among different classes of integrons, class 1 integron is most frequently detected in Vibrionaceae isolated from clinical and livestock sources. 70 In this study, we could detect class 1 integron from 25.0% of the isolates. Integrons contain site-specific recombination sites that can integrate and exchange specific DNA elements called gene cassettes. 62 But in our study, we could not amplify any class 1 integron gene cassette. It is suggested that the absence of gene cassettes in intI1-positive isolates could be because of the lack of resistance genes in gene cassette region or absence of 3′ conserved region or lack of hybridization site of primers or existence of early stop codon in the sequence.71,72
Our study demonstrated that mussel serves as the source of Vibrio spp. with a high prevalence of V. diabolicus and V. alginolyticus. The Vibrio spp. isolates harbored different virulence and antimicrobial resistance determinants. Consumption of mussels can cause human health risks and the high rates of resistance to antimicrobials could be in the treatment of Vibrio spp. infections. Therefore, proper handling and preparation of raw or improperly processed seafood are very important to reduce the public health risks. In addition, it is suggested that continuous monitoring of potential pathogenic Vibrio spp. is necessary to reduce the health risk related to seafood consumption.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.