The GDP-6-deoxy-α-d-manno-heptose is a key building block molecule in constructing lipopolysaccharide of Gram-negative bacteria. Therefore, blockage of the biosynthesis pathway of GDP-6-deoxy-α-d-manno-heptose is lethal or increases antibiotics susceptibility to pathogens. In this study, we assayed d-glycero-α-d-manno-heptose-1-phosphate guanylyltransferase (HddC) from Yersinia pseudotuberculosis (Yp) using an efficient assay method supplying its natural substrate. Using the method, 102 chemical compounds were tested to search inhibitory compounds and electrospray ionization mass spectrometry was used to detect the HddC from Y. pseudotuberculosis (YpHddC) reaction product, GDP-d-glycero-α-d-manno-heptose. Interestingly, one promising lead, ethyl 5-({[(5-benzyl-1, 3, 4-oxadiazol-2-yl) thio] acetyl} amino)-4-cyano-3-methyl-2-thiophenecarboxylate (Chembridge 7929959), was discovered. The inhibitory activity of the lead compound against YpHddC has been proven by blocking its nucleotidyltransferase activity transferring the GMP moiety to α-d-mannose-1-phosphate (αM1P). Chembridge 7929959 shows that the half maximal inhibitory concentration (IC50) is 0.222 μM indicating its affinity with αM1P.
Introduction
Yersinia pseudotuberculosis, a Gram-negative bacterium, infects various animals and humans.1Y. pseudotuberculosis mainly causes zoonotic infection in various hosts. In humans, it has been related with food-borne infection.2 The main clinical expressions of this bacterium are fever, abdominal pain similar to appendicitis, and diarrhea at times.3 Although the Y. pseudotuberculosis infection rarely has led to septicemia, it usually causes septicemia in patients with underlying diseases such as hepatic cirrhosis, diabetes, or iron overload.4 The distribution of Y. pseudotuberculosis infection causing yersiniosis is worldwide. Yersiniosis frequently occurs in winter due to growth characteristics of Y. pseudotuberculosis being enhanced in low temperatures.5 Although outbreaks of yersiniosis have been repeatedly reported in Europe, a significant occurrence was also recorded in the Aomori, Japan in the early 1990s. In Korea, there have been several septicemic cases of Y. pseudotuberculosis infections.6
Lipopolysaccharide (LPS) is a main component of the outer membrane that plays a critical role in host–pathogen interactions with the innate immune system.7 LPS layer plays as a robust barrier against toxic antibiotics during infection.8 Regardless of bacterial types or sites of infection, bacterial adaptive changes are caused by modulation of LPS synthesis. In general, these changes induce an immune system evasion resulting in increasing antimicrobial resistance and persisting inflammation.7 In most Gram-negative bacteria, glycero-manno-heptose and its derivatives are significant components of LPS and the mutant microbes lacking these heptoses result in a deep-rough phenotype sensitive to hydrophobic agents.9,10 The majority of glycero-manno-heptose in LPS are derived from ADP-l-glycero-β-d-manno-heptose biosynthesis pathway. Therefore, enzymes involved in the ADP-l-glycero-β-d-manno-heptose biosynthesis producing these molecules have been studied as targets for suggestion of new classes of antibiotics.11 However, GDP-d-glycero-α-d-manno-heptose (GDP-d-α-d-heptose), which is the product of d-glycero-α-d-manno-heptose-1-phosphate guanylyltransferase (HddC), a precursor molecule, becomes the O antigens or capsules in several species.12–15 The three previous steps are catalyzed by sedoheptulose-7-phosphate isomerase, d-glycero-α-d-manno-heptose-7-phosphate kinase, and d-glycero-d-manno-heptose-1, 7-bisphosphate phosphatase.16,17
Although, HddC can be used as a template to design novel antibiotics, no trial has been reported yet. Among various microbes, we selected Y. pseudotuberculosis to study with because it had been published that 6-deoxy-α-d-manno-heptose is essential for the proper construction of the repeating O antigens in this species.18 Therefore, HddC from Y. pseudotuberculosis (YpHddC), which synthesizes GDP-d-α-d-heptose has been studied. In the 6-deoxy-α-d-manno-heptose biosynthesis pathway from Y. pseudotuberculosis, the precursor molecule of 6-deoxy-α-d-manno-heptose is from GDP-6-deoxy-α-d-manno-heptose (GDP-6dd-heptose). The nucleotide-activated molecule is formed with two catalytic enzymes sequentially, GDP-mannose 4,6-dehydratase (WcbK) and reductase (WcbJ) (Fig. 1).16
The schematic diagram of the biosynthesis pathways of GDP-6dd-heptose in Burkholderia pseudomallei. GDP-6dd-heptose, GDP-6-deoxy-α-d-manno-heptose.
Recently, our laboratory reproved the first crystal structure of the HddC family has been determined.19 It was YpHddC and revealed its strong structural similarity to those of cytidylyltransferase and uridylyltransferase. Nevertheless, YpHddC uses GTP for catalysis rather than CTP and UTP. As a result, meaningful information is growing based on the structural and functional study. Therefore, YpHddC can be a good target enzyme to develop antibiotics against yersiniosis. In this article, we proved the molecular function of YpHddC and found a potent antibiotic candidate molecule using a one-step sequential assay method.
Materials and Methods
Gene cloning, protein expression, and purification
The pBT7 plasmid DNA embracing the YphddC gene coding for the YpHddC protein (NCBI Reference Sequence: WP_050090753.1) was synthesized and acquired from Bioneer (Daejeon, Republic of Korea). The transformation, overexpression, and purification of YpHddC were performed with the previously published method.19 The purified protein was buffer exchanged into 20 mM Bis-Tris pH 7.0 using Vivaspin 20 MWCO 10 kDa (GE Healthcare), a centrifugal device.
Identification of the activity of YpHddC in the GDP-6-deoxy-d-heptose pathway
The enzyme function of YpHddC was tested by using a previously reported protocol.11 The enzymes (except YpHddC) involved in the GDP-6-deoxy-d-heptose pathway from Burkholderia pseudomallei were used to identify the molecular function of YpHddC. The proper condition for YpHddC was examined and the best condition was established by analyzing with electrospray ionization mass spectroscopy (MS).
Chemical screening with a one-step sequential assay method
The screening of about one hundred chemical compounds (Supplementary Table S1) was performed with a one-step sequential assay method.11 The method has been designed previously to assay chemical compounds over target enzymes, of which the natural substrates are difficult to purchase. The same protocol, a convenient one-tube one-step assay, was used as published.11 Probability of inhibitory function of each chemical was investigated by detecting the peak of GDP-4-keto-6-deoxy-α-d-manno-heptose (m/z 616), the product from the GDP-6dd-heptose biosynthesis pathway. The protocol of the MS analyses was also done with an Agilent 1290 Infinity LC Systems (Agilent, Santa Clara, CA). The high performance liquid chromatography (HPLC) system was connected to an Agilent G6550A Accurate-Mass Q-TOF (Agilent) at the Korea Basic Science Institute (Western Seoul Center) equipped with an electrospray ionization source operating in negative ion mode. The parameters being used were optimized as follows: capillary voltage, 3,500 V; nebulizer pressure, 35 psi; drying gas flow rate, 14 L/minute; gas temperature, 498 K; octopole radiofrequency, 750 V; fragment voltage (insource CID fragmentation), 380 V. Liquid Chromatography-Mass Spectrometry (LC-MS) accurate mass spectra were recorded across the range m/z 100–1,700. The accurate mass calibration was carried out over a mass range of m/z 112.95855–2533.8923.
Steady-state kinetics
A colorimetric assay method with pyrophosphatase20 was used to study steady-state kinetics of YpHddC transferring the GMP moiety from GTP to d-glycero-α-d-manno-heptose-1-phosphate. The reaction products are GDP-d-glycero-α-d-manno-heptose and pyrophosphate (PPi). α-d-Mannose-1-phosphate (αM1P) purchased from sigma was used as a substrate instead of d-glycero-α-d-manno-heptose-1-phosphate. PPi was catalyzed by inorganic pyrophosphatase (IPP) and measured using the malachite green method.21 A color reagent used was a mixture of ammonium molybdate [(NH4)6Mo7O24], malachite green solution, and Tween 20. The mixture was filtered through a 0.20 μm PVDF syringe filter (Younginfrontier, Inc., Seoul, Republic of Korea) and allowed to stand at room temperature for 1 hour before use. For the kinetic studies, reaction mixtures, including 50 mM Tris-HCl (pH 7.5), 10 mM MgCl2, 0.04 unit IPP, and 0.01 mg/mL YpHddC with different concentrations of GTP (0.0039–1 mM) at the constant concentration of αM1P (1 mM) and with different concentrations of αM1P (0.0039–2 mM) at the constant concentration of GTP (0.25 mM) were used. After incubation at 25°C for an hour, 40 μL of reaction mixtures were mixed with the malachite reagent (160 μL). The mixtures were left for standing for 15 minutes to develop the color. The standard curve was plotted and instruments used were as published.
Half-maximal inhibitory concentration
A colorimetric assay method with pyrophosphatase20 was also used to study the dose-dependent inhibitory effect of ethyl 5-({[(5-benzyl-1, 3, 4-oxadiazol-2-yl) thio] acetyl} amino)-4-cyano-3-methyl-2-thiophenecarboxylate (Chembridge 7929959) from ChemBridge Corp. The 40 μL of reaction mixtures containing 50 mM Tris-HCl (pH 7.5), 10 mM MgCl2, and 0.01 mg/mL YpHddC with different concentrations of Chembridge 7929959 (0.0098–40 μM) were incubated at 25°C for an hour. The reaction mixtures lacking YpHddC with the same concentration of Chembridge 7929959 used as above were also incubated at 25°C for an hour and measured as blanks. The reaction was initiated by adding 1 mM αM1P and 0.25 mM GTP and stood for an hour. After incubation, 160 μL of the same malachite reagent used above was added and left for standing for 15 minutes. A microplate spectrophotometer (Spectramax 190; Molecular Devices Corporation, Sunnyvale, CA) was used to measure the absorbance at 660 nm. The percentage inhibition (%Inhibition) was calculated based on the difference in absorbance between the reaction mixtures with and without Chembridge 7929959. The half-maximal inhibitory concentration (IC50) value of Chembridge 7929959 was calculated by a nonlinear regression analysis using GraphPad Prism 7.03.
Results
Expression and protein purification
YpHddC containing a noncleavable N-terminal His6-tag was overexpressed in soluble form by Escherichia coli BL21 (DE3) and purified by Ni2+-affinity chromatography. The pooled YpHddC was dialyzed with buffer condition 50 mM Tris–HCl pH 8.0, 100 mM NaCl, and 223 mM imidazole. With one-step purification, its purity was up to 95% on the sodium dodecyl sulfate-polyacrylamide sel electrophoresis (SDS-PAGE). A buffer exchange has been done for the pool with 20 mM Bis-Tris pH 7.0. The concentrated protein solution (28 mg/mL) was used for the enzyme assay.
Chemical screening with a one-step sequential assay method
The enzyme activity of YpHddC and the mass spectrum of the TktA/GmhA/HddA/GmhB/YpHddC/WcbK mixture were confirmed with MS analysis as the previous method (Supplementary Fig. S1a). The screening of ∼100 compounds was performed with the one-step sequential assay method.11 It is a novel, fast method to search potent inhibitors against the biosynthesis pathways composed of many functional enzymes. A list of compound names is provided in Supplementary Table S1. Whether the GDP-6dd-hptose biosynthesis pathways by compounds are inhibited was confirmed by analyzing the MS peak of the final product. As a result of the screening, one potent inhibitor, Chembridge 7929959, blocking the GDP-6dd-heptose pathway was detected (Supplementary Fig. S1b). To specify which protein on the pathway is inhibited with Chembridge 7929959, a step-by-step enzyme analysis was performed with 1 mM Chembridge 7929959. Consequently, the MS peak of the reaction product of YpHddC, GDP-d-glycero-α-d-manno-heptose, was not detected in the TktA/GmhA/HddA/GmhB/YpHddC mixture containing 1 mM Chembridge 7929959. Therefore, we elucidated that Chembridge 7929959 inhibits the enzyme activity of YpHddC.
Enzyme kinetics of YpHddC
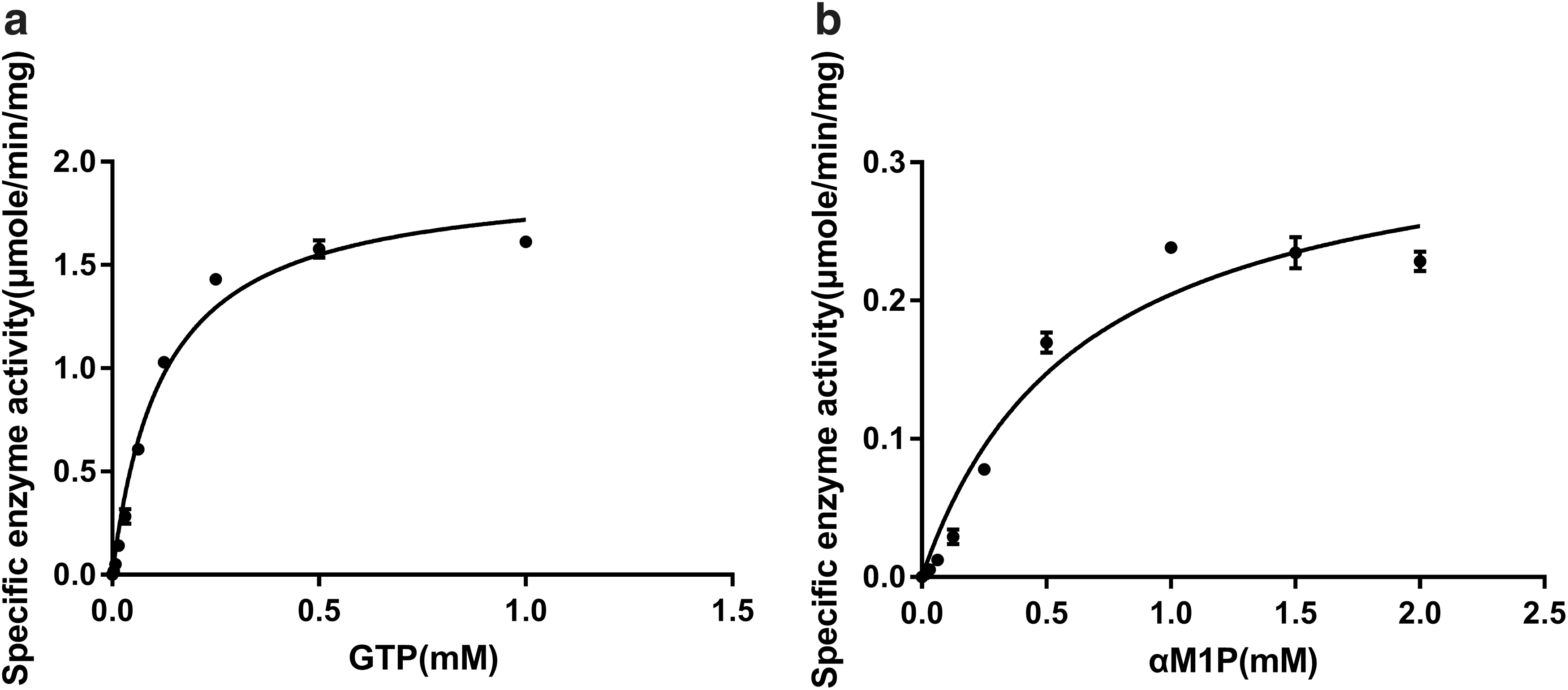
Since d-glycero-α-d-manno-heptose-1-phosphate, the biological substrate of YpHddC in the GDP-6dd-heptose biosynthesis pathway, is not easy to obtain, we carried out the enzyme kinetics of YpHddC with αM1P instead of the natural substrate. To determine the kinetic parameters for αM1P and GTP, enzyme assays were performed with various concentrations of GTP at a constant αM1P concentration (1 mM) and with various concentrations of αM1P at a constant GTP concentration (0.25 mM). The graphs of the specific activities versus concentrations of αM1P and GTP are shown in Fig. 2. The graphs indicated that the maximum enzyme activity was obtained at 0.25 mM GTP in the presence of 1 mM αM1P and at 1 mM αM1P in the presence of 0.25 mM GTP. The kinetic parameter values of GTP at the steady-state were 193.1 ± 6.87 minute−1 for kcat and 0.12 ± 0.01 mM for Km at the constant concentration of αM1P (1 mM). Those of αM1P were 33.49 ± 2.64 minute−1 for kcat and 0.639 ± 0.13 mM for Km at the constant concentration of GTP (0.25 mM).
The specific enzyme activities of YpHddC. The enzyme activities with (a) various concentrations of GTP with 1 mM αM1P and (b) various concentrations of αM1P in the presence of 0.25 mM GTP. αM1P, α-d-mannose-1-phosphate; YpHddC, d-glycero-α-d-manno-heptose-1-phosphate guanylyltransferase from Yersinia pseudotuberculosis.
IC50 value of Chembridge 7929959
The inhibitory effect of Chembridge 7929959 against YpHddC was established by the MS analyses. To reconfirm the inhibitory effect and identify dose-dependent inhibition of Chembridge 7929959 on YpHddC, PPi, a byproduct of the reaction, was quantified with a pyrophosphatase-coupled colorimetric assay. The dose-dependent inhibitory effect of Chembridge 7929959 was estimated at the saturated substrate concentrations (0.25 mM GTP and 1 mM αM1P). The results were plotted as log inhibitor concentration versus percentage inhibition of absorbance (Fig. 3). IC50 value determined from the dose-dependent inhibitory curve of Chembridge 7929959 is 0.222 μM.
A dose-dependent inhibitory curve for the inhibition of YpHddC by Chembridge 7929959.
Discussion
Yersiniosis is a foodborne infection mainly caused by Yersinia species. Due to increment of the disease, a study of the Yersinia species in various areas has been increasing.22–26Y. pseudotuberculosis is one of the bacteria causing yersiniosis. We have performed an antibacterial research targeting synthesis of bacterial LPS of Y. pseudotuberculosis. However, one of the main bottlenecks to continuous study in this field is purchasing commercially available actual substrates of target enzymes. Our antibacterial study against yersiniosis was also hampered by obtaining commercial biological substrates. Previously, however, we overcame the difficulty by employing the one-step sequential assay method. The assay method was applied to enzymes included in the biosynthesis pathways of ADP-l-glycero-β-d-manno-heptose and GDP-6dd-heptose from B. pseudomallei (Bp).11 The main merit of the method is the continual production of every substrate required for targeting enzymes and the concurrent assay of the whole enzymes in one system efficiently. To apply the method, five enzymes involved in the GDP-6-deoxy-d-heptose pathway from B. pseudomallei (Bp) were adopted and YpHddC was included instead of BpHddC. The method successfully provided the natural substrate of YpHddC and thus proved its molecular function. Based on the successful assay, a chemical screening for searching YpHddC inhibitor was carried out. An in-house chemical library composed of natural and chemical compounds was tested with the one-step sequential assay method. Finally, one promising chemical compound targeting YpHddC was detected and turned out to be Chembridge 7929959.
The inhibitory activity of Chembridge 7929959 against YpHddC was further confirmed again by obstructing the system for delivering the GMP moiety to αM1P. Since the natural substrate, d-glycero-α-d-manno-heptose-1-phosphate, for YpHddC was hard to get, a commercially available substrate was required as an alternative. Fortunately, αM1P can be used as an alternative to d-glycero-α-d-manno-heptose-1-phosphate for YpHddC. Therefore, αM1P was supplied as a substrate of YpHddC. It turned out to be working well and thus used for a kinetic study. With αM1P, enzyme kinetic parameters against the nucleotidyltransferase activity of YpHddC and the IC50 value were determined using a pyrophosphatase-coupled malachite green assay. The IC50 value of 0.222 μM of the Chembridge 7929959 was derived from the plot of log inhibitor concentration versus percentage inhibition of absorbance (Fig. 3). The IC50 value and kinetic parameters indicate that Chembridge 7929959 can be developed as a potent inhibitor against YpHddC. The multidrug resistance of pathogenic bacteria is threatening infected patients worldwide. Hence, a continuous requirement of novel antibacterial agents increases. In this study, we proved molecular function of YpHddC and established a method to screen inhibitory compounds. Thereby, the study showed that the enzymes involved in the GDP-6dd-heptose biosynthesis pathway can be a promising antibacterial target.
Chembridge 7929959 had been known to have low inhibitory activity against hepatitis B virus ribonuclease H.27 However, except the residual activity, no other molecular function of the compound has been known. Therefore, nullifying nucleotidyltransferase activity of YpHddC by Chembridge 7929959 is quite intriguing and can be used as a scaffold to develop antibacterial agents. Further research is being carried out to improve inhibitory activity of Chembridge 7929959 using its derivatives with X-ray crystallographic and biochemical techniques.
Ethics approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Footnotes
Disclosure Statement
The authors declare that they have no competing interests.
Funding Information
This research was supported by the Basic Science Research Programs, 2018R1D1A1B07050781 to D.H.S. and 2018R1D1A1B07050942 to M.-S.K., funded by the National Research Foundation of Korea granted by the Ministry of Education, Science, and Technology (MEST), Republic of Korea.
Supplementary Material
References
1.
GalindoC.L., RosenzweigJ.A, KirtleyM.L, and ChopraA.K.. 2011. Pathogenesis of Y. enterocolitica and Y. pseudotuberculosis in human yersiniosis. J. Pathog. 2011:182051.
2.
ByunJ.W., YoonS.S, LimS.K, LeeO.S, and JungB.Y.. 2011. Hepatic yersiniosis caused by Yersinia enterocolitica 4:O3 in an adult dog. J. Vet. Diagn. Invest. 2:376–378.
3.
VoskresenskayaE., SavinC., LeclercqA., TsenevaG., and CarnielE.. 2014. Typing and clustering of Yersinia pseudotuberculosis isolates by restriction fragment length polymorphism analysis using insertion sequences. J. Clin. Microbiol. 52:1978–1989.
4.
LjungbergP., ValtonenM., HarjolaV.P, Kaukoranta-TolvanenS.S, and VaaraM.. 1995. Report of four cases of Yersinia pseudotuberculosis septicemia and a literature review. Eur. J. Clin. Microbiol. Infect. Dis. 14:804–810.
5.
BradyM.F., and AnjumF.. 2019. Yersinia pseudotuberculosis. StatPearls, Treasure, Island, FL.
6.
HanT.H., PaikI.K, and KimS.J.. 2003. Molecular relatedness between isolates Yersinia pseudotuberculosis from a patient and an isolate from mountain spring water. J. Korean Med. Sci. 18:425–428.
7.
MaldonadoR.F., Sá-CorreiaI., and ValvanoM.A.. 2016. Lipopolysaccharide modification in Gram-negative bacteria during chronic infection. FEMS Microbiol. Rev. 40:480–493.
8.
NikaidoH., and VaaraM.. 1985. Molecular basis of bacterial outer membrane permeability. Microbiol. Rev. 49:1.
9.
ShlaesD., ShlaesJ., DaviesJ., and WilliamsonR.. 1989. Escherichia coli susceptible to glycopeptide antibiotics. Antimicrob. Agents Chemother. 33:192–197.
10.
VaaraM., and NurminenM.. 1999. Outer membrane permeability barrier in Escherichia coli mutants that are defective in the late acyltransferases of lipid A biosynthesis. Antimicrob. Agents Chemother. 43:1459–1462.
11.
ParkJ., LeeD., SeoE.K, RyuJ.S, and ShinD.H.. 2017. General assay for enzymes in the heptose biosynthesis pathways using electrospray ionization mass spectrometry. Appl. Microbiol. Biotechnol. 101:4521–4532.
12.
CzajaJ., JachymekW., NiedzielaT., LugowskiC., AldovaE., and KenneL.. 2000. Structural studies of the O-specific polysaccharide from Plesiomonas shigelloides strain CNCTC 113/92. Eur. J. Biochem. 267:1672–1679.
13.
WierzbickiI.H., ZielkeR.A, KorotkovK.V, and SikoraA.E.. 2017. Functional and structural studies on the Neisseria gonorrhoeae GmhA, the first enzyme in the glycero-manno-heptose biosynthesis pathways, demonstrate a critical role in lipooligosaccharide synthesis and gonococcal viability. Microbiologyopen, 6:e00432.
14.
WongA., LangeD., HouleS., ArbatskyN.P, ValvanoM.A, KnirelY.A, DozoisC.M, and CreuzenetC.. 2015. Role of capsular modified heptose in the virulence of Campylobacter jejuni. Mol. Microbiol. 96:1136–1158.
15.
DeShazerD., WaagD.M, FritzD.L, and WoodsD.E.. 2001. Identification of a Burkholderia mallei polysaccharide gene cluster by subtractive hybridization and demonstration that the encoded capsule is an essential virulence determinant. Microb. Pathog. 30:253–269.
16.
CuccuiJ., MilneT.S, HarmerN., GeorgeA.J, HardingS.V, DeanR.E, ScottA.E, Sarkar-TysonM., WrenB.W, and TitballR.W.. 2012. Characterization of the Burkholderia pseudomallei K96243 capsular polysaccharide I coding region. Infect. Immun. 80:1209–1221.
17.
ValvanoM.A., MessnerP., and KosmaP.. 2002. Novel pathways for biosynthesis of nucleotide-activated glycero-manno-heptose precursors of bacterial glycoproteins and cell surface polysaccharides. Microbiology, 148:1979–1989.
18.
KenyonJ.J., CunneenM.M, and ReevesP.R.. 2017. Genetics and evolution of Yersinia pseudotuberculosis O-specific polysaccharides: a novel pattern of O-antigen diversity. FEMS Microbiol. Rev. 41:200–217.
19.
KimH., ParkJ., KimS., and ShinD.H.. 2018. Crystal structure of d-glycero-α-d-manno-heptose-1-phosphate guanylyltransferase from Yersinia pseudotuberculosis. Biochim. Biophys. Acta Proteins Proteom. 1866:482–487.
20.
ShaS., ZhouY., XinY., and MaY.. 2012. Development of a colorimetric assay and kinetic analysis for Mycobacterium tuberculosisd-glucose-1-phosphate thymidylyltransferase. J. Biomol. Screen. 17:252–257.
21.
HarderK.W., OwenP., WongL.K, AebersoldR., Clark-LewisI., and JirikF.R.. 1994. Characterization and kinetic analysis of the intracellular domain of human protein tyrosine phosphatase beta (HPTP beta) using synthetic phosphopeptides. Biochem. J. 298:395–401.
22.
ReinhardtM., HammerlJ.A, KunzK., BaracA., NöcklerK., and HertwigS.. 2018. Yersinia pseudotuberculosis prevalence and diversity in wild boars in Northeast Germany. Appl. Environ. Microbiol. 84:e00675-18.
23.
KimJ., FukutoH.S, BrownD.A, BliskaJ.B, and LondonE.. 2018. Effects of host cell sterol composition upon internalization of Yersinia pseudotuberculosis and clustered β1 integrin. J. Biol. Chem. 293:1466–1479.
24.
WillcocksS.J., StablerR.A, AtkinsH.S, OystonP.F, and WrenB.W.. 2018. High-throughput analysis of Yersinia pseudotuberculosis gene essentiality in optimised in vitro conditions, and implications for the speciation of Yersinia pestis. BMC Microbiol. 18:46.
25.
BamyaciS., EkestubbeS., NordfelthR., ErttmannS.F, EdgrenT., and Å. Forsberg. 2018. YopN is required for efficient effector translocation and virulence in Yersinia pseudotuberculosis. Infect. Immun. 86:e00957-17.
26.
ChouikhaI., SturdevantD.E, JarrettC., SunY.C, and HinnebuschB.J.. 2019. Differential gene expression patterns of Yersinia pestis and Yersinia pseudotuberculosis during infection and biofilm formation in the flea digestive tract. mSystems, 4:e00217-18.
27.
TavisJ.E., ChengX., HuY., TottenM., CaoF., MichailidisE., AuroraR., MeyersM.J, JacobsenE.J, ParniakM.A, and SarafianosS.G.. 2013. The hepatitis B virus ribonuclease H is sensitive to inhibitors of the human immunodeficiency virus ribonuclease H and integrase enzymes. PLoS Pathog. 9:e1003125.
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.