
Abstract
Antimicrobial peptides (AMPs) are produced by all forms of life, ranging from eukaryotes to prokaryotes, and they are a crucial component of innate immunity, involved in clearing infection by inhibiting pathogen colonization. In the recent past, AMPs received high attention due to the increase of extensive antibiotic resistance by these pathogens. AMPs exhibit a diverse spectrum of activity against bacteria, fungi, parasites, and various types of cancer. AMPs are active against various bacterial pathogens that cause disease in animals and plants. However, because of the coevolution of host and pathogen interaction, bacteria have developed the mechanisms to sense and exhibit an adaptive response against AMPs. These resistance mechanisms are playing an important role in bacterial virulence within the host. Here, we have discussed the different resistance mechanisms used by gram-positive and gram-negative bacteria to sense and combat AMP actions. Understanding the mechanism of AMP resistance may provide directions toward the development of novel therapeutic strategies to control multidrug-resistant pathogens.
Introduction
Microbes are possibly the most ancient and adaptable organisms on the earth as they evolved over 3.5 billion years ago.1,2 Microbes reside in diverse as well as extreme habitats ranging from the gut of a different variety of insects and invertebrates, skin, blood, eye, and internal organs of different animals and human, hydrothermal vents, depth of the ocean, and poles.3,4 While microbes adopt diverse mechanisms to survive in different habitats, antimicrobial agents are the major causes that have helped them to coevolve within or in close proximity of human host and even in the presence of more advanced clinical settings.5,6
The rapid emergence of resistance has been observed against almost all classes of antimicrobials in the recent past.7,8 In the current scenario, judicious use or novel alternatives of conventional antibiotics are the only solution to combat the resistance problem.9,10 Antimicrobial peptides (AMPs) are considered one of the promising alternatives to conventional antibiotics that are currently under development.10–12 AMPs are usually short, cationic, and amphiphilic in nature with immunomodulatory activities.13,14
Importantly, bacteriocins are bacterial AMPs that represent a huge repertoire of potential AMPs with possible therapeutic applications with diverse bioactivities.11,15,16 In general, bacteria are much less prone to develop resistance against AMPs in comparison with conventional antibiotics.17–19 However, it is extremely difficult to predict, when and how resistance may evolve and spread against specific AMPs. In a recent study, it has been reported that AMPs have different pharmacodynamic properties when compared with conventional antibiotics. By implementing these differences in the theoretical framework, a lower probability is revealed to evolve resistance against AMPs. 20 Here, we have investigated the possible resistance mechanisms used by gram-positive and gram-negative bacteria against AMPs (Fig. 1), which could provide more insight for the development of future therapeutic strategies to fight against multidrug-resistant bacterial pathogens.

Diagrammatic representation of resistance mechanism against AMPs in gram-positive (left) and gram-negative (right) bacteria. Proteolytic degradation by extracellular, cytosolic (after uptake), and outer membrane-bound proteases, (1) sequestration within extracellular matrix-like biofilms composed of capsular polysaccharides and exo-polysaccharides, (2) electrostatic repulsion by alanylated LTAs and WTAs, (3) enhanced rigidity in outer membrane by lipid A acylation, (4) electrostatic repulsion by amine compound (aminosugars, PEA, or glycine) added lipid A, (5) electrostatic repulsion by aminoacylated PG, (6) pentapeptide alteration to restrict entry of lipid II binding AMPs, (7) efflux and export of AMPs by efflux pumps (8). AMPs, antimicrobial peptides; LTA, lipoteichoic acid; PEA, phosphoethanolamine; PG, phosphatidylglycerol; WTA, wall teichoic acid. Color images are available online.
How to Distinguish Among Bacterial Resistance?
The rapid emergence of bacterial resistance against available antimicrobial agents is a major problem in clinical settings.21,22 However, since the incidence of developing resistance against AMPs is low, it is important to investigate a resistance development process, which can differentiate the properties of resistant bacterial strains and thus making it possible to find a solution or recommend a suitable treatment. These efforts may enhance the efficacy of treatment and reduce the medical expenses. Disk diffusion and broth dilution are two conventional laboratory methods commonly used to determine resistance or susceptibility toward antimicrobial agents. Primarily, bacterial resistance to antibiotics is measured quantitatively by determining the minimum inhibitory concentrations through the macro- or microdilution method in a conventional laboratory setting. 23
Many commercially available antimicrobial resistance detection systems are based on standard conventional manual methods and provide almost similar results. The fully automated or semiautomated systems such as VITEK2, BD Phoenix, and MicroScan WalkAway are used to determine antimicrobial succeptability. 24 These systems rapidly detect bacterial resistance against antibiotics at low cost and are helpful in preventing the rapid spread of resistant strains.25–27 In the case of the heterogeneous population of antimicrobial-resistant bacteria where different levels of antimicrobial resistance have been observed, highly elegant techniques have to be used to detect different levels of resistance. GISA (glycopeptides intermediately susceptible to Staphylococcus aureus) and its heterogeneous variant (hGISA) are a good example of this, where the phenotypes of bacterial isolates are very common but show a variable level of reduced susceptibility toward glycopeptides.28,29
Bacterial resistance toward different antimicrobials is generally determined phenotypically by exposure to various antimicrobials. High-throughput subtyping methods have been found to be more beneficial over phenotypic assays not only in terms of time, efficiency, and accuracy, but also for avoiding other hurdles such as less expression of resistant phenotype.30–32 Therefore, high-throughput screening methods based on genomic, transcriptomic, and proteomic analyses are used to determine bacterial resistance and to minimize spreading of infection in clinical settings.33,34 Investigating the resistome may further provide insight into the mechanism of action and bacterial response toward resistance development.35,36 Therefore, all these methods for the determination of antimicrobial resistance support the rapid identification and application of targeted antimicrobial therapy in infectious diseases and may increase the life span of infected individuals. Although several new technologies such as digital imaging and genomic and proteomic approaches are being introduced, no single universally accepted method is available so far to detect bacterial resistance to antimicrobials. Therefore, it is important to further investigate the rapid evolution and mechanism of resistance against antimicrobials.
Cell Wall Modifications and Resistance Development
Lipid II
Lipid II is the precursor of cell wall biosynthesis, which is present in the outer leaflet of the bacterial cytoplasmic membrane. Peptidoglycan is a major structural component of bacterial cell wall, which contains repeating units of N-acetylmuramic acid (MurNAc) and N-acetylglucosamine (GlcNAc) and further crosslinked via peptides attached to MurNAc. Penicillin-binding protein (PBP) is a bifunctional enzyme that catalyzes the polymerization of sugar units and peptide crosslinking (transpeptidation) using lipid II as a substrate. 37 Upon binding to AMPs, lipid II restricts the peptidoglycan biosynthesis by physically sequestrating transpeptidase and transglycosylase, otherwise stabilizing the bacterial cell wall by crosslinking. 38
Recently, it has been reported that nisin and gallidermin interact with cell wall components other than lipid II. These components include cell wall teichoic acid (WTA) biosynthesis pathway, lipid III (undecaprenol-pyrophosphate-N-acetylglucosamine), and lipid IV (undecaprenol-pyrophosphate-N-acetylglucosamine-N-acetylmannosamine). Nisin and gallidermin interact with the precursors of cell WTA that ultimately stimulate pore formation in lipid bilayers. 39 Moreover, quantification of lipid II levels in different nisin-sensitive strains has revealed that nisin resistance is independent of lipid II levels. 40
d -Alanyl-lipoteichoic acid A
Gram-positive bacterial cell wall contains a thick peptidoglycan layer, linked with teichoic acids. Teichoic acids are further classified into WTA (linked with muramic acid residues of peptidoglycan)
41
and LTA (lipoteichoic acids), which are macroamphiphiles anchored hydrophobically to the glycolipid components in the outer layer of cytoplasmic membrane.
42

Cell wall- and membrane-related mechanisms of AMP resistance.
Sensitivity to cationic AMPs such as gallidermin, defensins, and protegrins has been shown to increase in dltA, dltB, and dltD deletion mutants of Staphylococcus Xylosus, which are devoid of
Alanylation of LTA also appears to affect the sensitivity of Streptococcus pneumoniae toward nisin.
48
The dltA deletion mutants of Listeria monocytogenes have shown a severe reduction in colonization in a mouse infection model. As the amount of surface protein involved in virulence (ActA, InlA, and InlB) remains unaffected, it suggests that
Penicillin-binding proteins
PBPs play an essential role in peptidoglycan assembly. They are classified as class A or class B, on the basis of structure and catalytic activity of the N-terminal module. The C-terminal ends of both classes catalyze peptide crosslinking between two neighboring glycan chains through their transpeptidase activity. 51 The N-terminal module of class A is involved in glycan chain elongation through its glycosyl-transferase activity 52 and the N-terminal module of class B is involved in the interaction with other proteins during septation and also regulates bacterial shape. 53
PBPs are well reported for their role in resistance development against antimicrobials. In a nisin-resistant strain of L. monocytogenes, a spontaneous and significant increase in putative PBP expression levels is observed in comparison with the wild-type susceptible strain. Interestingly, it has been found that this strain is more resistant toward mersacidin but sensitive to a variety of beta-lactam antibiotics. 54 Similarly, higher levels of PBP2A (a class B PBP) expression are observed in a nisin-resistant mutant of L. lactis IL1403 in comparison with the wild-type strain. This suggests that thicker and densely packed cell walls play an important role in the development of resistance against AMPs (Fig. 2). 49 The role of PBP4 (a class A PBP) of L. monocytogenes in the development of nisin resistance has also been confirmed by making a deletion mutant, which shows higher sensitivity to nisin. 55 Furthermore, PBP4 is also regulated by increased expression of LiaS (a histidine kinase element), which results in elevated levels of PBP and thus nisin resistance in Listeria. 56
Cell Membrane Modifications and Resistance Development
Outer membrane modifications
Gram-negative bacteria usually display resistance toward antimicrobials as they have an outer membrane (OM), which prevents the entry of antimicrobials to the cytoplasmic membrane. 57 The outer most layer of the OM facilitates the attachment of lipid A, which is an anionic dimer of glucosamine-linked fatty acid chains with flanking polar phosphate groups. The lipopolysaccharide (LPS) complex is formed by covalently bound lipid A units to a core polysaccharide and a specific O chain consisting of repeating oligosaccharide units. The complex without O chain is called lipo-oligosaccharide (LOS).58,59 Sublethal stress conditions such as heating and freezing and chelating agents such as EDTA can degrade the LPS layer and increase the susceptibility of gram-negative bacteria toward AMPs.60–62
Modifications in LPS are usually mediated by environmental factors through two-component signaling systems. LPS modulation often leads to increase in virulence, alters TLR4-mediated inflammatory response, and thus helps in the development of resistance against AMPs. 63 Modifications in LPS lipid A lead to the development of resistance against AMPs by compromising OM permeability and neutralizing the LPS negative charges. Occurrence of such type of modifications in LPS and the role of modified LPS in developing resistance against AMPs have been reported in Salmonella typhimurium, Yersinia sp., Escherichia coli, Pseudomonas aeruginosa, and Neisseria sp. 64 PagP is an OM enzyme of S. typhimurium, which catalyzes the transfer of palmitoyl group from phospholipids to lipid A, resulting in the synthesis of hepta-acylated lipid A.
This modification has strengthened the OM permeability barrier and enhanced resistance against AMPs such as C18G and protegrin. 65 Interestingly, PagP is found inactive in OM, but degradation of OM by self-uptake of AMPs acts as a stimulant for the activation of PagP. 66 The arnBCADTEF operon of Salmonella spp. is responsible for the biosynthesis and L-Ara4N residue to the 4′-phosphate group of lipid A. The neutralization of negative charge of the 4′-phosphate group plays an important role in resistance development toward polymyxin B. 67 In accordance with this, modification of LPS with L-Ara4N residue has been observed in Burkholderia cepacia. 68 On the contrary, Franscisella novicida has developed resistance toward polymyxin B in a different manner where lipid A phosphatase, LpxF, cleaves to the phosphate group at the 4′ position of lipid A. 69
The Rcs phosphorelay is another system that regulates AMP resistance in S. typhimurium via activation and expression of periplasmic protein Ydel, which is responsible for the oral virulence in vivo. 70 SurA, tolB, and pgm are other elements of AMP resistance, which are also involved in virulence regulation. SurA and tolB have been reported to stabilize the cell membrane, while pgm has been shown to be involved in LPS biosynthesis and in conferring resistance against polymyxin B.71,72 Glycosylation of LPS by almEFG operon enzymes has been reported to confer resistance against polymyxin B in Vibrio cholerae. 73 PmrAB is a two-component system that regulates LPS modification in Acinetobacter baumannii. It has been reported that PmrAB is involved in the development of resistance against colistin. 74 Other than LPS, anionic capsular polysaccharide of Klebsiella pneumoniae, P. aeruginosa, and S. pneumoniae has been reported to be involved in the development of resistance against polymyxin B and human β-defensin. 75
Phospholipid modifications
Wide variations in phospholipid composition and relative abundance among different bacterial species have been reported under different environmental conditions and growth phases. This relative variation of phospholipids in the bacterial cell membrane has an important role in making them resistance against different antimicrobials. 76 Phosphatidylglycerol (PG) and diphosphatidylglycerol (DPG) are the most common bacterial phospholipids. 77 Phosphatidylethanolamine (PE) is found only in Enterobacteriaceae and Bacilli but not in Staphylococcus or Listeria sp.78–80 Enhanced levels of PG over DPG were observed in a nisin-resistant strain of L. monocytogenes, confirming the role of phospholipids in resistance development. 81
Higher levels of anionic phospholipids such as DPG over cationic phospholipids such as PG, PE, and phosphatidylcholine (PC) result in increased penetration of AMPs such as nisin. 82 It has also been observed in nisin-resistant L. monocytogenes ATCC 700302 when the anionic phospholipid level is decreased. 83 Furthermore, nisin-resistant bacterial cells have been reported to have decreased levels of phospholipids, while a higher ratio of straight to branched-chain fatty acids, in comparison with nonresistant cells. 84 The addition of lysine to phospholipids (lysylation) results in shifting of PG from the anionic to cationic form, which repels cationic AMPs by reducing their binding to the cell membrane. Lysylated phospholipids are well studied in S. aureus, and have also been reported in gram-negative bacteria such as Rhizobium tropici and Caulobacter crescentus.85–87
Capsule production
The bacterial capsule is an additional and outer most protective barrier against AMPs, which is composed of repeating long-chain polysaccharides. 88 It has been reported that the K. pneumoniae capsule plays a role in enhancing the resistance levels against AMPs such as defensins, lactoferrins, and polymyxins. It has been shown that a higher level of capsular polysaccharides leads to reduced AMP binding to the OM and thus displays an increase in resistance toward polymyxins. 89 Capsule production has been shown to be associated with elevated resistance levels toward human AMP LL-37 in a capsule deletion mutant of Neisseria meningitides. 90 Furthermore, expression of capsule biosynthetic genes siaC and siaD has been found to be upregulated upon exposure to the sublethal doses of LL-37. In addition, it has been reported that K. pneumoniae and P. aeruginosa use their capsular polysaccharides in the development of resistance against polymyxinB and HNP-1. 75 Therefore, it is suggested that capsules have a potential role in resistance development against AMPs. 90
Biofilm Formation and Resistance Development
Biofilm formation is an important strategy of bacteria to develop resistance against antimicrobials. Biofilms composed of sessile bacterial populations that adhere to the surface in a highly organized manner for nutrient availability and circulation. 91 The slimy extracellular matrix secreted by bacteria during biofilm formation plays a key role in adherence to the surface and protection against outside killing agents such as AMPs. This slimy matrix contains a mixture of diverse substances, including cellulose, proteins, lipids, nucleic acids, and teichoic acids.92,93 Bacteria can form biofilms on hospital equipment, ventilators, and catheters involved in hospital-acquired infections. Pathogenic bacteria can also form biofilms on teeth surfaces and respiratory tracts, where after their maturation they become more heterogeneous and thicker, thus contributing to chronic infections. 94
Pseudomonas biofilm consists of a high level of polysaccharide alginate, which makes the biofilms thicker and more heterogeneous. Alginate overproducing Pseudomonas strain showed more resistance ability toward AMPs. 95 This has been shown further by the expression of Pseudomonas biofilm genes in E. coli, where flat and unstructured biofilms transformed into a complex architecture that shows high resistance to colistin. 96 Polysaccharide-mediated resistance toward AMPs has been reported in K. pneumoniae and B. cepacia, which have been isolated from cystic fibrosis patients and shown to be highly resistant toward LL-37 and human β defensin-3 (hBD-3). 96 In P. aeruginosa and S. typhimurium biofilms where extracellular DNA is an integral component, it has been reported that extracellular DNA chelates cations of AMPs and contributes to resistance development.97,98 In addition, the inducible biofilm gene psrA of P. aeruginosa has been reported to be associated with increased levels of resistance against polymyxin and indolicidin. 99 In total, biofilms are significantly contributing to the resistance development against AMPs and other antimicrobials.
Multiple Peptide-Resistant Factors
Lysyl-phosphatidylglycerol (L-PG) is generated by lysine esterification of one of the two hydroxyl groups of PG, conveying a net positive charge on the cytoplasmic membrane.100,101 Lysinylation is executed by multiple peptide-resistant factors (MprF), catalyzing the transfer of lysine residues from lysyl-tRNAs (transfer RNAs) to PG (Fig. 2). MprF play an important role in the development of resistance against AMPs because several bacterial pathogens are reported to have L-PG in their cytoplasmic membranes, including S. aureus,102,103 L. monocytogenes,80,104 and Mycobacterium tuberculosis. 105
MprF proteins are integral membrane proteins with two well-conserved functional domains, the hydrophilic cytoplasmic domain at the C terminus, which synthesizes L-PG, and a large hydrophobic flippase domain at the N terminus, which facilitates flipping of L-PG. 106 Both the domains of MprF are responsible for the development of resistant phenotype as one domain is involved in synthesizing, while the other is involved in translocating L-PG to the outer layer of the membrane107,108 L-PG-deficient mutants of S. aureus have displayed enhanced susceptibility to gallidermin and nisin, which further confirms the role of MprF in resistance development. 100 Similar effects are also observed in L. monocytogenes deletion mutants of L-PG, which show increased susceptibility toward gallidermin and human defensins HNP-1 and HNP-2. 104
Some gram-positive bacterium such as Clostridium perfringens SM101 contains two homologous genes of MprF. 109 MrpF1 encrypts alanyl phosphatidylglycerol synthase (A-PGS) while MrpF2 encrypts lysyl phosphatidylglycerol synthase (L-PGS).107,109 In Enterococcus faecium, both MprF1 and MprF2 homologues have been identified, but only MprF2 has been found to be engaged in aminoacylation and formation of L-PG and A-PG. 110 Enterococcus faecalis contains only single MprF, but forms both L-PG and A-PG. 111 Furthermore, MprF-encrypted A-PGS protein of C. perfringens has been produced in S. aureus and by making MprF deletion mutant, it has been demonstrated that the presence of A-PG instead of L-PG does not affect the susceptibility toward AMPs such as nisin and gallidermin. 112
In MprF deletion mutant of Pseudomonas, A-PG deficiency increases the susceptibility toward lipopeptide antibiotic daptomycin and other antibiotics such as ampicillin, oxacillin, and cefsulodin,113,114 suggesting that MprF play an important role in resistance development in both gram-positive and gram-negative bacteria. A parallel defense mechanism has also been reported in M. tuberculosis, where the lysX gene is found responsible for the production of two domains, lysyl-transferase (MprF) and lysyl-tRNA synthetase (LysU), producing L-PG. The deletion mutant of lysX exhibits increased susceptibility toward cationic antibiotics and AMPs. 105 As a whole, the presence of aminoacylated PGs is an alternate means for bacteria to protect themselves against AMP action and thus plays an important role in the development of resistance against AMPs.
Fab Operon
Fab operon is involved in the cytoplasmic membrane biosynthesis through saturation and elongation of phospholipids.115,116 Downregulation of fab operon (fabDG1G2Z1Z2) gene expression is reported in nisin-resistant strain of L. lactis. 49 the downregulation of fab operon decreases saturated fatty acid content in the membrane to alter the membrane density. These changes in membrane composition encourage the interaction of nisin with the cytoplasmic membrane to elevate resistance levels. Further investigation is required to understand the exact role of fab operon in the development and evolution of nisin resistance.
Bacterial Two-Component Systems Associated with AMP Resistance
Two-component systems of bacteria contain regulatory units that sense and respond in different environmental conditions. Usually, they contain a sensor, encoding sensory histidine kinase, and a regulator, encoding a cogent response regulator. Two major groups of two-component systems have been identified in bacteria to maintain cell membrane integrity and antimicrobial resistance or detoxification processes. One group is the BceS-like intramembrane-sensing histidine kinases (IM-HK), functionally and genetically linked to ABC transporters, and the second group is LiaS-like IM-HK, which functions as part of the two-component systems.
BceRS-Like Two-Component Systems
BceRS of Bacillus subtilis
BceRS-like IM-HK are functionally and genetically associated with ABC transporters (BceAB) that are composed of two transmembrane helices without any extracellular sensory domains.117,118 The presence of AMPs is recognized by the associated transporters along with IM-HK, which leads to the activation of cognate response regulator to induce transporter gene expression and ATP hydrolysis (Fig. 3).119,120 This pathway is associated with the development of AMP resistance in many homologous systems present in B. subtilis, S. aureus, Streptococcus mutans, E. faecalis, and Lactobacillus casei.120–124 LlrG/KinG is a homologue gene present in L. lactis on either side of the VanZ-like protein encoding gene. Downstream of the LlrG/King, an ABC transporter gene, ysaBC, is present whose expression is induced by nisin and confers resistance against nisin. 49 VanZ gene is also present in E. faecium, which confers resistance for teicoplanin. 125
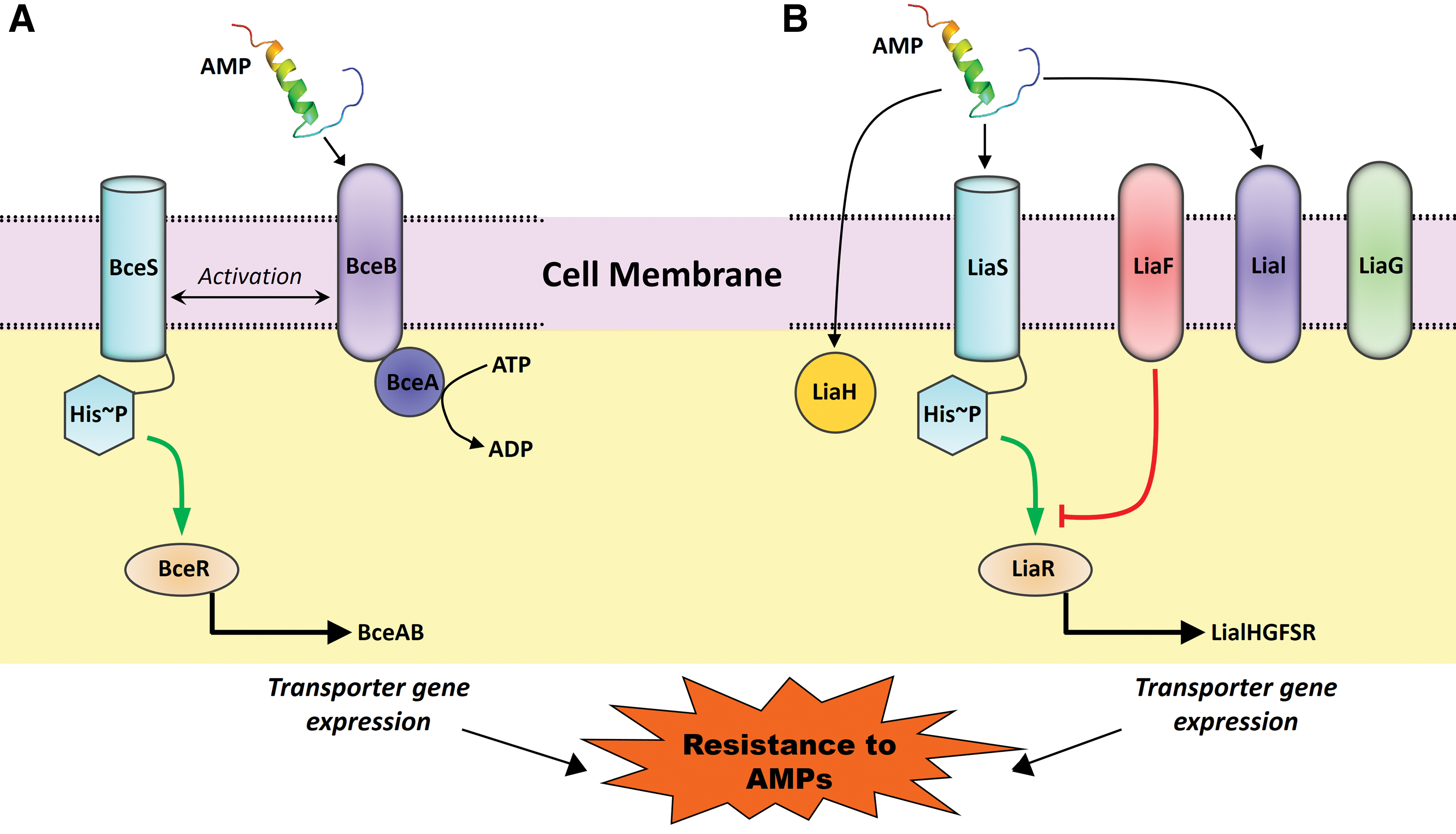
Two major types of two-component systems responsible for AMP resistance.
Other than BceRS-AB, sensing and detoxification modules of two paralogous peptides YxdJK-LM and PsdRS-AB are reported in B. subtilis genome. The YxdJK-LM system confers resistance against human AMP LL-37, 126 while PsdRS-AB responds mainly to lipid II-binding bacterial AMPs such as nisin, gallidermin, subtilin, actagardin, and lipid binding lipopeptide enduracidin. 121 These reports suggest the role of BceRS-like two-component systems in the development of resistance against AMPs.
Bacitracin resistance-associated of S. aureus
BraRS (bacitracin resistance-associated) is an IM-HK two-component system of S. aureus, which plays a crucial role for the development of resistance against bacitracin, nisin, and nukacin ISK-1.119,127,128 BraRS has been referred by three different names such as the BraRS, 119 BceRS, 129 and NsaRS130,131 and its expression is upregulated upon treatment with nisin and gramicidin. 127 BraRS is also associated with the ABC transporter protein called BraDE, which plays a crucial role in resistance development against bacitracin and nisin. A single mutation (A20E) in BraRS has been reported to enhance the resistance level 8–16-fold for nisin. 130 Thus, BraRS plays a crucial role in resistance development against AMPs in S. aureus.
GraRS of S. aureus
GraRS of S. aureus closely resembles BceRS of B. subtilis and is referred as VraCA. 132 It is associated with the development of vancomycin-intermediate S. aureus (VISA)-resistant phenotype. 133 GraRS induces the expression of ABC transporter gene, vraFG, and enhances resistance against vancomycin. 134 GraXRS-VraFG is reported as a five-component system involved in AMP sensing and signal transduction crucial for resistance development against AMPs in S. aureus. 135 The role of GraRS IM-HK in resistance development against AMPs has been reported in the GraRS mutant, which displays high susceptibility toward nukacin ISK-1 than nisin A, but no resistance against bacitracin. 128 Single and multiple amino acid changes within GraR have been found to be associated with the increased resistance against AMPs, including vancomycin and polymyxin B.133,136 GraRS equivalent, ApsRS, is reported in Staphylococcus epidermidis and is induced by nisin and other antimicrobials. 137
AnrAB transporter of L. monocytogenes
AnrAB transporter is an ABC-like transporter of L. monocytogenes and plays an important role in nisin resistance. By screening L. monocytogenes mutants with increased sensitivity to gallidermin, bacitracin, and a large number of lactam antibiotics, the permease module of AnrAB transporter has been identified, 138 and thus, AnrAB was recognized as a multidrug resistance transporter that plays a crucial role in the development of resistance against AMPs in L. monocytogenes. VirRS and RpoN are response regulators of two-component systems and also responsible for AnrAB regulation.139,140 Furthermore, AnrAB has high resemblance with the ABC transporter of S. pneumoniae D39 (Sp0912-Sp0913) and provides innate resistance to nisin, bacitracin, and gallidermin. Moreover, expression of Sp0912 is induced by high concentrations of nisin, bacitracin, and human AMP LL-37, 141 found to be regulated by another two-component system, named Rr01-Hk01, 117 such as AnrAB. All these reports suggest the role of two-component systems along with ABC transporters in resistance development against AMPs in bacteria.
LiaRS and LiaRS-Like Two-Component Systems
The LiaRS, a two-component system present in almost all members of Firmicutes, is engaged in cell membrane stress responses caused by the inhibitors of the lipid II cycle. The promoter of Lia operon responds to cell wall-active AMPs such as bacitracin and nisin and interferes with the lipid II cycle.142,143 LiaR plays a crucial role in resistance development against nisin via an ABC transporter permease encoded by the gene Lmo1746.144,145 LiaR mutant of group B Streptococcus is highly sensitive to cell wall damaging AMPs such as bacitracin, colistin, and nisin, in comparison with the parental strain. In the absence of LiaR, groupB Streptococcus strains become avirulent and do not cause sepsis and pneumonia in mouse models. 146 LiaRS homologue, CesSR, and LiaF homologue, lmg1650, are found in L. lactis.143,147 CesR mutant of L. lactis has shown about twofold higher sensitivity toward AMPs such as nisin, plantaricin C, and bacitracin when compared with the parental strain. Moreover, AMPs such as laactococcin 972, plantaricin C, and bacitracin act as inducers of the Llmg1650 promoter in a concentration-dependent manner, while nisin does not induce this promoter. 148 All these results suggest that LiaRS and LiaRS-like two-component systems play a crucial role in resistance development against AMPs (Fig. 3).
VraSR of S. aureus
VraSR is a vancomycin resistance response regulator that contributes to the development of resistance against cell wall targeting AMPs, and protection against cell damage through the modification of peptidoglycan biosynthesis pathway. A number of genes under the control of VraSR have been identified in S. aureus for lactams and glycopeptide resistance. Among them, the vraDE gene encodes an ABC transporter and is involved in nisin and bacitracin resistance. Notably, this ABC transporter is also regulated by BraRS, which suggests the multifunctionality of ABC transporters. 119 In addition, VraSR is activated when cell wall biosynthesis is inhibited by the treatment of mersacidin, daptomycin, and vancomycin.128,149 However, nisin treatment does not induce vraSR expression and suggests the involvement of other factors that vary with AMPs. 150
Treatment with lantibiotic nukacin ISK-1 induces vraSR expression and suggests that the differential expression of vraSR upon treatment of different AMPs is due to their structural differences and mode of action. 128 In VraSR deletion mutants, a prominent increase in susceptibility toward cell wall inhibiting AMPs such as nukacin ISK-1, bacitracin, and daptomycin is observed.128,151 VraSR exerts its resistance mechanism via cell wall thickening, which hinders the AMPs to reach its target molecules such as lipid II. However, resistance to mersacidin (whose target molecule is lipid II) does not develop although overexpression of cell wall synthesis occurs through the induction of vraSR two-component systems.149,152 Moreover, mutation in VraS (L114S and D242G) results in elevated resistance to mersacidin. 153 Therefore, more studies are required to understand the role of VraSR in the development of resistance against AMPs with respect to cell wall thickening.
LisRK and VisRS of L. monocytogenes
The LisRK is a two-component system of L. monocytogens (originally reported in strain LO28), which plays an important role in virulence and protection against antimicrobials such as cephalosporin and nisin.154,155 Another two-component system of L. monocytogene is VirR/VirS, which also plays a role in virulence through the upregulation of VirR by virulence regulator PrfA. 156 Interestingly, it has been reported earlier that VirRS can regulate genes such as dltA, MprF, and anrB that play a role in nisin resistance.138,139 All these results suggest a possible role of LisRK and VirRS in the regulation of resistance development toward AMPs in L. monocytogenes.
CprRK of Clostridium difficile
The CprRK is a two-component system of C. difficile, where Cprk functions as a sensor kinase and regulates orphan response. AMPs such as nisin, gallidermin, subtilin, mutacin 1140, and cinnamycin activate the CprK-CprR system and contribute to resistance development against these AMPs. 157 Furthermore, the CprRK system regulates expression of gene encoding ABC transporter, CprABC, which is found adjacent to CprK in the genome. A decrease in resistance against nisin and gallidermin has been observed upon insertional disruption of one of the CprABC transporter genes. Thus, CprRK, along with the CprABC transporter system, plays a crucial role in resistance development toward AMPs. 158
Role of Efflux Pumps in Resistance Development to AMPs
Bacterially encoded efflux pumps are well known for their role in resistance development against conventional antibiotics.159,160 The role of efflux pumps in the development of resistance against natural substances produced by host such as hormones and defense AMPs has also been reported.161,162 Gram-negative bacterial efflux pumps are involved in resistance development against AMPs, which belong to the RND (resistance, nodulation, and cell division) family transporters. 163 The RND family transporter consists of an inner membrane AMP antiporter, an OM transporter, and a periplasmic accessory protein, which can stabilizes the whole efflux system.164,165 RND family efflux pumps are well reported in different Neisseria species for their involvement in resistance development. 166
MtrCDE efflux pumps of Neisseria gonorrhoeae and N. meningitides have been reported to develop resistance against AMPs such as LL-37 and protegrin-1.167,168 It has also been demonstrated that VexAB-To1c, an RND family efflux pump, can provide protection and develop resistance against polymyxin B. 169 In addition, AcrAB efflux pumps of K. pneumoniae have been reported to act against cationic AMPs such as polymyxin B and human defensins, including HNP-1, HBD-1, and HBD-2. 170 Apart from these, MisR of N. gonorrhoeae, a two-component response regulator, has been found to be involved in intrinsic resistance development against cationic AMPs such as LL-37. 171 RosAB, a temperature-regulated MFS superfamily efflux pump of Yersinia enterocolitica, is reported to be involved in resistance development toward cationic AMPs such as polymyxin B. 172
AMP Degradation and Resistance Development
Proteases play a dynamic role in the maturation and degradation of AMPs. Host proteases process the AMP precursors into biologically active AMPs, while active AMPs can also be degraded into inactive peptide fragments by bacterial proteases (Fig. 4). In this way, bacteria evolved to develop resistance against AMPs. LL-37, a human cathelicidin AMP, has been reported to be degraded by an elastase of P. aeruginosa and a protease of Proteus mirabilis. 173 The zinc metalloprotease of P. mirabilis, ZapA, is reported to degrade LL-37 and β-defensin1. 174 ZmpA and ZmpB, two zinc metalloproteases isolated from Burkholderia cenocepacia, have been reported to degrade the LL-37 and β-defensin1, respectively. 175

Other examples of AMP resistance mechanisms. Bacterial protease-mediated degradation of AMPs, protein-mediated sequestration of AMPs, and downregulation of AMPs by pathogenic bacteria shown here. Color images are available online.
Porphyromonas gingibalis, an oral cavity pathogen associated with chronic periodontal disease, is reported to secrete proteases known as gingipains, which cleaves AMPs at the site after arginine and lysine residues and thus degrades and inactivates LL-37 and β-defensin3.176,177 AMP degrading omptin proteases are also present in the Enterobacteriaceae family and degrade many host proteins or peptides.178,179 OmpT is another optimin, which has been reported to cleave an AMP protamine. 180 The crop of Citrobacter rodentium is another example of optimin family proteases, which prevents the activation of PhoPQ two-component system by AMPs. 181 In addition to this, OmpT of enterohemorrhagic and enteropathogenic E. coli strains has been shown to degrade LL-37. 182 Furthermore, capsular fragment 1 and Pla of Yersinia pestis have been found to cleave the pulmonary AMP cathelicidin. 183 PgtE, a PhoP-regulated OM protease of S. typhimurium, has also been reported to cleave α helical AMPs and thus provides the chances for the development of resistance against AMPs. 184
Protease-mediated degradation of AMPs is a well-known phenomenon for bacteriocins produced by bacteria. Nisinase is a protease produced by some resistant Bacillus strain and it cleaves nisin. 185 Nisinase is produced during sporulation in strains of Bacillus cereus and Paenibacillus polymyxa, and provides resistance against AMPs such as nisin. 186 Interestingly, nisinase is active against AMP such as subtilin, but not against antibiotics such as bacitracin, polymyxin, and gramicidin. 185 One such example is nisin-resistant protein NSR protease, which is localized on the cell membrane and degrades nisin by cleaving its C-terminal tail. 187
Resistance Development by Protein-Mediated Sequestration
Extracellular or surface-linked proteins of some gram-positive bacteria directly bind to the AMPs and inhibit their activity on the cell membrane.188–192 In this way, protein-mediated sequestration of AMPs plays a significant role for resistance development in bacteria (Fig. 4). Streptococcal inhibitor of complement (SIC), a hydrophilic secreted protein produced by Streptococcus pyogenes, binds and sequesters AMPs, including LL-37, α-defensins, and lysozyme.193–195 Staphylokinase is another extracellular protein secreted by S. aureus, which binds and sequesters AMPs such as cathelicidin, human defensin HNP-1, and HNP-2.191,192 Proteins present on the cell surface also contribute to resistance development by sequestering AMPs. For example, the M1 protein of S. pyogenes found on the cellular surface of many clinical isolates is associated with host tissue adherence and invasive diseases.196,197 M1 protein binds LL-37 by its hypervariable extracellular portion, sequesters it, and reduces the activity of LL-37, resulting in streptococcal survival in neutrophil extracellular traps. 188
Infection and Altered AMP Production: A Key to Resistance Development
Pathogenic bacteria are coevolved with the host and develop different mechanisms to overcome antimicrobial response created by the host. While cell wall or cell membrane modifications are the major strategies to combat AMPs provided by the host, pathogenic bacteria repress transcription of AMPs and thus evade their actions. By downregulating AMP production, pathogenic bacteria develop resistance against AMPs and facilitate their invasion into the host (Fig. 4). Human enteric pathogens, V. cholerae and E. coli, downregulate the expression of cathelicidin, LL-37, and HBD-1. Cholera toxin (CT) and labile toxin (LT), produced by V. cholerae and E. coli, respectively, regulate the expression of AMPs via multiple downstream signaling pathways. 198 However, in some cases, V. cholerae isolated from the small intestine biopsies of infected patients does not show any downregulation of the AMP expression at the transcriptional level. 199 Shigella spp. inhibits the expression of β-defensins and LL-37 in intestinal epithelial cells via type III secretion system and the mxiE transcriptional regulator.200,201 Peneth cells of small intestine constitutively produce α-defensins called cryptidins in mice. 202 Absence of crptidin increases the susceptibility of mice to Salmonella infections. 203 Enteric Salmonella infections inhibit AMP production by the peneth cells via direct interaction between Salmonella and small intestinal epithelium. 204
Repression of AMP gene expression has also been reported in invertebrate hosts during vibrioses. Transcriptional repression of damicornin, an AMP produced by scleractinian coral Pocillopora damicornis, is reported during infection of a coral pathogen Vibrio coralliilyticus, wheras no effect on transcription is observed when P. damicornis is infected with a nonvirulent strain of V. coralliilyticus. 205 Furthermore, using RNA sequencing and targeted approaches, downregulation of other AMP expressions is observed in the host/pathogen infection model of P. damicornis and V. coralliilyticus. 206
Conclusions
AMPs are one of the most diverse antimicrobials produced by all forms of life for their survival in different competitive environments. The emergence of resistance against AMPs represents a major challenge to humanity in the present scenario. A detailed understanding on the mechanism of resistance development could shed light on the design of new therapeutics against AMP resistance (Table 1). Furthermore, upon dissecting the resistance mechanism, host defense AMPs such as defensins and cathelicidins could be used as potential compounds for host-directed therapies to treat drug-resistant human pathogens such as M. tuberculosis. 207 Although bacteria may coevolve for resistance development against AMPs, however, these resistance mechanisms are relatively nonspecific, which results in a negligible or low level of resistance.
Summary of Diverse Resistance Mechanisms for Antimicrobial Peptides
AMP, antimicrobial peptide; BraRS, bacitracin resistance-associated; LPS, lipopolysaccharide; PBP, penicillin-binding protein; SIC, streptococcal inhibitor of complement.
On the contrary, a diverse repertoire of AMPs may serve as reliable alternatives to conventional antibiotics.11,208–210 Specially, bacteriocins provide endless opportunities to obtain new AMPs as bacterial diversity is enormous. 211 AMPs are easy to modify and thus provide chances to improve their efficacy. 212 In addition, AMPs are highly efficient against complex biofilms and have their potential in in vivo animal models with less cytotoxicity.213,214 Considering all the abovementioned facts, many AMPs are being examined for anti-infective therapies in clinical settings.215–217
The use of AMPs as therapeutic agents or drugs should be carefully investigated for resistance development because the equivalent resistance mechanism is already present in producer strain. Next, it is worth to consider the perspective of the transfer of the resistance mechanism as AMP treatment delivers a high selective pressure for the development of resistance.18,218 However, further investigations are required to understand the mechanism of resistance development along with the coevolution of drug-resistant bacteria in humans in hospital settings.
Footnotes
Acknowledgments
The authors thank Juli for her help in the preparation of figures and stimulating discussions during the article preparation. They are also thankful to Gopinath Venugopal for his critical comments on the article.
Disclosure Statement
The authors declare no conflict of interest.
Funding Information
The present work is not supported by any funding source.