
Abstract
Acinetobacter baumannii is a relevant opportunistic pathogen, and one of the main microorganisms responsible for outbreaks in nosocomial infections worldwide. Its pathogenicity is mainly due to its resistance to multiple antibiotics and to its ability to form biofilms on abiotic surfaces. The objective of this study was to characterize the biofilm formation cycle of A. baumannii isolated from a patient in a hospital and compare its antibiotic resistance with the planktonic cells. To study biofilm formation, the classical microtiter assay was used, with crystal violet staining and optical density reading to classify the type of biofilm. Also, the effect of gentamicin and colistin on bacterial biofilm was studied with an extra step of antibiotic addition. For the characterization of the different biofilm formation stages, the strain was grown on a coverslip, and the stain was made with a mixture of fluorophores markers to visualize the biofilm with a confocal laser microscope. It was possible to differentiate the A. baumannii biofilm formation stages. Through these observations, it was possible to estimate the time elapsed between each stage. As the strain was susceptible to colistin and gentamicin, both antibiotics were evaluated after the biofilm was formed. Neither antibiotics showed an effect on the eradication of A. baumannii biofilm.
Introduction
In recent years, the Infectious Diseases Society of America has emphasized the resistance to antibiotics by the bacteria Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter spp., which have been nicknamed as “the ESKAPE pathogens,” because of their ability to “escape” from the antibiotic action. 1
A. baumannii is an opportunistic Gram-negative pathogen, which is commonly found in intensive care units (ICUs) or surgery rooms, where the frequent use of antibiotics has allowed the selection of resistance against the vast majority of known antibiotics. It is known for its environmental persistence, surviving up to 5 months on abiotic surfaces. 2
Among the mechanisms of resistance to antibiotics, modification of the target site, enzymatic inactivation, the presence of efflux pumps and disturbances in the permeability of the outer membrane have been described in Gram-negative bacteria. 3 On the other hand, the biofilm formation by some microorganisms constitutes an alternative form of resistance to a wide variety of antibiotics and other antimicrobial agents.
The bacterial community development such as a biofilm and the expression of efflux pumps are mechanisms of resistance that began to been noticed in the last years and are associated with the difficulty of eradication of microorganisms by antibiotic treatment and lack, in some cases, of laboratory techniques that contribute to its detection. 4
Bacterial biofilms are bacterial communities irreversibly attached to a surface or between cells, embedded in a matrix of an extracellular polymer of own production composed of polysaccharides, proteins, extracellular DNA, and water channels.5,6
These communities have physiological and genetically differentiable attributes from planktonic forms, respect to their metabolism, growth rate, and gene transcription, which is proposed as a strategy of the bacterial population to survive, 5 as it can be reflected in their antibiotic resistance for example.
Biofilm formation occurs mainly in five stages with an initial adhesion of planktonic bacteria to different surfaces through flagella and fimbriae, among other factors. Then, the production of extracellular polymers, which leads to an irreversible adhesion occurs with the formation of microcolonies and maturation of the biofilm. The three-dimensional (3D) cell mass varies in its bioarchitecture constituting “mushroom-like” structures. Finally, a dispersion step occurs when the planktonic cells disengage from the biofilm, being able to colonize new sites.5–7
The objective of this work was to characterize the biofilm formation cycle of A. baumannii isolated from a hospital and compare the antibiotic resistance of mature biofilm with its planktonic counterpart. In the present work, we evaluated two in vitro static models to study the biofilm, one based in the classical crystal violet (CV) staining in microtiter plates and the other in image analyses from confocal microscopy and subsequently morpho-topological parameters calculation. The obtained results demonstrated that A. baumannii could form biofilms over the abiotic surface very fast and that antibiotic resistance changes in the biofilm bacteria compared with their planktonic counterpart. Considering these results, the development of fast techniques for the detection of this opportunistic pathogen and the assessment of antimicrobial resistance in biofilm compared with planktonic cells are necessary.
Materials and Methods
Bacterial strain and culture conditions
A. baumannii C100 strain was obtained from a clinical isolate of a tracheal secretion from a patient admitted to the ICU from Hospital de Clínicas, Montevideo, Uruguay. This strain was kept in Luria Bertani (LB) broth supplemented with glycerol (20%) at −80°C. The criteria for selecting strain C100 (aka HCA38*) were as follows: (1) it was selected from a collection previously well-characterized as strong biofilm forming strain; (2) the susceptibility to various antibiotics was studied by microdilution method using MicroScan system (Beckman Coulter, Pasadena) and interpreted according to the European Antimicrobial Susceptibility Testing Committee—EUCAST; (3) the presence of different resistance genes and sequence type had been previously determined. 8
A. baumannii was resistant to amikacin, ciprofloxacin, ceftazidime, cefepime, imipenem, meropenem, tobramycin, and trimethoprim-sulfamethoxazole, remaining susceptible to gentamicin (MIC ≤4 μg/mL) and colistin (MIC ≤0.5 μg/mL). C100 belonged to the sequence type ST79 and harbored blaOXA-51 and blaOXA-23. 8
Biofilm quantification through the CV technique
Bacterial ability to produce biofilm was assessed using a semiquantitative microtiter technique as previously described. 9 Briefly, the strain was grown overnight in LB at 37°C under static conditions. Twenty microliters from this culture were inoculated into 180 μL of LB in 96 flat-bottomed well, polystyrene microtiter plate and incubated for 48 hr at 37°C without shaking. 10 Planktonic bacteria were removed, and attached bacteria were washed three times with phosphate buffered saline (PBS). The plates were stained with 1% CV for 15 min at room temperature. Then the excess of dye was removed with subsequent PBS three washes, and CV was solubilized with 200 μL of 95% ethanol. The biofilm biomass was measured by optical density at 590 nm (OD590) using a Microplate Reader (Varioskan, Thermo Scientific). Strains were classified as follows: OD ≤ ODc = no biofilm producer; ODc < OD ≤ (2 × ODc) = weak biofilm producer; (2 × ODc) < OD ≤ (4 × ODc) = moderate biofilm producer; and (4 × ODc) < OD = strong biofilm producer; where ODc were control wells with media without bacteria. 11
Antibiotic biofilm assay
To assess the effect of gentamicin and colistin on bacterial biofilm, the classical biofilm quantification technique using CV in microtiter plates was performed with an extra step of antibiotic addition as previously described. 12 After 48 hr of biofilm formation, the planktonic cells were removed, fresh media with different antibiotic concentrations were added, and plates were incubated for another 24 hr. The different antibiotic concentrations were 1 and 2 μg/mL for colistin and 4, 8, and 16 μg/mL for gentamicin. All the measures were performed in triplicate. The means and standard errors were calculated for all experiments, and one-way ANOVA was applied to evaluate significant differences among the treatments.
Biofilm formation in static conditions
C100 strain was plated on MacConkey agar at 37°C and for 24 hr. Precultures were made from isolated colonies in LB broth, and incubated at 37°C for 24 hr.
To achieve the development of the biofilm in a static model, 30 mL of LB broth in plastic tubes containing a sterile coverslip were inoculated with 100 μL of the preculture. These tubes were incubated under static conditions at 37°C for different times (12, 16, 18, 21, 24, 28, and 32 hr).
Live/dead staining
Once the corresponding incubation time had elapsed for each tube, a stain was made with three markers to visualize the biofilm with a ZEEIS 800 Confocal Laser Microscope and Zen Blue 2.3 software. To perform the staining of the biofilm, each coverslip was removed from the tube, washed with PBS to remove the planktonic bacteria, and incubated in an opaque chamber for 15 min with the fluorophores mixture (5 μg/mL propidium iodide, 10 μg/mL Wheat Germ Agglutinin, and 10 μM Syto9 (Thermo Fisher Scientific)). Finally, bacteria were incubated with paraformaldehyde inside a chamber isolated from the light, to fix them on the coverslip. Each coverslip was washed with PBS, placed on a slide, and sealed with enamel.
Acquisition of images in confocal laser scanning microscope
Acquisition of 3D image stacks was performed using a ZEEIS 800 Confocal Laser Microscope and Zen Blue 2.3 software. The visualization was carried out with a 63 × oil immersion objective, 1.4 numerical aperture, and using three excitation lines of solid-state lasers 405, 488, and 561nm. Three random fields were selected in each coverslip, and the stacks were obtained using a step size of 0.3 μm between one image and the next on the z-axis. Image size was 1,024 × 1,024 pixels and pixel size of 0.070 μm.
Processing of images with IDL 7.2 and Fiji
The images were processed with IDL 7.2 (Interactive Data Language, ITT, CO) applying segmentation procedures as Threshold, Median, Fill remove, First deviation Kernal, visualization, 3D reconstruction, and determination of biofilm descriptors as previously described.7,13
After segmentation, 3D images of the biofilm were obtained. The quantity, volume of live and dead bacteria, and volume of the extracellular matrix in the different regions of interest were calculated. The hexagonal lattice (degree of compaction of the biofilm) was obtained from the two-dimensional model of distance from each stack. Data were processed with OriginPro 8 and analyzed with the Prism7 software.
Results
Biofilm quantification under different antibiotic concentrations
A. baumannii C100 was characterized as a strong biofilm producer strain (OD590 > 0.56, mean = 1.2) at 48 hr. This time was selected since we had compared 24 and 48 hr of biofilm formation and no significant differences were observed among both times. The effect of two antibiotics on the biofilm was assessed using the classical CV assay in microtiter plates after another 24 hr of incubation. As the strain was susceptible to colistin and gentamicin, we wondered whether both antibiotics could exert any impact on the biofilm formation. The results showed that neither colistin nor gentamicin had a significant effect on the biofilm (Fig. 1). Both antibiotics were unable to affect the biofilm formation even when A. baumannii C100 was susceptible to these antibiotics. However, reduction of the biofilm was observed at 1 μg/mL of colistin, but it was reverted at 2 μg/mL. In the case of gentamicin, we observed a reduction in the biofilm biomass that was dose dependent as the higher effect was observed in the highest antibiotic concentration. In all cases, the reduction observed was not significant compared with the biofilm without antibiotics and the strain kept inside the category of strong biofilm formation.
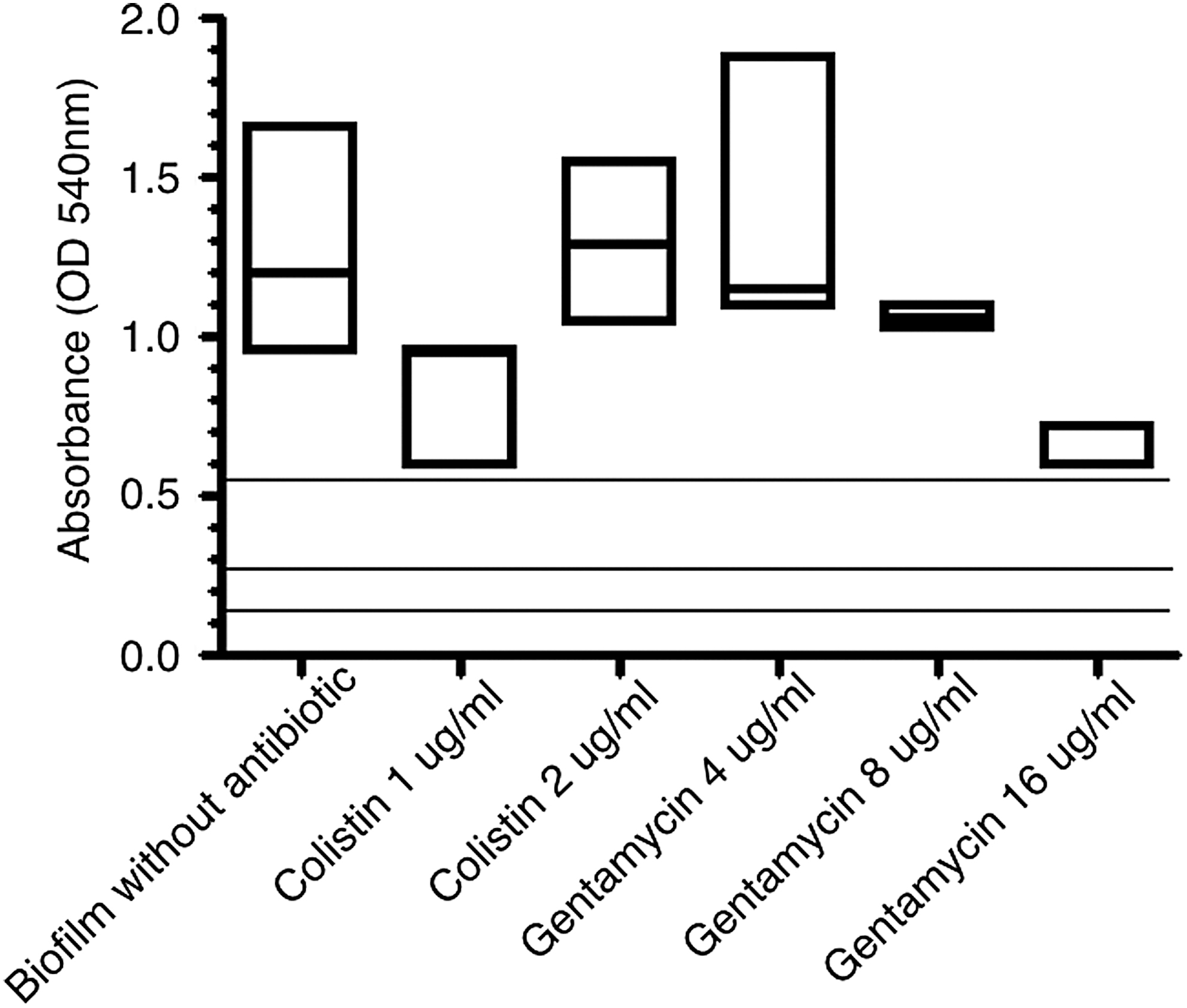
Effect of colistin and gentamicin on Acinetobacter baumannii biofilm. Box plot of the absorbance values, the line inside the box represents the median value. The lines at 0.14, 0.28, and 0.56 represents the values for the biofilm classification as no biofilm formation, weak and moderate biofilm respectively. Values above 0.56 were considered as strong biofilm formation.
A. baumannii biofilm evolution over time
After processing of the images obtained with IDL 7.2 it was possible to get morpho-topological data of the biofilm as the total number of live/dead bacteria, bacteria volume, and extracellular matrix volume (Fig. 2). Based on these data, we studied how the bacteria number (N/stack) evolved over time, as shown in Figure 2A. It can be observed that the greatest total bacterial number was observed at 18 hr after incubation and that after this peak, the number of bacteria decreased. In Figure 2B and C the bacteria and extracellular matrix volume were calculated for each time point, and the maximum volume was observed at 21 hr. Dead bacteria were quantified using live/dead staining, and as it is observed in Figure 2D, dead bacteria were present during all the evaluated period.

Biofilm morpho-topological parameters from confocal image analysis.
According to the obtained images, 3D reconstruction and morpho-topological parameters showed that the initial reversible adhesion of A. baumannii occurs after 12 hr (Fig. 3, Panel 12) and its irreversible adhesion and formation of microcolonies at 16 hr (Fig. 3, Panel 16). Macrocolonies and the stage of maturation of the biofilm were observed at 18 hr (Fig. 3, Panel 18) with the corresponding increase in the number and volume of bacteria and also in complexity and compaction, as seen in the lattice diagrams. As it was possible to observe through the lattice model, the compaction increased along time until 18–21 hr when some desegregation of the biofilm started and the distance between bacteria increased (Fig. 3, Panel 18 and 21). Bacterial dispersion was observed at 24, 28, and 32 hr (Fig. 3, Panel 24, 28, and 32) according to the results obtained using the lattice diagrams.
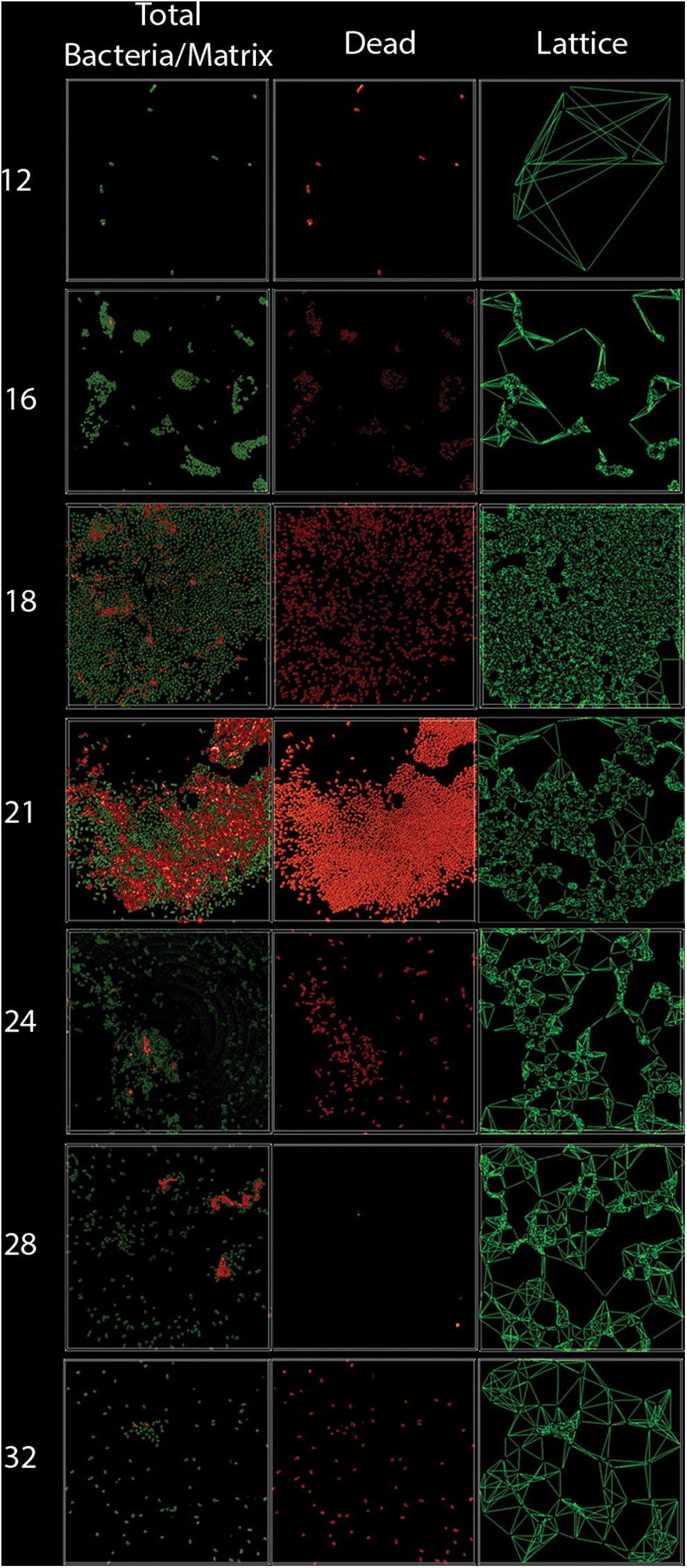
Three-dimensional reconstruction of biofilm during the time. Images of total bacteria (Syto9 staining), matrix (wheat germ agglutinin staining), dead bacteria (propidium iodide staining), and lattice two-dimensional model at 12, 16, 18, 21, 24, 28, and 32 hr. Total bacteria and matrix is observed in green, dead bacteria in red, and the lattice model is depicted as lines connecting one bacterium with the six more close ones. Color images are available online.
Discussion
Given that there are few and limited antecedents related to the characterization of A. baumannii biofilms from the clinical origin, the information obtained in this work may be valuable to understand the biofilm formation cycle and to look for biofilm eradication strategies.
In recent years, A. baumannii has rapidly developed resistance to antimicrobials, and multidrug-resistant strains have been isolated.14,8 Moreover, the World Health Organization published a list of antibiotic-resistant priority pathogens, where carbapenem-resistant A. baumannii was categorized in the critical group with the highest urgency for research and development new antibiotics. The strain used in the present work was isolated from a patient admitted to an ICU. The antibiotic resistance profile showed that bacteria were sensitive to colistin and gentamicin considering this strain as extensively drug-resistant according to Magiorakos et al. 15 The mechanism for acquired resistance to polymyxins reported in Gram-negative bacteria is based on the drug target modification. The lipid A, the anchor of the lipopolysaccharide (LPS), is modified by the addition of cationic groups such as 4-amino-L-arabinose (L-Ara4N), phosphoethanolamine (pETN), or galactosamine. 16 These changes affect the binding of colistin to the cell membrane and prevent disruption of the cell membrane integrity. 17 A. baumannii lacks all the genes required for L-AraN biosynthesis, 16 so the mechanism for resistance relies on pETN, which is mediated by the chromosomally encoded pmrCAB operon or due to mcr-1, a mobile colistin resistance gene which limits the use of this antibiotic for the treatment of multi-resistant microorganisms.18,19
Other colistin-resistance mechanisms have been described in A. baumannii, related with the complete loss of LPS, 20 the presence of efflux pumps, 21 or the disruption of the global regulator H-NS by insertion sequence ISAba125. 22 On the other hand, the use of aminoglycosides is still an option for A. baumannii treatment, usually in combination with β-lactam agents. There are also several mechanisms for aminoglycoside inactivation through N-acetylation, O-nucleotidylation, and/or O-phosphorylation. 23 Other resistance mechanisms to these antibiotics involve 16S rRNA methylation 24 and overexpression of different efflux pumps such as AdeABC 25 and AbeM, 26 and RND and MATE efflux pumps, respectively. The correlation between the presence of the different resistance mechanisms and the resistance levels achieved may vary depending on the expression levels. 27 Here, we report that gentamicin was unable to eradicate the biofilm, although a nonsignificant decrease in the biofilm biomass was observed.
Our results suggest that biofilm formation confers tolerance to A. baumanni to both colistin and gentamicin according to Ciofu et al. 28 Antibiotic tolerance in biofilm-forming bacteria has been widely studied, and several mechanisms have been described as a limitation of antibiotic diffusion through the matrix, horizontal transmission of genes, inactivation of the antibiotic by changes in metal ion concentrations and pH values, and the bacterial metabolic status.11,28 As it was reported by Gerson et al., further and deeper analysis of the mechanism of colistin resistance and/or tolerance is required. 23 A. baumannii strains are particularly resistant to desiccation, and it was reported that biofilm formation could influence this capacity. 30 The presence of dead bacteria during the entire biofilm formation as observed in the present work could protect the live ones from desiccation. The stratification of the biofilm could explain its capacity to withstand long period of starvation and desiccation in diverse environments including hospitals as it was previously reported. 10 Also, these dead bacteria could be a source of nutrients, and resistant genes for other microorganism highlighting the relevance of A. baumannii biofilms in biomedical context.
In comparison with other biofilm-forming microorganisms such as some Proteus mirabilis strains, which has a cycle of biofilm formation of 7-day in static conditions 7 or Pseudomonas aeruginosa between 1 and 4 days, 31 this A.baumannii strain has a considerably faster biofilm formation cycle in this static model. This can be taken as a base to consider antibiofilm bioactive strategies since it should have a faster effect than others focused on microorganisms that form slower biofilms.
In a previous report, we have shown that A. baumannii rapidly colonizes the respiratory and digestive tracts of patients admitted to an ICU. This colonization is strongly associated with subsequent infection of the lower respiratory tract. 32 Further studies are needed to determine whether this rapid kinetics of biofilm formation could be linked to rapid gastrointestinal and respiratory colonization of hospitalized patients. Moreover, A. baumannii was recently found as part of the oral microbiome, so the role of this bacteria in multispecies biofilm should be considered. Particularly, it was associated with Porphyromonas gingivalis the causing agent of periodontal diseases. 29 There is evidence that oral plaque could be a reservoir for potential respiratory pathogens; moreover the severity of the periodontal disease is linked to adverse respiratory conditions. 33
The development of antibiofilm materials is important for the design of clinical instruments. Some studies indicate that A.baumannii can form biofilms on a variety of different surfaces such as polycarbonate, a thermoplastic material that is often used in the construction of medical devices. A study conducted on different materials indicated that A.baumannii forms weak biofilms on glass. 34 On the other hand, other authors have demonstrated that clinical isolates can form biofilms on different substrates, including glass or plastic.34–36 Our results are in agreement with these reports as A. baumannii was able to form biofilms on glass in 21 hr.
These findings justify the need to continue studying the in vitro compatibility of medical materials that could be colonized by A. baumannii and enable its persistence in the hospital environment, 34 and the development of molecules that successfully interfere with the biofilm formation cycle.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by CSIC I + D 2017 grant.