
Abstract
Acinetobacter baumannii is the main species of the Acinetobacter genus; however, non-baumannii Acinetobacter (NBA) species causing infections have been described for the past years, as well as antimicrobial resistance. In this study, we describe the occurrence of two multidrug-resistant (MDR) IMP-1-producing Acinetobacter bereziniae isolates recovered from bloodstream infections in different patients but in the same intensive care unit among 134 carbapenem-resistant Acinetobacter screened. Antimicrobial susceptibility testing revealed resistance to carbapenems, extended spectrum, and antipseudomonad cephalosporins, amikacin, and trimethoprim-sulfamethoxazole. Both A. bereziniae isolates shared the same ApaI-pulsed-field gel electrophoresis (PFGE) pattern. Whole-genome sequencing of both isolates revealed that blaIMP-1 was embedded into an In86 Class I integron carrying also sul1, aac(6′)-31, and aadA genes. A new sequence type (ST1309 Pasteur) was deposited. The virulence genes lpxC and ompA, seen in A. baumannii, were detected in the A. bereziniae strains. Recognition of A. bereziniae causing invasive MDR infection underscores the role of NBA species as human pathogens especially in at-risk patients.
Introduction
A
Although carbapenem resistance in A. baumannii is mainly mediated by carbapenem-hydrolyzing class D β-lactamase (CHDL) OXA-enzymes, non-baumannii Acinetobacter (NBA) usually carry metallo-β-lactamases, with NDM, IMP, and VIM being the most frequently identified enzymes in both nosocomial and environmental samples.6–9 IMP was the first acquired metallo-beta-lactamase (MBL) described 10 and is currently spread worldwide.11–13 IMP enzymes may be detected in A. baumannii, but it is mainly harbored by NBA, such as A. radioresistens, A. ursingii, and Acinetobacter bereziniae, which are opportunistic and formerly known as environmental species. However, NBA species have risen as relevant pathogens, causing nosocomial infections in increasing frequencies for the past years.14–20 Since carbapenemase genes are usually associated with mobile genetic elements in NBA, those species can be implicated in horizontal gene transfer to more frequent clinical species, such as A. baumannii.
Among the NBA, A. bereziniae is considered an emerging pathogenic species and has been associated with some carbapenemase enzymes. As in other Acinetobacter species, A. bereziniae carries intrinsic oxacillinases, which are variants of OXA-228-like group able to confer carbapenem resistance in vitro. 21 In addition, A. bereziniae has been associated with IMP-1, IMP-19, IMP-5, SIM-1, VIM-2, and NDM-1 metallo-beta-lactamases, being able to harbor concomitantly even two different types of MBL.22–24
This study aims to describe the clinical and the microbiological findings of bloodstream infection (BSI) due to A. bereziniae in two patients from a teaching hospital in inner Brazil.
Materials and Methods
This study was approved as a retrospective study by the Local Research Ethics Committee (Process CAAE 49985115.5.0000.0059). We were granted an exemption from the requirement to obtain written informed consent from the participants and/or their legal guardians because the isolates included in the study had already been stored, on an ongoing basis, in the Culture Collection of the Department of Microbiology and Immunology, UNESP, Botucatu, São Paulo, Brazil.
As part of a retrospective study on the mechanisms leading to carbapenem resistance in Acinetobacter species from BSIs of patients treated at Botucatu Medical School Hospital/UNESP (BMSH/UNESP), a tertiary hospital in inner Brazil with 415 beds (52 intensive care unit [ICU] beds), during the period of 2007–2014, a total of 134 isolates from nonrepetitive individuals were screened for the main CHDL, serine, and metallo-carbapenemase genes.25–27 Of which, those that presented positive PCR results to blaIMP-like genes were further investigated. blaIMP allele was determined by Sanger sequencing. Partial rpoB gene and 16S-23S rRNA gene intergenic spacer (ITS) region sequencing28,29 was carried out to identify the species, since the isolates were negative to the blaOXA-51-like gene. ApaI-restricted genomic DNA fragments were resolved using pulsed-field gel electrophoresis 30 and the restriction patterns were analyzed using the software BioNumerics v. 7.6.2 (Applied Maths, Sint-Martens-Latem, Belgium). Presence of large plasmids was evaluated by using 10 U of S1-nuclease (Promega) detected with pulsed-field gel electrophoresis (PFGE) and the molecular weight Lambda PFG Marker (NewEngland Biolabs) to estimate their sizes. Small plasmids were extracted with PureYield™ Plasmid Miniprep System Protocol (Promega). Conjugation and transformation were attempted using the A. bereziniae strains as donors and Escherichia coli J53 and E. coli DH10β as recipient strains, respectively. In addition isolates of A. baumannii ATCC 19606, ATCC 13304 and ATCC 747 were also employed for conjugation experiments. Selection was made in MacConkey plates containing sodium azide (100 mg/L; Sigma Aldrich, St. Louis, only for E. coli) and imipenem (1 μg/mL; Sigma Aldrich) or cefotaxime (1, 2 μg/mL; Sigma Aldrich).
Minimum inhibitory concentration values were determined by using the Sensititre™ Gram Negative GNX3F AST Plate (ThermoFisher Scientific, MA) according to the fabricant recommendations. For polymyxin and colistin (Sigma, St. Louis, MO), the in-house broth microdilution method was carried out in untreated 96-well polystyrene microplates and cation-adjusted BBL Mueller–Hinton broth (BD, Franklin Lakes, NJ), since nonreliable results were found for Enterobacteriaceae by using the Sensititre approach. 31
Breakpoints employed to define susceptibility, intermediate or resistance followed Clinical and Laboratory Standards Institute recommendations 32 except for tigecycline, for which the U.S. Food and Drug Administration breakpoints were applied (susceptible: ≤2 μg/mL; resistant: ≥8 μg/mL).
Isolates were categorized as multidrug resistant (MDR, nonsusceptibility to ≥1 agent in ≥3 antimicrobial categories among aminoglycosides, antipseudomonal carbapenems, antipseudomonal fluoroquinolones, antipseudomonal penicillins + β-lactamase inhibitors, extended-spectrum cephalosporins, folate pathway inhibitors, penicillins + β-lactamase inhibitors, polymyxins, tetracyclines) or extensively drug-resistant (XDR, nonsusceptibility to ≥1 agent in all but ≤2 categories), as described elsewhere. 33
The IMP-producing isolates were subjected to whole-genome shotgun sequencing. Genomic DNA was extracted using the Promega Wizard Extraction Kit, according to the manufacturer's instructions and by adding 5 μL of proteinase K (20 mg/mL) and lysozyme (40 mg/mL) during the initial lysis step. DNA libraries were prepared at the Strategic Laboratory, Adolfo Lutz Institute, and sequenced using the Ion Torrent S5 (Life Technologies). The 520 Chip paired with a 200-bp library was chosen for maximum coverage. Raw reads were de novo assembled using SPAdes v.3.1. The FASTA files were submitted to online pipelines for bacterial identification on rMLST and Kmer tool, detection of antimicrobial resistance and virulence genes on CARD, ResFinder, and BactDB. Sequence types were determined by the Institute Pasteur MLST Scheme. Contigs carrying resistance genes were manually annotated using the Geneious v.9.1.9 (Biomatters) and CLC v.11.1 (QIAGEN). Images of genetic environment were done using the EasyFig software. 34
Using the raw reads of L98 and L65 strains, and other genomes of A. bereziniae deposited in GenBank database, a whole genome phylogenetic analysis based on the concatenated alignment of the high-quality single nucleotide polymorphisms (SNPs) was performed using CSI Phylogeny (version 1.4) with default input parameters. 35 A. bereziniae XH901 strain was used as reference genome. Sequences of L65 and L98 were deposited in GenBank under accession numbers SIRG01000000 and SIRF01000000, respectively.
Results
Out of the 134 carbapenem-resistant Acinetobacter spp. analyzed, 2 (1.3%) presented positivity for the blaIMP-like gene, further identified as blaIMP-1 by sequencing. We did not detect any other carbapenemase gene (SPM, IMP, VIM, KPC, NDM, and OXA-48) in the remaining 132 isolates. Those isolates were recovered from two different patients, as described hereunder.
Patients characterization
Patient 1, male, 62 years old, admitted to the emergency room after being ejected out his car after automobile collision with another vehicle. He did not present comorbidities. After the initial measures for treatment of traumatic injuries, patient remained hospitalized in the ICU of the emergency room (ICU-ER). On day 7 presented positive blood catheter culture for Acinetobacter sp. L65 (initially identified as A. lwoffi by Vitek 2, BioMérieux). Blood culture revealed also the presence of Pseudomonas sp. On the same day, meropenem therapy was initiated for 14 days. Tracheostomy was performed on day 14 following local protocol for intubated patients. On day 17 peripheral blood culture grew again Acinetobacter sp. and the therapy was changed to ampicillin-sulbactam; patient responded well. He remained hospitalized until total recovering of traumatic injuries by the day 45, when he was discharged presenting overall good condition and without clinical signs of infection and microbiological cultures testing negative.
Patient 2, female, 66 years old, history of hypertension, type 2 diabetes mellitus, and hypothyroidism. Admitted to the emergency room complaining of cough for 1 week and worsening of basal dyspnea for 3 days. Initial X-ray indicated increased cardiac area, with diffuse infiltrate and signs of venous congestion. Orotracheal intubation (OTI) was initiated because of low SatO2, but post-OTI shock was verified. The new X-ray after the OTI showed bilateral pulmonary congestion and cardiomegaly, suggesting decompensated congestive heart failure due to bronchial infection. Antimicrobial therapy was promptly initiated with levofloxacin 750 mg on day 16, same day in which two blood cultures were positive and the tracheal secretion quantitative cultured negative, but patient did not evolve well. On day 17 therapy was changed to vancomycin + piperacillin/tazobactam and patient presented worsening of overall conditions, with transfer to the ICU-ER. On day 30 presented a positive blood culture for Acinetobacter sp. L98, with MDR profile. Hemodialysis therapy was started on day 50 due to kidney failure. Patient remained hospitalized at ICU-ER and evolved to toxic-metabolic encephalopathy secondary to hypernatremia and sepsis, with multiple complications: respiratory failure, critical neuropathy, cardio respiratory arrest in electrical activity without pulse, thrombus pulmonary embolism treated with anticoagulation therapy, acute renal failure, fungal urinary tract infection, and anemia. On day 110 tracheal secretion was positive for imipenem-resistant Pseudomonas sp. (in counts >1 million colony forming U/mL), treated with polymyxin B, but patient evolved to death on day 128 due to multiple organ failure.
Isolates characterization
Partial rpoB and ITS region sequencing identified both isolates (L65 and L98) as A. bereziniae, and PFGE revealed similarity of 100% between them (Supplementary Fig. S1). Putative plasmids were detected in S1-PFGE (two plasmids of sizes ∼55 kb and <<20 kb) and in kit extraction (∼7 kb) (Supplementary Figs. S2 and S3). The blaIMP-1 gene was unsuccessfully transferred from the A. bereziniae to the A. baumannii ATCC 19606 strain by conjugation after several attempts.
According to the susceptibility tests, both A. bereziniae were considered MDR, presenting resistance to carbapenems, extended spectrum and antipseudomonad cephalosporins, amikacin, and trimethoprim-sulfamethoxazole. Although resistance to ciprofloxacin was not detected, both isolates presented intermediate susceptibility to this drug. Polymyxin B resistance was not detected.
Identification of both isolates was confirmed as A. bereziniae by the rMLST (100% support and 48 exact matches) and kmer (z-score 1007.3 for L65 and 1006.5 for L98) analysis on whole genome sequencing data. Additional genome sequencing data are presented in Supplementary Table S1. The same antimicrobial resistance genes were detected in both strains: blaIMP-1, blaOXA-356, and CARB-8 (resistance to beta-lactams), sul1 (resistance to sulfonamides), aph(3′)-Via, aac(6′)-31, and aadA1 (resistance to aminoglycosides), msr(E) and mph(D) (acquired resistance to macrolides). In addition, the efflux pump gene adeF was also detected.
Analysis of blaIMP-1 genetic context revealed its occurrence into a class I integron, In86, which also carried the sul1, aac(6′)-31, and the aadA1 genes (Fig. 1).
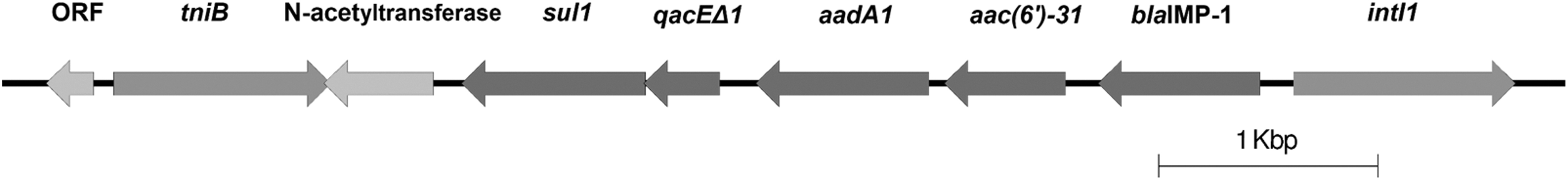
Schematic representation of contig harboring the In86 with the blaIMP-1. The total fragment length is 7,129 base pairs (bp), and the scale represents 1000 bp (1 Kbp).
Through genome assembly analysis, no circularized plasmids were found, only contigs containing coding regions for proteins present in plasmids and other mobile genetic elements.
After uploading our data on MLST (Pasteur Scheme) website, novel alleles were assigned for all the seven genes, and the novel sequence type 1309 was assigned to the alleles combination cpn60 (194); fusA (186); gltA (180); pyrG (91); recA (194); rplB (101); rpoB (169).
A phylogenetic tree built based on whole genome SNPs of L65, L98, and genomes of eight strains available in GenBank showed that the genomes of L65 and L68 were closely related (Fig. 2). Interesting, two branches grouped carbapenemase-producing strains from India, Brazil, and Argentina, as well as a noncarbapenemase-producing strain from the United States. An intermediate branch grouped L65 and L98 together.

The whole genome SNP-based phylogeny tree of 10 Acinetobacter bereziniae genomes available in GenBank. The legends correspond to strain identification (GenBank accession number) country, year of isolation, and carbapenemase reported. All genomes were selected from strains recovered of human infections. Branch labels indicate substitution per site (patristic distances). SNP, single nucleotide polymorphism.
According to the BACdb website the following putative virulence genes associated with A. baumannii species were detected in both the L65 and L98 A. bereziniae strains: ompA, which codifies an important outer membrane protein of A. baumannii and A. nosocomialis, associated with multiple virulence pathways such as inhibition of the alternative complement activation by binding factor H, biofilm formation on abiotic surfaces, and adherence to and invasion of host cells; bfmR, associated with regulation of two-component system; lpxC (associated with lipopolysaccharide production) and ABZJ_00085 and ABZJ_00086, putative IS4 family transposases open read frame.
Discussion
In this study, we identified two MDR carbapenem-resistant IMP-1-producing A. bereziniae strains isolated from BSIs of patients with bacteremia attending in a teaching hospital in Brazil. A. baumannii is considered a major threat in health care-associated infections, but NBA species have emerged as important opportunistic agents carrying and spreading antimicrobial resistance determinants. 5
Infections caused by A. bereziniae are rarely reported and this species is not frequently isolated. Nevertheless, it has already been identified in the nosocomial environment worldwide, including in Brazil, where studies had shown that this species may be the reservoir of blaIMP-1 and blaNDM-1 genes6,36 or even carry more than one MBL gene. 23 Our results correlate to this occurrence, since both strains carried blaIMP-1, which was inserted into In86, as first described in Acinetobacter species and Pseudomonas putida from the 2000s. 37 This same genetic context was found by Mendes et al. (2007) in a A. baumannii strain isolated from infected skin and soft tissue in São Paulo, Brazil (GenBank accession no. AJ640197). More recently, In86 was also detected in A. bereziniae from a blood and tracheal secretion of two distinct patients attending at two distinct hospitals in the city of São Paulo, Brazil. 6 Conjugation experiments failed to prove the occurrence of plasmid-borne blaIMP-1, and hybridization experiments targeting the blaIMP-1 were not conclusive as well (data not shown). Therefore, with this evidence, we cannot assume that In86 is allocated into a plasmid. Long reads sequencing could answer this limitation of our study, but it was not performed.
Contrary to the clonal diversity reported by Cayô et al., maybe explained by the source of the isolates, we verified that both A. bereziniae strains of our study shared the same PFGE pattern and only a few SNPs, even though the samples were recovered from blood of two different patients more than 1 year apart (2012–2013). Such finding underscores the persistence of A. bereziniae in this ward and its potential as a pathogenic bacterium since it was isolated from BSIs in at-risk ICU patients.
To the best of our knowledge, this is the first study analyzing the resistance and virulence determinants by whole genome sequencing (WGS) of IMP-producing A. bereziniae isolates and contribute the knowledge of MBL among such species, since blaNDM-1 was already reported in isolates from a neighbor country, Argentina.38,39 WGS has permitted the total analysis of the bacterial genome, allowing the identification of resistance and virulence genes.
In the A. bereziniae isolates, several resistance genes were detected, in accordance with the MDR pattern detected by the phenotypic methods. The OXA-228 group, which includes the OXA-228, OXA-229, OXA-355, and OXA-356 enzymes, is the intrinsic OXA group in this species. Thus, L65 and L98 A. bereziniae carried the expected intrinsic OXA-enzyme.21,40
Compared with the available A. bereziniae genomes by SNPs analysis, the L65 and L98 strains were closely related to each other, but were not grouped with carbapenemase-producing strains from Brazil, Argentina, or India, or with the other genomes used in our analysis. There are few genomes of this species deposited in public databases and very little information about genomic diversity of A. bereziniae. Our phylogenetic analysis showed point differences among the analyzed genomes, suggesting that this species can present similar genetic features among lineages circulating worldwide. This evidence can be illustrated by the fact that other genomes of A. bereziniae from Brazil (374/14 strain, not published) used in our analysis was grouped with a strain from the United States, and not with L65 and L98.
Indeed, by using the A. baumannii database, we identified five putative virulence genes in both A. bereziniae isolates, at least one (ompA) with recognized function associated with adherence and invasion, biofilm formation and persistence on abiotic surfaces, highlighting the pathogenic potential of these strains. Adequate antimicrobial therapy is, therefore, mandatory to improve patients' survival. In our description, one of the two cases (L98) evolved to death, but a lot of comorbidities were also reported.
Conclusion
This study reinforces the importance of identifying NBA, since despite the low frequency of infections caused by A. bereziniae, the two strains reported in this study were isolated from invasive infections. Moreover, both strains were considered MDR and harbored the blaIMP-1 gene embedded into In86 (flanked by Int1 and tniB). Such finding represents a reservoir of antimicrobial resistance genes that can further disseminate to other strains or even species, impairing successful antimicrobial therapy in severe infections.
Footnotes
Acknowledgments
We thank the team of curators of the Institut Pasteur Acinetobacter MLST system for curating the data and making them publicly available, and especially the kind assistance of Dr. Raffaele Zarrilli.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the grant nos. 2015/13179-4, 2016/20727-0, and 2017/16988-6, Fundação de Amparo à Pesquisa do Estado de São Paulo. C.H.C. has received Productivity Research Fellows from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.