
Abstract
Although the human intestinal microbiome has been shown to harbor antimicrobial drug resistance genes (ARGs), the prevalence of such genes in a healthy population and their impact on extraintestinal infections that occur in that community are not well established. This study sought to identify ARG prevalence and their mobile elements in the intestines of a healthy community population at a California University, and compared these genes to those previously identified among uropathogenic Escherichia coli isolated from patients with urinary tract infection from the same community. We isolated Gram-negative bacteria (GNB) from fecal samples of healthy volunteers and screened them by polymerase chain reaction for class 1 integron cassette sequences and ARGs encoding resistance against ampicillin, trimethoprim-sulfamethoxazole, gentamicin, and colistin. We found antimicrobial-resistant GNB from 83 (81%) of 102 nonredundant rectal swab samples. Seventy-four (72%) of these samples contained β-lactamase genes (blaTEM, blaSHV, blaCTX-M, blaOXA, and blaOXY), dihydrofolate reductase (DHFR) genes (dhfr-A17, dhfr-A12, dhfr-A7, dhfr-A5, dhfr-A21, dhfr-A1, dhfr-A13, and dhfr-7), and aminoglycoside resistance genes (aadA5, aadA2, aadA1, and aadB). Integron sequences were found in 37 (36%) fecal samples. These genes were found in 11 different GNB species. The high prevalence of clinically common ARGs and integrons harbored by GNB in the intestine of a healthy population suggest that human intestines may serve as a major reservoir of these mobile ARGs that appear in E. coli strains causing extraintestinal infections in the same community.
Introduction
The emergence of antibiotic-resistant bacteria and antibiotic resistance genes (ARGs) has become a growing public health concern worldwide. Gram-negative bacteria (GNB), including Enterobacteriaceae and glucose nonfermenter species, are implicated in a large number of community- and health care-associated infections. They are also common residents of the human intestinal microbiota as well as of the environment.1–10 Human and veterinary medicines rely on the use of antibiotics to treat infections caused by bacterial pathogens. Antibiotics are also used in food animal production as growth promoters. Extensive use of antibiotics over the last several decades has led to the rapid emergence and spread of resistant microbes, especially among enteric bacteria.11,12 Due to the nature and complexity of the microbial communities harbored in our intestinal environment, the human gut has been recognized as a potential reservoir of ARGs.2,3,13 However, ARGs are also found in food animal intestines as well as in a variety of produce items humans eat.2,12,14–30 Thus, food may serve as sources of ARGs that enter the human intestine. The prevalence of ARGs in healthy human intestine and their impact on human infections are not well characterized.
The ability of bacteria to disseminate mobile ARGs by horizontal gene transfer (HGT) has resulted in the rapid acquisition of resistance by enteric bacterial pathogens. ARG mobile elements include integrons, transposons, and plasmids. Integrons, which permit the simultaneous integration of multiple exogenous gene cassettes, represent an important vehicle for the rapid horizontal transfer of resistance across bacterial populations and thus could contribute to the sudden increase in prevalence of multidrug-resistant infections in a community.31,32 In particular, class 1 integrons are the most ubiquitous class of integrons in enteric bacteria and have been found in all common pathogens, including Escherichia, Klebsiella, Salmonella, Shigella, and other disease-causing Enterobacteriaceae. More than 70 different gene cassettes conferring resistance to most of the known β-lactam drugs, aminoglycosides, trimethoprim, rifampicin, chloramphenicol, and erythromycin have been reported in class I integrons. 11 This is of further concern due to the increasing prevalence of extended-spectrum beta lactamase (ESBL)-producing GNB, which are difficult to treat and often resistant to other families of antibiotics, in particular trimethoprim-sulfamethoxazole and fluoroquinolones. 7 Furthermore, the recent global spread of GNB containing plasmid-mediated colistin resistance has greatly challenged clinical management of infections caused by them. 17 For example, colistin remains an important therapeutic agent for infections caused by carbapenem-resistant Enterobacteriaceae, in which there are limited alternative treatment options. 18
Previous studies have shown food and the environment contain ARGs found in pathogenic bacteria.12,19,20 Environmental reservoirs of ARG-containing pathogens include lakes and rivers, 21 wastewater treatment plants,22,23 houseflies, 24 livestock,12,25,26 soil and manure,27,28 retail meat products,12,28,29,30 companion animals,5,6,8,25,33 alfalfa sprouts, 34 retail spinach, 14 and other vegetables. 35 The findings from these studies indicate that human health may be impacted by uptake of pathogens from the environment, many of which harbor ARGs on mobile genetic elements. Thus, humans may be acquiring pathogens as well as transmissible ARGs through food and other environmental hosts.
A recent study by Yamaji et al. compared ARGs and genotypes of Escherichia coli isolates from patients with urinary tract infection (UTI) in a California university community to those of E. coli isolated from meat (pork, chicken, beef, and turkey) obtained from retail stores in the surrounding area. 28 They found that despite 12 shared genotypes (sequence types) between humans and retail meat E. coli isolates, human isolates contained more ARGs than meat isolates. 28 These findings led us to speculate that people may acquire uropathogenic E. coli from external sources (food or environment), but that these E. coli strains may acquire ARGs from commensal bacteria already in the gut. Therefore, we sought to understand the diversity and abundance of mobile ARGs in the gut of healthy humans, and to elucidate potential links between ARGs harbored in commensal gut bacteria and the acquisition of those ARGs by pathogenic GNB.
Materials and Methods
Sample collection
We prospectively cultured fecal samples from 113 healthy volunteers at a university campus in northern California between June and October 2017. Eligible participants included those between 18 and 65 years of age, with no medical history of urinary tract corrective surgery or abnormality, or bladder catheterization or hospitalization within the 6 months before sample collection.
At recruitment, participants were provided a preaddressed collection kit containing a Cary-Blair transport media rectal swab (Becton Dickinson BBL™), two biosafety bags, and detailed collection instructions. Each kit also included a questionnaire regarding antibiotic use, history of UTI, and diet and lifestyle characteristics. Participants were instructed to send the rectal swab back to the laboratory by United States Postal Service mail immediately after collection. Once delivered, the study coordinator analyzed samples within 48 hours.
Fecal sample analysis
Fecal swab tips were placed in a 1.5-mL Eppendorf tube containing 1 mL of Luria-Bertani broth and vortexed for 60 seconds. A 10 μL aliquot of fresh fecal material was dilution streaked onto MacConkey agar plates and incubated overnight at 37°C. All samples were screened for resistance against four antimicrobial agents: ampicillin (AMP) (32 μg/mL), gentamicin (GENT) (16 μg/mL), trimethoprim-sulfamethoxazole (TMP-SMX) (4–76 μg/mL), and colistin (COL) (2 μg/mL), as well as on one MacConkey agar plate containing no drug. All antimicrobial agents were dissolved in nuclease-free water and filter sterilized. Dimethyl sulfoxide (DMSO) was used to prepare TMP-SMX solution (final concentration <5% DMSO). Dihydrofolate reductase (DHFR) mediates trimethoprim resistance. We tested trimethoprim-sulfamethoxazole in this study as this combination drug is commonly used to treat UTI and other bacterial infections. Interpretive criteria from the Clinical Laboratory Standard Institute 36 or literature recommendations 37 were used to determine resistance. E. coli 25922 (ATCC) was used as a reference strain.
DNA extraction and ARG identification
Five bacterial colonies were randomly selected from each plate. If multiple colony morphologies were noted, all colonies with unique morphologies were sampled. If less than five colonies were present, all colonies were selected for analysis. Single colonies were selected and inoculated into 3 mL tryptic soy broth and incubated in a shaking incubator for 20 hours at 37°C. Basic procedures for DNA extraction by a freeze-thaw method were performed as previously described.14,31 The 2 mL aliquots of the cultures were centrifuged, and the pellets were resuspended in a test tube with 350 μL of distilled water, boiled for 10 minutes in a water bath, and then cooled on ice for 2 minutes. The samples were centrifuged for 2 minutes at 13,000 rpm and the supernatants were stored at 20°C before they were subjected to polymerase chain reaction (PCR) tests.
Two microliters of the resulting supernatant was used as template DNA in 25 μL of PCR mixture. Bacterial isolates that grew in the presence of AMP were examined for β-lactamase gene families by multiplex PCR as described previously.
38
These β-lactamase gene families included the following: TEM variants (blaTEM-1 and blaTEM-2), an SHV variant (blaSHV-1), CTX-M (all variants), OXA variants (blaOXA-1, blaOXA-4, and blaOXA-30), and KPC variants (blaKPC-1 to blaKPC-5). Samples that showed bands corresponding to CTX-M universal variants were further examined for CTX-M group variants: CTX-M group 1 (blaCTX-M-1, blaCTX-M-3, and blaCTX-M-15), CTX-M group 2 (blaCTX-M-2), CTX-M group 9 (blaCTX-M-9 and blaCTX-M-14), and CTX-M group 8/25 (blaCTX-M-8, blaCTX-M-25, blaCTX-M-26, and blaCTX-M-39 to blaCTX-M-41). Isolates that tested positive for CTX-M groups were sequenced to determine variant type. Bacteria that grew on plates containing GENT or TMP-SMX were examined for 5′ and 3′ conserved sequences flanking the class I integron gene cassettes. Thus, the entire cassette sequences harbored in the integron were analyzed. If present, these gene cassettes were sequenced to detect the presence of aad and dhfr gene types for GENT and TMP-SMX, respectively. Bacterial colonies isolated from colistin-containing plates were examined for mcr-1-5 genes by multiplex PCR as previously described.
39
All isolates were speciated by 16S ribosomal RNA (rRNA) sequencing. Species diversity was calculated by the Shannon diversity index:
where:
H = the Shannon diversity index
Pi = fraction of the entire population made up of species i
S = numbers of species encountered
∑ = sum from species 1 to species S
Primers used in these procedures are noted in Supplementary Table S1. PCR products were visualized on a 1.5% agarose gel stained with SYBRsafe DNA gel stain (Invitrogen) and visualized under UV transillumination.
Sequencing analysis
Each PCR product that showed an electrophoretic band of an expected molecular weight was cleansed and submitted for direct sequencing. Sanger sequencing was performed on a DNA analyzer (Applied Biosystems 3730 DNA analyzer) at the University of California, Berkeley, DNA Sequencing Facility. Isolates showing bands for CTX-M group families were sequenced to determine variant type. All nucleic acid sequences were compared against those deposited in the National Center for Biotechnology Information database using the BLAST program to search for regions of high similarity. Nucleic acid sequences of the class 1 gene cassette amplicon were compared against previously deposited dhfr and aad gene sequences in the NCBI Reference Gene Catalog (Bacterial Antimicrobial Resistance Reference Gene Database, version 2019-09-06.1). Sequences were also compared against available gene sequences for blaCMY, blaNDM, blaGES, blaVIM blaIMP, blaOXA, blaCTX-M, blaDHA, sul1, and mcr1-mcr9. Genes were considered to be present if reference sequences were ≥98% identical with ≥80% base pair alignment.
Isolates genus and species were identified through the amplification of the 16S rRNA gene. Species were determined by the criteria of >98% sequence identity and genus by >95% sequence identity. For amplicons over 1,000 base pairs in length, bidirectional sequencing was performed.
Results
Between June 2017 and February 2018, we obtained 113 unique rectal swab samples. Of these, 11 exhibited no growth on a MacConkey control plate, and were not further analyzed. Of the 102 remaining samples, 78 (76%) contained GNB that exhibited resistance to AMP, 14 (14%) to GENT, 47 (46%) to TMP-SMX, and 10 (10%) to COL. Only 19 (19%) samples contained isolates that were sensitive to all drugs tested (Fig. 1). One AMP-resistant sample, one TMP-SMX-resistant sample, and one GENT-resistant sample were not able to be cultured in broth, and thus were not considered for further analysis.
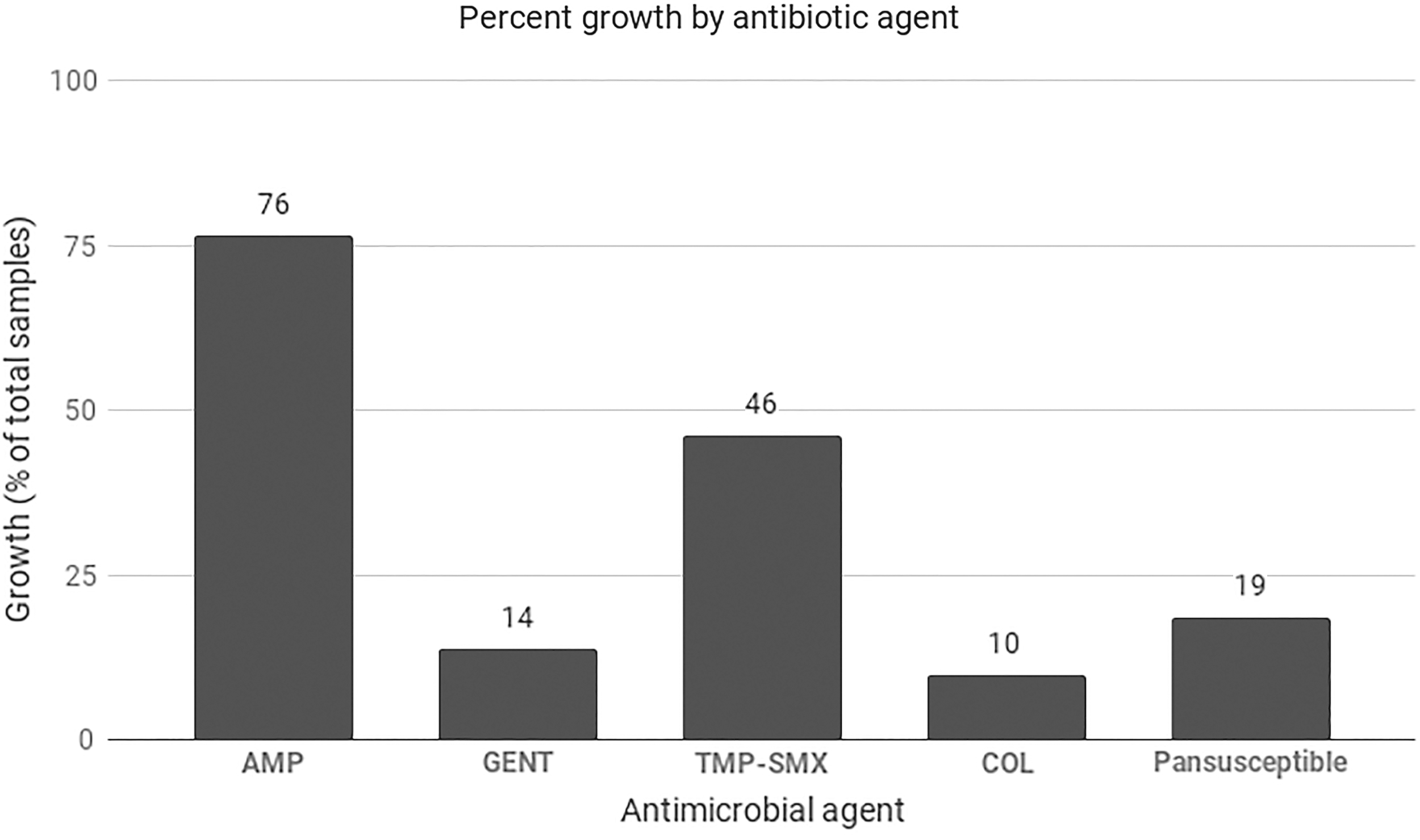
Bacterial growth (%) on MacConkey plates containing antimicrobial agents. Pansusceptible isolates were defined as those that exhibited growth on MacConkey control plate, but no growth in the presence of each of four antimicrobial agents tested.
Bacterial growth on at least one drug-containing plate was observed in 83 (81%) of 102 samples. Resistance to only 1 antimicrobial agent was observed in 31 (30%) of samples, 40 (39%) were resistant to 2 antimicrobial agents, and 12 (12%) were resistant to 3 or more agents (Table 1).
Colony Growth Was Observed in 83 Fecal Samples on MacConkey Plates Containing Distinct Types of Antimicrobial Agents
The number of types of antimicrobial agents to which a sample contained bacterial isolates that displayed resistance.
AMP-resistant, ampicillin resistant; COL-resistant, colistin resistant; GENT-resistant, gentamicin resistant; TMP/SMX-resistant, trimethoprim-sulfamethoxazole resistant.
Detection of β-lactamase genes
Three hundred sixty-five colonies obtained from 77 fecal samples containing AMP-resistant isolates were analyzed for the presence of β-lactamase genes. Of the 77 fecal samples, 68 (88%) contained at least one β-lactamase gene. Among the 102 total fecal samples, TEM variants (blaTEM-1 and blaTEM-116) were detected in 57 (55%); 19 (18%) contained an SHV variant, 1 (1%) contained an OXA variant (blaOXA-1), and 15 (15%) contained at least one CTX-M variant. Upon further analysis of CTX-M subgroups, 6 (6%) had CTX-M group 1 (i.e., blaCTX-M-15), 2 (2%) had CTX-M group 2, and 4 (4%) contained CTX-M group 9 variants (i.e., blaCTX-M-14, blaCTX-M-27). Two (2%) contained more than one CTX-M group variant. Six samples contained a CTX-M gene that did not belong to any of the four CTX-M family members tested. Sequence analysis of these PCR products detected an OXY variant sequence in four of these samples.40,41 Twenty-five isolates from nine fecal samples contained confirmed CTX-M variants. No KPC or CTX-M group 8/25 gene types were detected in this study.
Detection of dihydrofolate reductase and aminoglycoside adenyltransferase genes contained in class 1 integron gene cassettes
Two hundred twenty-four TMP-SMX-resistant colonies and 54 GENT-resistant colonies were recovered from 58 fecal samples; they were analyzed for the presence of a class 1 integron gene cassette to detect dhfr and aad gene types, as well as other resistance genes as described above. Gene cassettes were detected in 37 (64%) of 58 samples and ranged from 1 kb to 2.2 kb in size. We detected dhfr genes in 27 (47%) of 58 samples, consisting of the following gene variants: dfrA17, dfrA12, dfrA7, dfrA5, dfrA21, dfrA1, dfrA13, and dfr7. Aad gene types were detected in 20 (34%) of 58 samples, consisting of the following variants: aadA1, aadA2, aadA5, and aadB (Table 3). No other genes were detected in these samples.
Dhfr-A17 and aadA5 were the most prevalent genes detected, present in 17 (29%) and 14 (24%) of 58 samples, respectively (Table 3). Consistent with previous literature, dfrA17 and aadA5 genes were frequently located on the same gene cassette, with 13 (35%) of 37 gene cassettes containing both genes. 42 Twenty-five (68%) of the 37 samples contained both dhfr and aad gene types.
Detection of mobile colistin resistance genes
Thirty-eight COL-resistant colonies were isolated from 10 samples and analyzed for mcr-1-5 genes. No mcr genes were detected by PCR or sequencing in these samples.
Characterization of bacterial communities by 16S rRNA sequencing
Six hundred seventeen resistant GNB isolates were recovered from 102 samples and speciated by 16S rRNA sequencing. Within these, 11 unique genera were identified; AMP-resistant bacteria consisted of 6 distinct genera, TMP-SMX-resistant bacteria consisted of 7 distinct genera, GENT-resistant bacteria consisted of 2 genera, and COL-resistant bacteria consisted of 7 genera. The relative abundance of each bacterial genus is shown in Table 2.
Number of Fecal Samples Containing Bacterial Colonies from Each Genus
Colonies were isolated from bacteria resistant to indicated antimicrobial agent. Colony growth from fecal culture may represent multiple species. Class 1 integron cassette sequences were analyzed only for TMP-SMX- and GENT-resistant isolates.
ARG, antimicrobial drug resistance gene; n/a, not applicable.
Based on Shannon diversity index, COL-resistant colonies had the highest species diversity (H = 1.9), followed by those resistant to AMP (H = 1.2), TMP-SMX (H = 0.8), and GENT (H = 0.3) (Fig. 2).
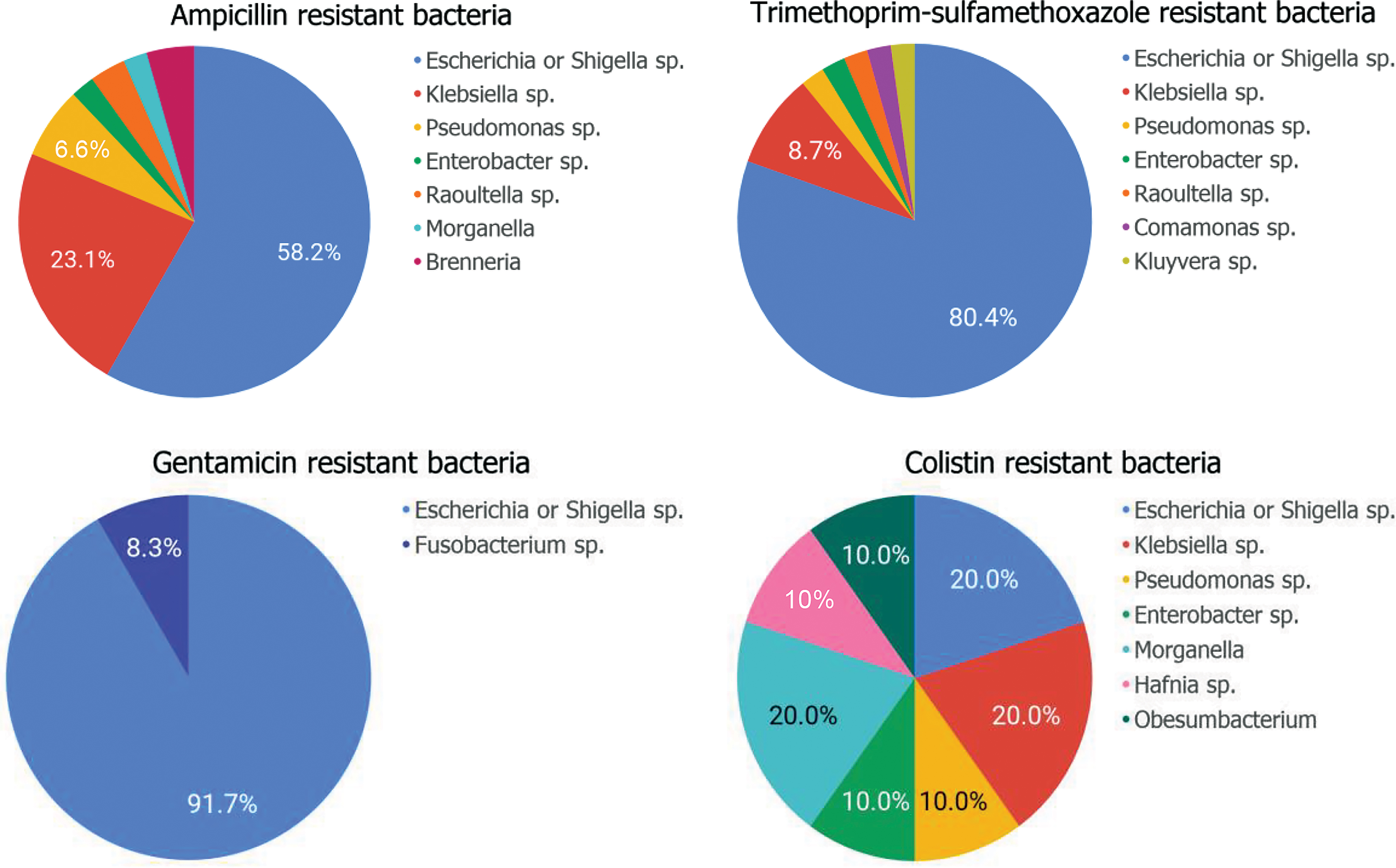
Relative frequency of genera represented by bacterial colonies exhibiting resistance to ampicillin, trimethoprim-sulfamethoxazole, gentamicin, and colistin. Color images are available online.
Species identified in these samples included the following: Escherichia spp. (E. fergusonii, E. coli, Shigella flexneri, Shigella sonnei, Shigella dysenteriae, Shigella boydii), Klebsiella spp. (K. pneumoniae or quasipneumoniae, K. oxytoca, K. aerogenes, and K. variicola), Raoultella ornithinolytica or planticola, Enterobacter spp. (E. ludwigii, E. bugandensis, E. xiangfangensis, and E. aerogenes), Pseudomonas aeruginosa, Morganella morganelli, Comamonas jiangduensis or terrigena, Kluyvera cryocrescens, Fusobacterium varium, Hafnia paralvei, and Obesum bacterium proteus.
Although Escherichia sp. and Klebsiella sp. represented the majority of our fecal samples (79%), there were 21 samples that contained other types of bacteria (i.e., Enterobacter sp., Pseudomonas sp., Shigella sp., Morganella sp., Raoultella sp., and Kluyvera sp.). Among these 21 samples, 13 (62%) harbored the antibiotic resistance genes described above.
Discussion
Our results demonstrate that there is an abundance of ARGs (β-lactamase, dhfr, and aad) carried by commensal GNB in healthy human intestine. Only 19% of rectal swab samples contained GNB that exhibited no phenotypic resistance to any of the four classes of drugs used to treat extraintestinal infections caused by enteric bacterial pathogens.
A study of gene cassettes in human UTI E. coli isolates collected from the same college community from 1999 to 2001 found similar ARGs harbored within the gene cassettes of these uropathogenic E. coli. 31 Dhfr-A17 and aadA5 were the most common cassette genes among these UTI isolates, which are the same ARGs found in GNB in fecal samples in this study. Of these ARGs, blaSHV and dhfr-7 were significantly more frequent among fecal samples (25% and 7%, respectively) than in UTI samples (1% and 0%, respectively), while dhfr-A17 was significantly more common in UTI (56%) than in fecal samples (29%). AadA1 and aadA5 were also significantly more common among UTI isolates (25% and 56%) than fecal samples (9% and 24%). Comparisons between other genes may be limited by a relatively small number of observations. The findings of this study are summarized in Table 3.
Comparison of Antimicrobial Drug Resistance Genes Found in Isolates from Fecal Samples (This Study) Compared to Urinary Tract Infection Samples Obtained from the Same Population in 2016–2017 and 2009
p-value <0.05 indicates a significant difference in genetic component between fecal samples and UPEC isolates.
UPEC, uropathogenic E. coli.
A more recent study conducted at the same college community between 2016 and 2017 found that among 233 E. coli isolates cultured from human UTI cases, 97 (42%) were resistant to AMP and 76 (78%) of these contained at least one β-lactamase gene. β-lactamase gene types from these isolates included blaTEM, blaCTX-M group 1, blaCTX-M group 9, blaSHV, and blaOXA 43 (Table 3). Forty (17%) of the 233 E. coli isolates were resistant to TMP-SMX. 43
A 2012 study by Adams-Sapper et al. analyzed Gram-negative pathogens from patients with blood stream infections (BSIs) at an urban public hospital in San Francisco for the presence of ARGs on class 1 integron gene cassettes. 45 Of 371 GNB BSI isolates from these samples, 90 contained class 1 integrons, among which 69 (77%) harbored gene cassettes. The most frequent integron cassette genes identified were dhfrA genes dhfrA12, dhfrA15, dhfrA16, and dhfrA17, and aad genes, including aadA1, aadA2, aadA5, and aadB. The most frequent gene cassette configuration among these samples was dfrA17-aadA5, found in 31 (45%) of 69 GNB isolates. Thus, there is no significant difference in the number of GNB harboring this gene cassette sequence between BSI isolates and commensal bacteria in this study. A 2013 study by the same author found that E. coli isolates from BSI patients contained β-lactamase genes similar to those of fecal GNB in our healthy volunteers 9 ; these included blaCTX-M (CTX-M-15 and CTX-M-14), blaTEM, and blaOXA. There was no significant difference between number of isolates containing CTX-M variants or resistance to TMP-SMX in clinical BSI isolates compared to those in commensal bacteria in this study. 9 These findings indicate that pathogenic GNB isolates share similar patterns of ARGs with those of commensal intestinal GNB, demonstrating further potential for acquisition of extraintestinal pathogenic E. coli of ARGs by HGT from intestinal reservoirs.
Recent literature suggests geographic differences in gut ARG distribution. A 2008 review of fecal carriage of AMR in E. coli isolated from the gut of healthy adults in Paris found that only 2 (0.6%) of 332 fecal E. coli isolates contained ESBL CTX-M-15 gene. 2 This study identified at least 8 Escherichia spp. isolates containing CTX-M-15, and 9 (9%) of 103 samples contained bacteria harboring CTX-M variants. These findings indicate a varying geographical distribution of the prevalence of AMR genes in healthy individuals, or a change in the prevalence of ARGs over time, or both, which may be affected by differences or change in ingested food products containing AMR bacteria. Indeed, a 2018 study analyzed the resistome of 1,267 human gut samples from various countries. The findings indicated country-specific gut microbial signatures and significant differences in the gut resistome among different nationalities.45,46 These studies, however, did not examine which gut GNB species carried the ARG.
In this study, isolates from a given fecal sample usually contained the same set of ARGs, even when the sample contained multiple species of bacteria. Forty-one (62%) of 76 AMP-resistant samples contained isolates in which all isolates of a given sample had the same composition of β-lactamase genes. These isolates belong to eight different GNB species (E. fergusonii, E. coli, E. marmotae, S. flexneri, S. sonnei, K. oxytoca, K. pneumoniae, and K. dysenteriae). Our findings also suggest that Klebsiella species were more likely than non-Klebsiella species to possess SHV variants (p < 0.001), while Escherichia species were more likely than non-Escherichia species to harbor TEM variants (p < 0.001). This observation suggests high frequency of HGT of ARGs among a host's intestinal GNB microbiota when ARGs enter the intestine. Within integrons, 25 (68%) of 37 gene cassettes within a given fecal sample contained all isolates with the same composition of ARGs. This further supports the claim that genes are readily horizontally transferred to one another within the human gut, with integrons playing a particularly important role in this process as they can contain multiple ARGs conferring resistance to more than one antibiotic class. 32
In the study of E. coli isolates by Ajiboye et al., pathogenic strains from food animal origins and human UTI isolates shared many of the same gene cassette sequences, 31 as well as those found in commensal bacteria in this study (Table 3). Such mobile elements carrying ARGs may be introduced into the human intestine by food contaminated with pathogenic as well as commensal bacteria harboring these ARGs, creating an abundant intestinal reservoir of ARGs that then may get transferred into GNB strains that cause extraintestinal infections.
The scope of this study was limited to analysis of aerobic or microaerophilic GNB species. This design was intentional as most mobile ARGs found in GNB pathogens are restricted to these species. However, it must be noted that the majority of gut microbial organisms are anaerobic and thus the findings of our study may underestimate the frequency of HGT of ARGs that may originate in anaerobes. In addition, our analysis of integron sequences was limited to class 1 integrons and to strains resistant to GENT or TMP-SMX, which would underestimate the prevalence of ARGs in integrons in fecal samples from this healthy college study population. Many of the ARGs in this study's AMP-resistant strains may be harbored by integrons, together with other ARG cassettes. In addition, for SMX-TMP- and GENT-resistant isolates, this study looked for ARGs through sequencing of the class 1 integron gene cassette, if present. Thus, ARGs that may be present outside of the integron cassette were not detected. Further studies that investigate HGT between a broader range of microbial species must be conducted to wholly reveal the reservoir of ARGs among intestinal bacterial organisms, and the implications for human health.
This study also utilized a screening method to identify ARGs based on the patterns of phenotypic resistance. It is possible that the GNB isolates harbored ARGs that conferred resistance to antimicrobial agents we did not use for screening. Further investigations into the resistome separated by GNB species in commensal bacteria will illuminate the implications of HGT between bacterial species.
Our study showed abundance and diversity of resistance genes among healthy adults in a particular college community. While these results may not be generalizable to other communities, it highlights the need for community-based comparisons between commensal and pathogenic bacteria. These observations made in a non-health care environment raise questions about how these ARGs were introduced into the intestines of people residing in this community, including food and environment as sources. It calls for further investigation into possible risk factors for acquisition of ARGs other than just exposures to antibiotics, as they clearly pose a risk to public health. The findings in this study illuminate the relationship between gut microbial ARG content and infections such as UTI caused by Gram-negative bacterial pathogens. These observations may have global implications for the spread of antibiotic resistance through mobile genetic elements, as infections become increasingly difficult to treat with current antibiotic therapies.
Footnotes
Acknowledgments
We would like to thank Reina Yamaji for her assistance in the study design process, as well as Clarissa Araujo Bourges for her consistent support in classification of study isolates. We would also like to thank the University instructors who were instrumental in recruitment for this study. We would also like to the University Department of Environmental Health and Safety for their assistance in ensuring safe shipment of biological specimens.
Disclosure Statement
No competing financial interests exist. The funding bodies did not play a role in the experiments or their interpretation.
Funding Information
This study was supported by Centers for Disease Control and Prevention program to combat antibiotic resistance under BAA no. 200-2016-91939.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.