
Abstract
IncF ColV plasmids are important plasmid incompatibility group that are currently restricted to the Enterobacteriaceae. These plasmids carry an important repertoire of virulence-associated genes (VAGs) that contribute to the ability of avian pathogenic Escherichia coli to cause disease in poultry. VAGs found on ColV plasmids have also been linked to urosepsis and meningitis in humans but the mechanisms that elicit these disease conditions are not well understood. Recently we described the sequence of a ColV plasmid pSDJ2009-52F that carried the typical repertoire of VAGs and a complex resistance gene locus flanked by IS26, an insertion element that plays an important role in mobilizing antibiotic resistance genes on plasmids and genomic islands. We recovered complete ColV-like plasmid sequences from public databases that shared >80% sequence identity with pSDJ2009-52F in geographically diverse regions of the world over a 20-year timeframe. Previously we noted that pSDJ2009-52F carries a unique genetic signature in the class 1 integron within the complex resistance locus that was presumably created by the action of IS26. Here we show that most ColV-like plasmids that are closely related to pSDJ2009-52F also carry the same signature. Our studies provide insight into how these signature-bearing plasmids and the mobile genetic elements they carry traffic between E. coli sequence types over large geographic distances.
Introduction
E
Plasmids are important vehicles for capturing and assembling antibiotic resistance, heavy metal resistance, and virulence genes2–5 and their presence can impact how pathogens evolve.2,6–8 Numerous plasmid incompatibility types circulate in human-sourced ExPEC, but IncF plasmids play a key role in housing genes conferring resistance to last line antibiotics, including the New Delhi metallo-β-lactamase gene blaNDM-1, 9 the cephalosporinase gene blaCTX-M, 10 and various virulence genes. 8 ColV plasmids often carry an extensive suite of virulence determinants, including, but not limited to, increased serum survival (iss) and aerobactin biosynthesis (iuc operon) and other iron acquisition genes. 11 IncF ColV plasmids are widespread among APEC strains and can be found in commensal human flora 12 and in E. coli causing urosepsis8,13,14 and neonatal meningitis.15,16
IS26 is an insertion element that plays a major role in the capture and assembly of multiple antibiotic resistance genes in complex resistance gene regions found on diverse plasmid backbones.3,17–19 IS26 has no recognized target site specificity, 18 nonetheless insertions next to class 1 are reported frequently20,21 possibly because they are unlikely to infer a significant fitness cost and afford the host bacterium more than one mechanism to acquire antibiotic resistance genes. IS26 often creates random deletions in DNA that abut its insertion site. We have previously investigated IS26-associated deletion events as unique genetic signatures for the purpose of tracking genetic elements that purvey multiple drug resistance.4,20,22 Collectively these and more recent genomic surveillance studies have shown that the conserved structure of class 1 integrons is frequently altered by the action of insertion elements, particularly IS26.8,21–24 One such class 1 integron structure contains the trimethoprim resistance gene cassette dfrA5 and only 24 bp of the 3′-conserved segment (CS). It has been reported in association with multiple drug-resistant E. coli from the feces of Australian production animals4,20,21 and in human patients with hemorrhagic colitis 3 and urosepsis 8 and is often carried on Tn3 family, mercury resistance transposons.
Recently, we characterized a ColV-like plasmid, pSDJ2009-52F, in a multiple drug-resistant E. coli with ST58 from a patient with urosepsis. Although ST58 E. coli are found in healthy and diseased agriculturally important animals24–26 and flies, 27 the environment, 28 and wildlife, 29 it is not noted to be a frequent cause of human disease. 8 Plasmid acquisition is known to play a critical role in pathogen evolution.2,6,7 Carriage of pSDJ2009-52F may have played an important role in the ability of ST58 strain 2009-49 (urine isolate) to cause urinary tract infection followed by urosepsis 8 (2009-52). Here we searched public databases for plasmids closely related to pSDJ2009-52F. We identified IncF ColV plasmids similar to pSDJ2009-52F from different countries, years, and hosts and from E. coli with diverse sequence types (STs) and investigated their phylogeny.
Materials and Methods
GenBank reference to plasmid pSDJ2009-52F
We have recently described the isolation, sequencing, and characterization of plasmid pSDJ2009-52F (GenBank accession no. MH195200.1) from E. coli strain 2009-52 (GenBank accession no. SAMN07661348). 8
Single nucleotide polymorphism analysis
Parsnp from the Harvest Tools suite 30 was used to identify single nucleotide polymorphisms (SNPs) between pSDJ2009-52 and the five most closely related plasmids in GenBank as determined by BLASTn analysis. The x flag was implemented to filter out recombination events, resulting in an alignment of 62% of the plasmid backbones. Gingr, also from Harvest Tools, was used to visualize the SNP locations in the .ggr file and the matching coordinates confirmed from the exported .vcf file.
BEAST2 analysis
To identify plasmid evolution over time, we used a Bayesian Markov chain Monte Carlo method implemented in the BEAST2 package. 31 The Generalized Time Reversible (GTR)-I substitution model was used as the plasmids were very closely related. A strict molecular clock on a constant population size coalescent model was assumed and analyzed as reported previously. 32
Results
Plasmid information
Plasmids with >95% sequence identity over 80% of the length of pSDJ2009-52F, and including the repA linked virulence genes characteristic of IncF-ColV plasmids, were downloaded from GenBank. Twelve plasmids with high sequence identity (>80%) to pSDJ2009-52F are given in Table 1 along with a description of their salient features. Of note, 8 of 11 plasmids were isolated from E. coli sourced from humans including strains from patients with gastrointestinal disease and strains from apparently healthy humans (plasmids cited as site: feces excluding strain 2013C-4390). These plasmids are found in E. coli with eight STs from geographically diverse regions around the globe and multiple sources including humans, chicken, canine, swine, and turkey. The strains carrying these related plasmids were sourced over a 22-year period with 1990 being the earliest isolation date. Besides pAPEC-O78-ColV (IncF24) all others were plasmid MLST F2:A-:B1.
Plasmids with High Sequence Identity to pSDJ2009-52F
Cells are blank if information was not provided or could not be ascertained. Plasmid pSDJ2009-52F 8 is identified in bold.
ST, sequence type.
Virulence and antibiotic resistance gene carriage
A full set of genes typical of ColV plasmids 11 were observed in all but two plasmids, p2013-4390 and 134q. These include the iro operon (iron-related), iuc operon for aerobactin biosynthesis, ferric aerobactin receptor iutA, sit operon for iron/manganese transport, cvaABC/cvi for colicin V secretion and immunity, putative type 1 secretion system genes etsABC, avian hemolysin hlyF, outer membrane protease ompT, and conjugal transfer exclusion protein traT and iss for increased serum survival and complement resistance. Plasmid 134q was missing the etsABC genes and hlyF and ompT, and plasmid “unnamed” from strain 2013C-4390 were missing the iutA gene and the iuc operon. Because of the presence of an IS1 immediately after the sit operon in this plasmid, the normally adjacent iuc operon is likely the target of an IS1-mediated deletion. The remaining nine plasmids contained the full set of standard IncF ColV virulence genes present in pSDJ2009-52F, although with occasional SNPs (Table 2).
Percentage Sequence Identity Across Whole Open Reading Frame Based on BLASTn Analyses to Virulence Genes Found on IncF colV Plasmids
Representative genes have been used as markers for larger operons (e.g., iroN, iucD, and sitD). Italicized plasmids were those not used in comparative analyses. Blank cells indicate gene absence. The plasmid in bold indicates query sequence for all BLAST searches (and not one that came up in the search result).
As expected, antibiotic resistance genes were less conserved across the plasmids with only one plasmid, pECAZ147_1, exhibiting an identical profile as pSDJ2009-52F. Nine of 11 plasmids contained a blaTEM-1 ampicillin resistance gene variant, 8 of 11 contained the sul2-strA-strB sulfonamide and streptomycin resistance gene cluster, 9 of 11 contained trimethoprim resistance gene dfrA5, and 6 of 11 contained tetracycline resistance gene tetA, whereas sulfonamide resistance gene sul1 and kanamycin resistance gene aphA1 were singularly represented. dfrA5 is a resistance gene cassette typically linked with carriage of a class 1 integron. As such the low carriage rate of sul1 is unusual given that sulfonamide resistance gene is typically resident in the 3′-CS of class 1 integrons.
Comparative SNP analyses
Plasmids that shared at least 90% sequence identity over the full length of plasmid pSDJ2009-52F, specifically pECAZ147_1, plasmid “unnamed” from strain 2013C-4390, pG749_1, pDB4277, pSF-088-1, and pCERC4, were examined further. The high similarity of the plasmid backbones is evident in Fig. 1. As described previously, the major differences among these plasmids reside in the assembly of the complex resistance locus (CRL) resident in each plasmid. pSDJ2009-52F contains an atypical class 1 integron containing a dfrA5 trimethoprim resistance cassette bearing only 24 bp of the 3′-CS carried on a hybrid Tn1721/Tn21 transposon (Supplementary Fig. S1A). A copy of the composite transposon Tn6029 that harbors the blaTEM-1 gene and sul2-strA-strB gene cluster encoding resistance to ampicillin, streptomycin, and sulfonamides abuts the truncated 3′-CS. The resistance gene locus is essentially identical with the locus found on EHEC plasmid pO26-CRL111 sourced from a case of hemorrhagic colitis in 19983,4; however, the left arm of the transposon lacks the aphA1 gene of Tn6026 (Tn6029) and the mercury resistance transposon on pO26-CRL111 lacks a copy of Tn1721. pECAZ147_1 and “unnamed” (strain 2013C-4390) contain an identical resistance region comprising the class 1 integron, hybrid Tn21/1721 transposon, and Tn6029 to pSDJ2009-52F, whereas pG749_1, pDB4277, and pSF-088-1 are missing the leftmost arm of Tn1721 including the tetracycline resistance genes tetA and tetR. pCERC4 was isolated from an E. coli from a healthy human living in Sydney and lacks Tn6029 (Supplementary Fig. S1B).
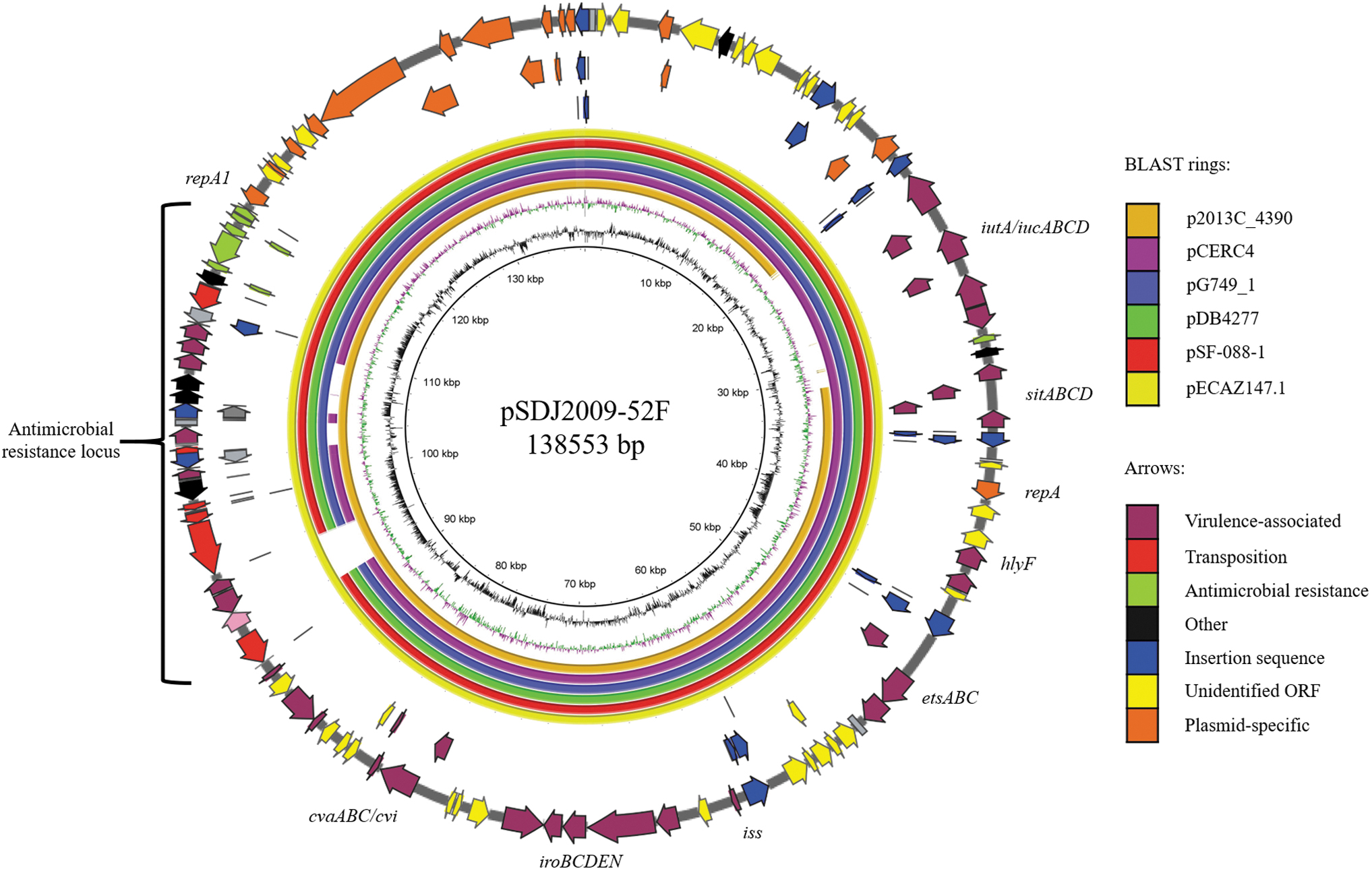
BLAST comparison, produced by BRIG, 33 of plasmids in GenBank matching at least 90% sequence identity to pSDJ2009-52F via BLAST analysis. Outside ring (arrows) shows gene content of pSDJ2009-52F along three different reading frames that correlate with regions in the plasmids that have come up in the BLAST analysis. Plasmid “unnamed” has been renamed according to host strain 2013C-4390.
Parsnp identified 20 unique SNPs across all plasmid backbones compared with pSDJ2009-52 (62% sequence coverage shared across all plasmids), 2 of which were shared across all 6 plasmids. These two SNPs were located in the putative type 1 secretion system gene etsA gene and conjugal transfer gene traD. Neither of these SNPs were predicted to affect protein function using the PROVEAN web program. 34 pECAZ147_1 contained two additional SNPs in traD; however, these were also tolerated. The closest relative to pSDJ2009-52F was pDB4277. It differed from pSDJ2009-52F only by the two SNPs described previously. pDB4277 was isolated from a UPEC strain of unknown ST in the United Kingdom in 2004. The unnamed plasmid from Shiga toxin-producing E. coli strain 2013C-4390 also only contained these two SNPs and an identical atypical In22 class 1 integron and hybrid Tn1721/Tn21 transposon to pSDJ2009-52F but lacked the iutA gene and iuc operon. pCERC4 and pG749_1 were the next most closely related with four SNPs each, two of which were unique in both cases. These were located in an uncharacterized open reading frame and the relaxase gene traI involved in plasmid conjugation for pCERC4, and mercury resistance protein merP and traI for pG479_1. Plasmid pECAZ147_1 from an ST540 E. coli isolated in Israel in 2012 housed an identical complex antimicrobial resistance region to pSDJ2009-52F but contained four unique SNPs. These were in conjugative transfer proteins traD (two SNPs) and traG, and an uncharacterized open reading frame. The most disparate backbone sequence was pSF-088-1, with 10 unique SNPs, 8 of which were within mobile elements IS1 and IS2. The remaining two were within uncharacterized open reading frames.
BEAST2 analysis
Time-measured phylogeny (Supplementary Fig. S2) estimated the oldest common ancestor from which these plasmids have evolved is 4.6 thousand years. The closest evolutionary relation to pSDJ2009-52F is plasmid “unnamed” from strain 2013C-4390 isolated from a Shiga toxin-producing E. coli.
Molecular signatures
Plasmid pSDJ2009-52F houses a characteristic IS26-mediated deletion in the class 1 integron and retains only 24 bp of the 3′-CS. Of note, most plasmids shown in Table 1 carried this signature, and five of these fell above the 90% query cover cutoff (Fig. 1).
Discussion
IncF plasmids are a prominent incompatibility group among E. coli from human and animal sources. 35 Their dominance is probably because of their highly conjugative nature in conjunction with their host range being limited to Enterobacteriaceae. Consistent with this view we observed plasmids that share high sequence identity with pSDJ2009-52F in E. coli with diverse STs (Table 1). Of the 11 strains, 8 unique STs were represented including the multiresistant clonal group ST131 (pG749_1: H22, pCERC5: undisclosed), well documented for causing extended spectrum beta-lactamase (ESBL)-producing and fluoroquinolone-resistant extraintestinal infection. 36 IncF plasmids described here not only contain extensive virulence gene cargo associated with extraintestinal disease, but 9 of 11 plasmids contain a class 1 integron, a reliable proxy for multiple drug resistance.37,38 Consistent with this, all nine plasmids carry numerous antibiotic resistance genes. IncF type plasmids are inherently conjugative if the tra locus is intact. The observation that diverse E. coli carry MDR IncF ColV plasmids is a cause for concern, as these strains tend to fly under the radar because of the current research bias toward those that have established associations with genes conferring resistance to extended-spectrum β lactams. 35
Two plasmids in this study, pCERC4 and pSF-088-1, were isolated from strains of E. coli ST95, a pandemic lineage that has not yet received the “elite” global status of ST131 because of the relatively low frequency of multiresistant strains. 39 However, ST95 is responsible for a significant number of human ExPEC infections and systemic disease in poultry.40–43 One study 43 found that a cohort of 116 ST95 strains form biofilms, resist host serum bactericidal activity, and adhere to and subsequently invade mammalian kidney cell lines. They found no distinguishable differences between strains of human and avian origin in terms of virulence attributes and pathogenesis. This poses a serious risk in terms of human clinical infection as ST95 strains are generally virulent and recognized human pathogens. 39 Evidence linking APEC with episodes of human disease is mounting.42,44–47 Carriage of ColV plasmids likely plays a key role as IncF ColV plasmids are typically associated with APEC. 11 The report of isolation of MDR ColV plasmid pCERC4 from commensal fecal E. coli of a healthy human that had not received antibiotics for at least 6 months 48 shows that plasmids carrying important virulence gene cargo reside in the flora of healthy humans and pose a potential threat for urinary tract infection (UTI).12,49
The highly related plasmids represented here have been isolated from three animal sources and humans and were derived from diverse geographic locations (Table 1). The similarity in virulence gene content and the minor variations in complex antimicrobial resistance loci among the IncF plasmids described here highlight the global dissemination of the IncF incompatibility group and their propensity to house pathogenicity determinants. ColV plasmids are important in this regard because they are resident in APEC that cause disease in poultry, carry a significant armory of virulence-associated genes associated with human and food animal disease, and acquire antimicrobial resistance creating strains capable of high-risk infection in a single genetic transfer event.7,8 Evident in the analysis described here is the propensity of ColV plasmids to acquire CRL. Numerous mobile elements are involved in the assembly of these CRL and they may undergo rearrangement and recombination events in the host in response to selective pressures exerted by the niche they inhabit. Time-measured phylogeny (Supplementary Fig. S2) estimated the oldest common ancestor from which the plasmids in this study have evolved is 4.6 thousand years. Antimicrobial selective pressures in recent times have likely played a role in the formation and rearrangement of CRL described in this plasmid cohort, but natural selection of the ColV plasmid backbones predates the modern antibiotic era. Therefore it is possible that this backbone has, in part, become established because of the beneficial cargo it carries that includes efflux proteins, colicin/colicin immunity, heavy metal resistance, and increased serum survival.
The observation of this plasmid backbone in several animal species and human commensal and pathogenic bacteria is cause for concern. Our studies underscore the need for a “One Health” approach to understand the real impact of reliance on antimicrobials to control infectious disease and to produce food for a burgeoning world population.
Footnotes
Authors' Contributions
J.M.: Conceptualization, data curation, formal analysis, investigation, visualization, and writing—original draft (lead). P.R.C.: Investigation, resources, supervision, visualization, and writing—review and editing. S.P.D.: Conceptualization, funding acquisition, project administration, supervision (lead), writing—original draft, and writing—review and editing.
Disclosure Statement
No conflicts of interest to declare.
Funding Information
This study was supported by Ausgem, a collaborative partnership between the New South Wales Department of Primary Industries' Elizabeth Macarthur Agricultural Institute (EMAI) and the ithree Institute at the University of Technology Sydney (UTS). J.M. is the recipient of an Australian Government Research Training Program Scholarship. These results are helping to inform the OUTBREAK decision support system funded by the Medical Research Future Fund Frontier Health and Medical Research Program.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.