
Abstract
With the stagnancy of antibiotics development, polymyxins have become the last defense for treatment of multidrug-resistant (MDR) Gram-negative bacteria, whereas the effect of polymyxin monotherapy is limited by resistance. The objective of this study was to evaluate the effects of polymyxin B (PMNB)-vorinostat (SAHA) combination therapy against Gram-negative pathogens in vitro and in vivo. The antibacterial activities of PMNB and SAHA were evaluated by susceptibility testing. The synergistic effect was assessed by checkerboard tests and time-killing kinetics experiments. Cellular morphology studies and reactive oxygen species (ROS) assay were conducted to explore potential mechanisms. Also, Galleria mellonella models were made to evaluate the antibacterial effects in vivo. PMNB-SAHA had the synergistic effect against all tested isolates, reducing >2 log10 colony-forming units (CFU)/mL at 40 minutes, and showed more powerful antibacterial effects than PMNB alone in the 24-hour window. Cellular morphology study showed the change of membrane and disruption of integrity. ROS assay showed more oxidative stress in combination than PMNB or SAHA monotherapy. In animal models, PMNB-SAHA showed a higher survival rate than that of monotherapy. This study is the first to report the synergistic antibacterial effect of PMNB-SAHA therapy against MDR Gram-negative bacteria. Further clinical research is needed to confirm the results.
Introduction
With the rapid emergence and spread of drug resistance, Gram-negative pathogens with multidrug-resistant (MDR) have become a serious crisis for nosocomial infection and human health. 1 Treatments for infections by MDR pathogens, including MDR Enterobacteriaceae, Acinetobacter baumannii, and Pseudomonas aeruginosa, find it hard to achieve the desired therapeutic effect, causing higher mortality. For intensive care unit (ICU) patients, 70% of the infections were caused by Gram-negative pathogens. 2 Previous occupation of ICU rooms by patients with MDR Gram-negative bacterial strains is an independent risk factor for subsequent room occupants to be infected by these bacteria. 3 Some MDR bacterial isolates are even resistant to almost all antibiotics. With the lack of effective novel antibiotics, polymyxins have become the last line of antibiotic defense. 4
Polymyxins (A–E) are a type of cationic cyclic polypeptide antibiotics. As previously reported, polymyxin B (PMNB) and colistin (polymyxin E) have been used in clinical work. 5 However, PMNB and colistin share the same antibiotic mechanisms, as they compete with Mg2+ and Ca2+ in lipid A binding, destroy the outer membrane of bacteria, and give rise to cytoplasmic outflow and cell death. 6 Unfortunately, when using polymyxin monotherapy to treat Gram-negative bacteria, modification of lipid A and loss of lipopolysaccharides may occur, which increases the emergence of polymyxin resistance (PR). 7
Colistin is a class of inactive antibiotics. Colistimethate sodium injected intravenously is converted to the active version of colistin slowly and incompletely in vivo. 8 Comparatively, PMNB as the active version itself reaches effective blood concentration rapidly. 9 Previous studies also reported lower nephrotoxicity of PMNB compared with colistin. 10 Therefore, PMNB was chosen in our study.
Histone deacetylases (HDACs) can remove acetyl groups, causing chromatin condensation and gene silencing, and are associated with inhibition of transcription and regulation of cell signaling pathways generally.11,12 Vorinostat (suberoylanilide hydroxamic acid or SAHA) is a hydroxamic acid derivative that can inhibit zinc-dependent HDACs (class I, II, and IV).13,14 Several studies demonstrated that SAHA could induce DNA damage and produce oxidative lesions in many types of tumor.14–16 It has been approved by the U.S. Food and Drug Administration as an oral drug treating relapsed and refractory cutaneous T cell lymphoma (the recommended treatment protocol is 400 mg/day with food).17,18
In addition, recent reports indicated that antibacterial activity of polymyxins was enhanced for MDR Gram-negative bacilli via synergistic combination with the anthelmintic drug.19,20 Meanwhile, there is a strong correlation between rapid target polymyxins therapeutic concentration attainment and antibacterial activity and rapid emergence of PR bacteria. 21 Thus, to achieve the desired therapeutic effect at a lower drug concentration, a novel PMNB combination therapy is important and imperative. Previous research also showed that SAHA had effects against malaria and human immunodeficiency virus.22,23 However, the effects of SAHA on bacterial cells are unclear and may be related to enhance oxidative stress. 24 We hypothesized that SAHA can reduce the concentration of PMNB without lowering the therapeutic effect. Therefore, here, we investigated the potential synergistic antibacterial activity of PMNB and SAHA against MDR and polymyxin-resistant Gram-negative bacteria.
Materials and Methods
Bacterial isolates, drugs, and Galleria mellonella
The antibacterial activity of the PMNB and SAHA was tested through six Gram-negative bacterial strains. Among these isolates, Escherichia coli K-12 strain BW25113, Acinetobacter baumannii ATCC 17978, Pseudomonas aeruginosa PA14, and Klebsiella pneumoniae ATCC 43816 were wild-type standard strains. Acinetobacter baumannii HR13 and Klebsiella pneumoniae KP024, two clinical MDR wild-type pathogens, were isolated from ICU patients at a tertiary care hospital in Anhui, China. In addition, three Gram-positive bacteria were used to study the antibacterial activity of SAHA including wild-type standard strains Staphylococcus aureus RN450 and Enterococcus faecalis ATCC 29212, as well as one clinical wild-type isolate methicillin-resistant Staphylococcus aureus HR165. All isolates (from the Anhui Center for Surveillance of Bacterial Resistance, Hefei, China) were stored in cation-adjusted Muller-Hinton Broth (CAMHB) with 50% glycerol in cryovials at −80°C and grown in CAMHB or on broth agar at 37°C. PMNB was obtained from Sigma-Aldrich (St. Louis, MO). It was dissolved in ultrapure water and sterilized with a 0.22-μm cellulose acetate membrane filter (Solarbio Science and Technology Co., Ltd., Beijing, China). SAHA was ordered from Yuanye Bio-Technology Co., Ltd. (Shanghai, China) and dissolved in 99% dimethyl sulfoxide (DMSO; Sigma-Aldrich). All prepared solutions were stored at −20°C within 1 month. Galleria mellonella larvae (Kaide Ruixin Co., Ltd., Tianjin, China) in their final-instar stage were stored in the dark and used within 7 days from shipment.
Susceptibility testing
The minimum inhibitory concentrations (MICs) were measured by using the broth microdilution method for PMNB and SAHA in all strains. Overnight cultures were diluted 200-fold into fresh CAMHB, grown to log phase OD600 = 0.3 (the optical density of growth cultures was measured at 600 nm), about 5 × 108 CFU/mL, and finally diluted 1,000-fold into broth. A final bacterial inoculum of 5 × 105 CFU/mL was placed in each tube. PMNB and SAHA were added separately to the first tube and diluted serially to the last tube. The ranges of concentrations were 0.25–16 mg/L for PMNB, and 16–1,024 mg/L for SAHA. Tubes were read after overnight incubation at 37°C. The MICs were identified as the lowest drug concentrations that inhibit the visible growth of the bacteria. Susceptibility was determined by using Clinical and Laboratory Standards Institute (CLSI) breakpoints and the European Committee on Antimicrobial Susceptibility Testing (EUCAST).25,26 After overnight treatment or untreatment (controls), MICs of PMNB were used to evaluate the change of polymyxin susceptibility at 24 hours.
Synergistic effect testing
The synergistic effect was evaluated by the micro-dilution checkerboard assay. A single colony of bacteria was incubated overnight at constant rotation (5 × g) in CAMHB at 37°C. The overnight cultures were adjusted to about 108 CFU/mL and diluted by 1:1,000 with broth (5 × 105 CFU/mL) bacterial suspension. Ninety-six-well microtiter plates were used, and 100 μL of the 5 × 105 CFU/mL bacterial suspensions was prepared and mixed with drugs solution with the same final concentration, respectively, into each horizontal or vertical well. The ranges of selected concentration depended on previously tested MICs, which were 0, 1/16 × , 1/8 × , 1/4 × , 1/2 × , and 1 × MIC from low to high in each well for PMNB, as well as 0, 1/32 × , 1/16 × , 1/8 × , 1/4 × , 1/2 × , and 1 × MIC for SAHA. The microwell plates were incubated at 37°C for 24 hours before reading. The fractional inhibitory concentration index (FICI) was calculated to assess synergistic effect. The FICI was calculated by the equation (MIC of drugA in combination/MIC of drugA alone) + (MIC of drugB in combination/MIC of drugB alone) and defined as follows: FICI ≤0.5 means synergy; 0.5 <FICI ≤4.0 means indifference; FICI >4 means antagonism.27,28
Time-kill studies
Time-kill studies were conducted in all strains by using PMNB or SAHA alone and in combination, according to the method previously described. 29 Synergy is defined as a reduction of ≥2 log10 CFU/mL by using combined therapy compared with the most active monotherapy. 30 Bacterial strains were cultivated overnight in 5 mL CAMHB. The cultures were diluted to 1:200 with 10 mL of fresh broth and grown up to the OD600 value 0.25–0.3 (log-phase) with a shaker at 37°C. Then, they were diluted to a concentration of 5 × 105 CFU/mL and transferred to borosilicate glass tubes for each strain. PMNB (2 mg/L), SAHA (50 mg/L), or both compounds were added to achieve corresponding final concentrations. Samples and untreated cultures were removed at 0, 10, 20, 30, and 40 minutes and 2, 4, 8, and 24 hours after 10- and 105-fold dilutions with saline. Samples of bacterial cell suspension (1 mL) were aseptically placed on agar plates and incubated overnight at 37°C to measure the viable colony cells counts (CFU/mL). Controls (untreated) were set for each experiment.
Cellular morphology study
Scanning electron microscopy (SEM) was used for detecting the cell morphologic changes of polymyxin-susceptible Acinetobacter baumannii ATCC 17978 and polymyxin-resistant Acinetobacter baumannii HR13. As the previous time-kill studies, bacteria were treated with neither, one, or both of PMNB (2 mg/L) and SAHA (50 mg/L) for 2 hours in broth. Cultures were centrifuged at 4,600g for 10 minutes; the obtained supernatants were discarded; the bacterial pellets were resuspended in 1 mL of phosphate-buffered saline (PBS), and then they were washed three times in 1 mL PBS as described earlier. After that, the bacterial pellets were resuspended in 0.5 mL PBS with 2.5% glutaraldehyde and fixed for 2 hours at 37°C. Samples were observed and assessed at least 300 cells per assay on the SEM.
Reactive oxygen species assay
To measure intracellular reactive oxygen species (ROS) generation, the fluorescent probe carboxy-H2DCFDA [5(6)-carboxy-2’,7’-dichlorodihydrofluorescein diacetate] from Thermo Fisher (USA) was used. Carboxy-H2DCFDA entered A. baumannii cells, and it was converted into a membrane-impermeable cognate by cellular esterases or oxidized to a fluorescent form by superoxide, hydrogen peroxide (H2O2), or hydroxyl radicals. 31 The signal of fluorescence was analyzed by using a flow cytometer (Beckman Coulter CyAn ADP analyzer). Acinetobacter baumannii ATCC 17978 cells were grown with neither, one, or both PMNB (2 mg/L) and SAHA (50 mg/L) for 30 minutes; washed twice with precooled 1 × PBS to scavenge the agents; and concentrated by centrifugation (4,600g for 2 minutes). Samples were incubated with 5 μM carboxy-H2DCFDA for 10 minutes at 37°C and used to measure ROS accumulation in flow cytometry.
In addition, we also used the fluorescent reporter dye, 3’-(p-hydroxyphenyl) fluorescein (HPF; Thermo Fisher, USA) to further verify, which was oxidized by hydroxyl radicals with high specificity. Cells were treated with PMNB alone, SAHA alone, or PMNB-SAHA, or with 0.15% H2O2 at 37°C with gentle shaking for 30 minutes. After being washed twice with precooled 1 × PBS and concentrated by centrifugation, 5 μM HPF was added to treated or untreated cultures. Fluorescence values were measured by a BioTek Synergy HT Multi-Mode Plate Reader (BioTek, USA) with an excitation/emission setting of 490/515 nm. 32
To further determine that ROS is a key factor in lethality for the PMNB-SAHA combination. DMSO and thiourea, effective hydroxyl-radical scavengers, can prevent ROS accumulation, which were used to study ROS-dependent or -independent modes of killing. According to our previously described time-kill experiment, prepared samples (1 mL) were placed on agar plates containing 5% DMSO or thiourea at a final concentration of 600 mM, and they were used to study antibacterial effects in combination. In addition, the effect of growth kinetics was studied in DMSO or thiourea alone (Supplementary Data).33,34
G. mellonella killing-treatment assay
According to previous studies, 35 we built the G. mellonella infection model to evaluate the antibacterial activity of PMNB combined with SAHA in vivo. G. mellonella were stored at 15°C within 5 days from shipment. Sixteen larvae (250–350 mg in weight) were selected randomly and used in each group. Bacterial cells (Acinetobacter baumannii ATCC 17978 and HR13) were washed twice in normal saline (NS; 0.9% saline) and then diluted to OD600 0.35. Bacterial cells were injected into the hemocoels of each larva through the last left proleg by a 50-μL Hamilton syringe (Hamilton Company, Shanghai, China). Each group of larvae was injected with 10 μL of bacterial suspensions containing ∼105 CFU/larva of bacteria in NS. After injection, larvae were incubated at 37°C. Larvae were considered dead if they did not respond to touch, and the number of dead larvae was recorded daily for 6 days. G. mellonella larvae were infected with a lethal dose of A. baumannii, and drugs (PMNB [2.5 mg/kg] or SAHA [8 mg/kg] alone and in combination) were inoculated into a different proleg within 20 minutes. Drug doses were based on those used for humans. Two control groups (NS injection group and no injection group) were set for each experiment.
Statistical analysis
All statistical analyses were performed with SPSS 21.0. One-way analysis of variance with post hoc test was used to calculate the change of antibacterial activity, cell length, and fluorescence values in each drug alone or in combination. Survival curves using Kaplan–Meier methods were drawn and compared with the log-rank test to compare the differences between groups. A p-value of <0.05 indicates that the difference is significant.
Results
Antimicrobial susceptibility of PMNB and SAHA
For all Gram-negative strains, MICs of PMNB and SAHA are shown in Table 1. According to the CLSI and EUCAST, MDR clinical isolates (Acinetobacter baumannii HR13 and Klebsiella pneumoniae KP024) were resistant to PMNB, whereas others were susceptible in this study. SAHA alone had antibacterial activity at a concentration of up to 512 μg/mL for all Gram-negative isolates, which is higher than Gram-positive bacteria (Supplementary Table S1).
Minimum Inhibitory Concentrations of Polymyxin B and Vorinostat
MIC, minimum inhibitory concentration; PMNB, polymyxin B; SAHA, vorinostat.
The changes to the PMNB MICs after overnight treatments are shown in Table 2. For the controls and combination group, MICs of PMNB did not change (within twofold of initial MICs) at 24 hours. With the PMNB alone, MICs of PMNB remained unchanged for polymyxin-resistant isolates whereas they increased remarkably more than eight times for polymyxin-susceptible isolates at 24 hours.
Treatment of Polymyxin B With or Without Vorinostat Impact on Minimum Inhibitory Concentrations at 24 Hours
The values of PMNB MICs are shown in brackets.
Controls, no treatment.
Micro-dilution checkerboard assays
We evaluated synergistic interaction (FICI <0.5) between PMNB and SAHA by calculating the FICIs (Fig. 1). For MDR and polymyxin-resistant clinical isolates, the PMNB-SAHA combination reduced the MIC of PMNB effectively from 4 to 0.5 μg/mL and of SAHA to 64 μg/mL against Acinetobacter baumannii HR13 (Fig. 1D). For Klebsiella pneumoniae KP024, the combination reduced the MIC of PMNB from 8 to 1 μg/mL and of SAHA to 32 μg/mL (Fig. 1F). This indicates that SAHA is a potential companion of polymyxin in treating infections of MDR and polymyxin-resistant Gram-negative pathogens.

The inhibitory concentration is shown in a combination of PMNB and SAHA.
Time-kill studies
The time-killing kinetics of PMNB (2 mg/L) or SAHA (50 mg/L) alone and the combination for all selected isolates are shown in Figs. 2 and 3. Against the four polymyxin-susceptible strains, PMNB monotherapy showed rapid bactericidal effect with ∼2.5 log10 CFU/mL killing for Escherichia coli BW25113 (Fig. 2A), 2.8 log10 CFU/mL killing for Pseudomonas aeruginosa PA14 (Fig. 2B), 2.4 log10 CFU/mL killing for Acinetobacter baumannii ATCC 17978 (Fig. 2C), and 2.5 log10 CFU/mL killing for Klebsiella pneumoniae ATCC 43816 (Fig. 2E) within 40 minutes. However, bacterial counts returned within 24 hours (Fig. 3). With the combination, there were nearly 5 log10 CFU/mL of bacterial killing within 40 minutes (Fig. 2) and they maintained stable antibacterial activity for up to 24 hours (Fig. 3). The bacterial counts of combination were reduced >2 log10 CFU/mL more than that of PMNB monotherapy (Fig. 2). In addition, bacterial killing of SAHA monotherapy was not significant compared with the growth control group (p > 0.05) (Figs. 2 and 3).
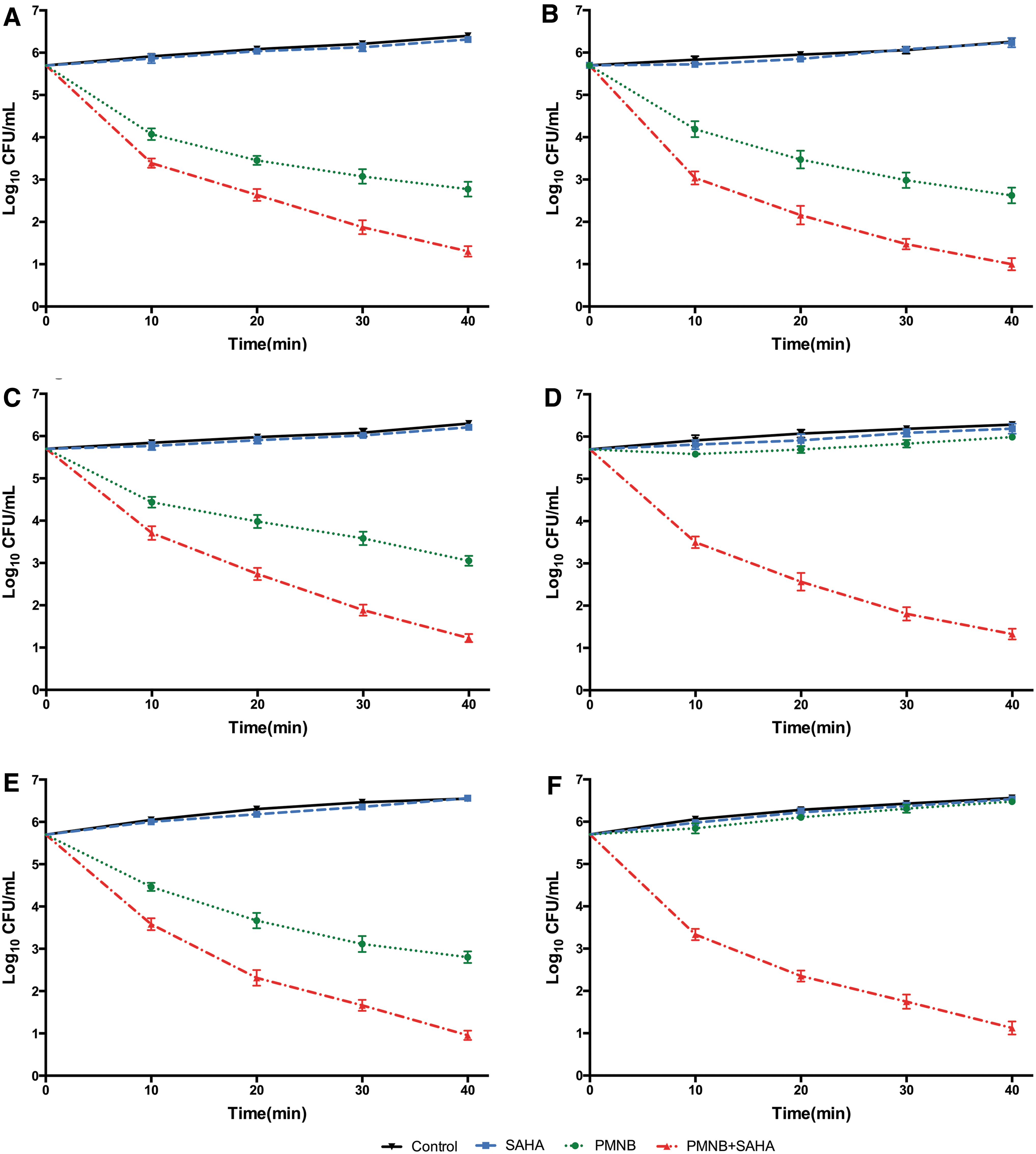
Synergistic effect of PMNB (2 mg/L) combined with SAHA (50 mg/L) against Gram-negative bacterial strains in 40 minutes.
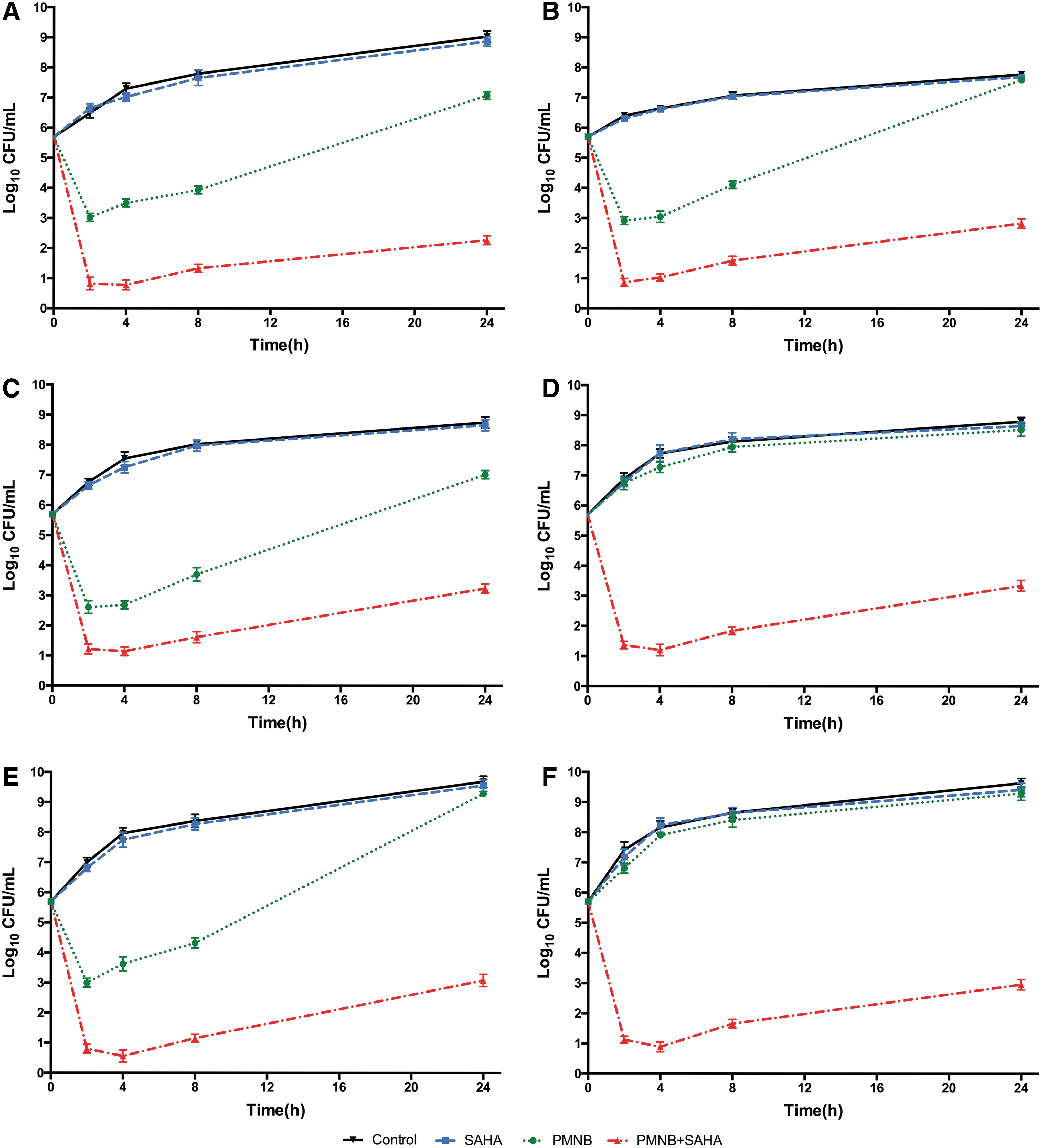
Twenty-four hours-kill kinetics of PMNB and SAHA monotherapy and combination therapy against Gram-negative bacterial strains.
Against the polymyxin-resistant isolates, Acinetobacter baumannii HR13 (Fig. 2D) and Klebsiella pneumoniae KP024 (Fig. 2F), monontherapy with PMNB reduced far <1 log10 CFU/mL of bacterial counts (Fig. 3D, F). However, synergistic activity was observed in the combination therapy within 40 minutes, with almost 5 log10 CFU/mL of bacteria reduced rapidly (Fig. 2D, F) and no regrowth at 24 hours for Acinetobacter baumannii HR13 (Fig. 3D) and Klebsiella pneumoniae KP024 (Fig. 3F).
Cellular morphology studies
The SEM images of polymyxin-susceptible Acinetobacter baumannii ATCC 17978 and polymyxin-resistant Acinetobacter baumannii HR13 incubated with PMNB, SAHA, or both are shown in Fig. 4. The SEM results revealed that the cell membrane surfaces of bacteria were smooth, and the average cell length remained ∼2 μm in the untreated group (Fig. 4A, E). For Acinetobacter baumannii ATCC 17978, PMNB monotherapy led to visible shortening of the cells (p < 0.01) and the surface of the cell membrane became rough and shrunk (Fig. 4B). Conversely, no significant effects were found in bacterial size (p > 0.05) and membrane surface for Acinetobacter baumannii HR13 (Fig. 4F). SAHA alone had minimal impacts on the surface of bacterial cell membrane and cell length (p > 0.05) for both (Fig. 4C, G). With the PMNB-SAHA combination, bacterial cell length was shortened (p < 0.01), but the bacterial surface was smoother than PMNB monotherapy (Fig. 4D, H).

Impact of PMNB and SAHA treatment on the morphology of polymyxin-susceptible and polymyxin-resistant Acinetobacter baumannii. Images from SEM for polymyxin-susceptible Acinetobacter baumannii ATCC 17978
ROS assays
There was a need to confirm whether treatment of the PMNB-SAHA combination induced ROS production. The ROS levels of Acinetobacter baumannii ATCC 17978 are shown in Fig. 5. For the carboxy-H2DCFDA, SAHA monotherapy group had almost no change of fluorescent intensity compared with the control group and PMNB monotherapy showed relatively low production of ROS, whereas the combination group showed notable ROS accumulation (Fig. 5A). For the HPF, we treated cultures with H2O2 as a positive control, which was previously demonstrated to induce hydroxyl radicals. 36 We observed an about threefold increase in fluorescence, indicating an increase in ROS production (p < 0.001). We next assayed the A. baumannii cultures treated with PMNB alone and observed that monotherapy induced an about twofold increase in fluorescence compared with the untreated controls (p < 0.01). SAHA alone did not alter the level of fluorescence (p > 0.05). However, when treated with PMNB-SAHA, we observed that the combination induced an about twofold increase in fluorescence values compared with PMNB alone (p < 0.001).
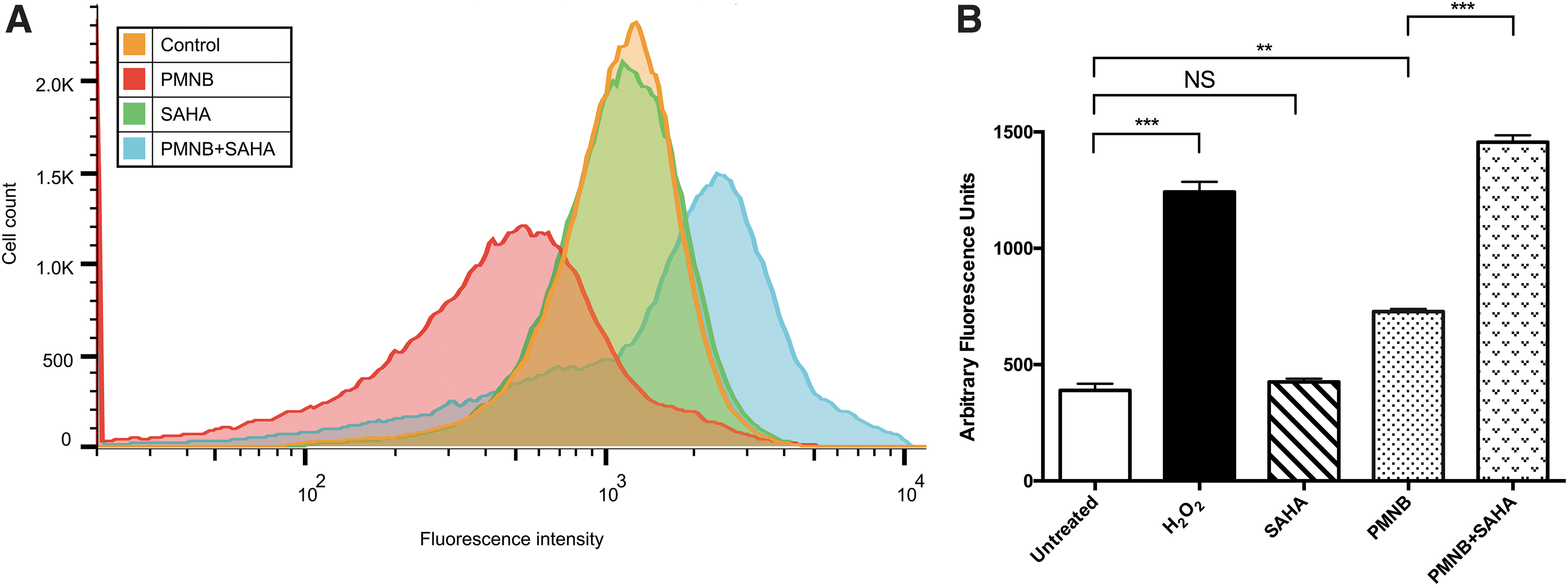
The generation of ROS in PMNB and SAHA for Acinetobacter baumannii ATCC 17978.
Then, we sought to determine whether ROS production was required for the rapid killing. Therefore, we utilized the time-killing kinetics experiments (Fig. 6). After the addition of 5% DMSO, the protection was found in PMNB monotherapy (reduced by 1.7 log10 CFU/mL; p < 0.05) and combination group (reduced by 3 log10 CFU/mL; p < 0.01) compared with no DMSO added (Fig. 6A). The bactericidal effect of PMNB-SAHA combination was reduced to about 1/1,000 after 5% DMSO was added. Likewise, when we detected the presence of thiourea, we found a striking decrease in the ability of PMNB-SAHA combination to kill A. baumannii compared with the combination without thiourea (reduced by 1.9 log10 CFU/mL; p < 0.01). PMNB with thiourea was reduced by 0.8 log10 CFU/mL compared with without thiourea (p < 0.05) (Fig. 6B). Obviously, treatment with DMSO or thiourea markedly inhibited the rapid decrease in antibacterial activity. No protection was observed in SAHA alone with DMSO or thiourea (p > 0.05). The addition of DMSO or thiourea alone did not change the growth kinetics (Supplementary Fig. S1).

ROS accumulation was inhibited in time-killing curves for Acinetobacter baumannii ATCC 17978. As the hydroxyl-radical scavengers.
G. mellonella killing-treatment model
In the infection models of Acinetobacter baumannii ATCC 17978 and HR13, the survival rate of G. mellonella with treatment of PMNB alone was 62.5% (Fig. 7A) and 31.25% (Fig. 7B) at 96 hours after inoculation. However, when SAHA was used as treatment, survival rates were almost identical to the untreated controls, which was consistent with the results in vitro. When the PMNB-SAHA combination was used, it protected the G. mellonella from infection of lethal bacteria more effectively than PMNB monotherapy did (p < 0.001), as 93.75% (Fig. 7A) and 87.5% (Fig. 7B) of G. mellonella were still alive in ATCC 17978 and HR13 infection models.
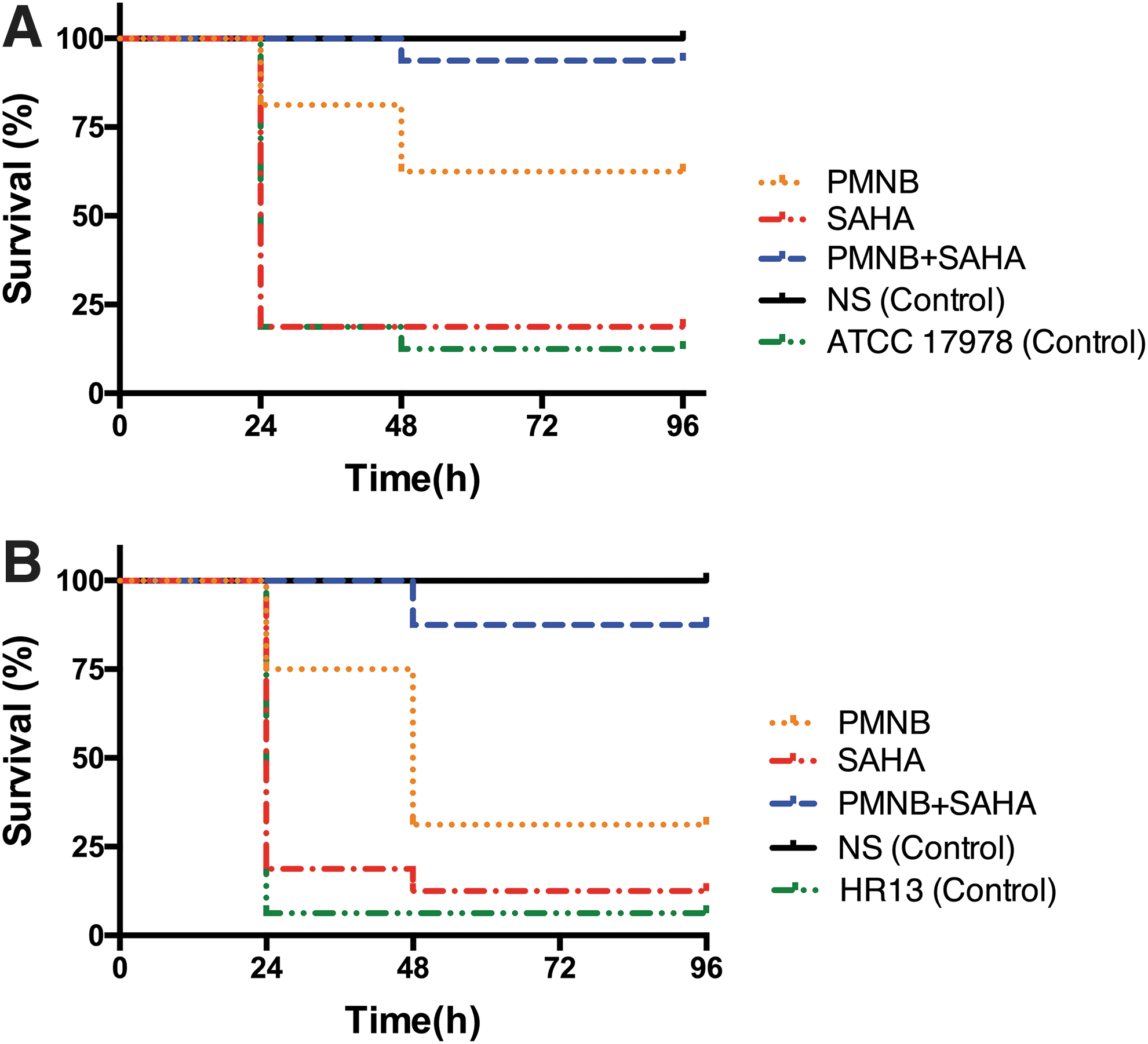
Effect of combined therapy with PMNB and SAHA on the Galleria mellonella infective model.
Discussion
The infections of MDR Gram-negative bacteria are becoming a public health concern. However, the development of new antibiotics has been limited over the recent decades. In addition, polymyxin monotherapy can transform primary infection into polymyxin-resistant bacterial infection, leading to more health care burden and higher mortality rates. Thus, novel treatment strategies are urgently needed. SAHA, as a derivative of hydroxamic acid, is used to treat patients with refractory and relapsed cutaneous T cell lymphoma. 17 The possible mechanisms of SAHA against proliferation include HDAC inhibition and DNA damage. 16 However, the mechanism of potential antibacterial effects still remains uncertain. In this study, we investigated the effects and mechanisms of PMNB-SAHA combination on Gram-negative infections.
In our study, SAHA alone showed no antibacterial activity for all Gram-negative isolates (compared with the controls, p > 0.05). PMNB showed certain antibacterial activity in a short time and gradually reduced in the next few hours. Conversely, PMNB-SAHA combination enhanced this certain antibacterial activity (∼5 log10 CFU/mL) and inhibited the growth of bacteria within 24 hours. Also, PMNB monotherapy reduced the susceptibility for PMNB and thus increased the MICs of originally polymyxin-susceptible isolates, which could explain the regrowth of bacteria observed at 24 hours, and this may account for the clinical failures. However, the combination therapy prevented the reduction of polymyxin susceptibility and showed a long-lasting antibacterial effect. In addition, the MICs of SAHA to Gram-positive isolates were lower than Gram-negative stains, which showed that the potential activity of SAHA for Gram-positive bacteria is worth studying.
For the SEM images, PMNB monotherapy caused a rough and shrunken cell membrane surface on Acinetobacter baumannii ATCC 17978, which indicated that the stability and integrity of the cell membrane was destroyed. However, there was no visible change in Acinetobacter baumannii HR13. It is possible that the PMNB resistance in Acinetobacter baumannii HR13 altered their membrane surface interaction. The combination therapy showed that the outer membrane surface remained smooth on the shortened bacterial cells. It suggested that the combination therapy prevented disruption of the cell membrane integrity and stability, which could be a change in the mechanism of action of PMNB.
As the results of ROS assay, when using carboxy-H2DCFDA, PMNB monotherapy showed an obvious reduction of ROS production compared with the control group. As previous research demonstrated, polymyxins can permeabilize the outer membrane of Gram-negative bacteria and destabilize it. 37 This may cause the intracellular substance to flow out, so the fluorescence intensity was not completely detected in the carboxy-H2DCFDA, which explains the reduction of ROS detected. In the combination, bacterial fluorescence intensity obviously increased for both, and the HPF fluorescence values were remarkably stronger than the sum of the fluorescence levels of PMNB and SAHA alone. These indicate that SAHA can increase production of ROS. However, fluorescence intensity of the SAHA monotherapy group was no change for the control group, and there was no difference between SAHA monotherapy with and without DMSO or thiourea. This may be because SAHA alone cannot pass through the outer membrane of Gram-negative bacteria, and thus cannot induce the increase ROS production. Also, compared with the combination without DMSO group, the antibacterial activity was significantly inhibited after adding ROS inhibitors. This means that DMSO and thiourea cleared the production of oxidative stress caused by SAHA alone, which is the underlying main mechanism for PMNB-SAHA combination.
Hence, combining our results with previous morphological studies, we supposed that PMNB destroys the integrity of the outer membrane, letting SAHA into the bacterial cell, which causes oxidative stress and enhances the generation of ROS. Interestingly, the HPF fluorescence values of PMNB alone significantly increased (p < 0.01), and antibacterial activity was also reduced after ROS inhibitors were added. These indicate that an increase of ROS production may also be a potential bactericidal mechanism of PMNB, which was consistent with previous reports.34,36 SAHA enhances the production of ROS, which is accompanied by the hyperpolarization of mitochondrial membrane, 38 and causes the outflow of cations to the cytoplasm, such as Mg2+ and Ca2+. It also inhibits the electrostatic interaction between the positively charged (PMNB) and the negatively charged lipid A, and therefore, reduces the extent of disruption of the cell membrane integrity and stability. This may also explain why the cell membrane of the combination therapy group was smoother than that of the PMNB monotherapy group. Thus, in the combination group, the main bactericidal mechanism of PMNB may be production of ROS as well as disrupting the cell membranes to some extent and thus letting SAHA in and exhibiting synergistic antibacterial activity.
Recently, G. mellonella has been used as a well-accepted invertebrate insect model to study the effects of antimicrobial combination in vivo. The main advantages of this model include the low cost of maintenance, the fast life cycle, the possibility of using a large number of caterpillars, and the innate immune system, which are evolutionarily conserved relative to mammals. We investigated PMNB or SAHA alone and in combination in the infection model of Acinetobacter baumannii ATCC 17978 and HR13. SAHA monotherapy showed no effects in both infection models (p > 0.05). PMNB monotherapy showed some effects, whereas PMNB-SAHA combination was significantly more effective (p < 0.001), especially in the polymyxin-resistant infection model (HR13). This was consistent with our in vitro results that provided preliminary evidence in vivo. This combination could be a useful promotion of existing therapeutic methods.
Overall, this study is the first to report the strong and constant bactericidal effects of PMNB-SAHA combination against MDR Gram-negative bacteria. However, we should acknowledge some limitations. First, we chose MDR, a polymyxin-resistant A. baumannii strain, and a K. pneumoniae strain, which were reported as a top bacterial “superbug” by the World Health Organization for research.39,40 There still remains other polymyxin-resistant Gram-negative isolates, such as P. aeruginosa and E. coli, which could also be tested to further confirm our results. Second, the concentration of SAHA (50 mg/mL) we used in vitro was higher than the blood concentration of clinical treatment and changes in pharmacokinetics of the combination are unclear. Then, we used G. mellonella models to test the effects of drug combination at clinically treated concentrations in vivo; however, more effective mammal models are further needed to validate the results at clinically relevant blood concentrations of SAHA and explore other possible mechanisms of combination as well as the effect of pharmacokinetics in vivo when combined therapy is used.
With the stagnant development of new antibacterial drugs, this study gives hope for using novel drug combinations to treat MDR Gram-negative bacteria. Further clinical studies are needed to confirm the therapeutic effects of PMNB-SAHA combination.
Conclusion
PMNB-SAHA combination, as a novel therapy, can increase antibacterial activity of PMNB against MDR Gram-negative pathogens mainly via the generation of ROS and reduce the emergence of PR. Further clinical research is needed to confirm its efficacy.
Footnotes
Acknowledgments
The authors wish to thank all the students in the Institute of Bacterium Resistance of Anhui Medical University (Hefei, China) for their helpful assistance.
Ethical Statement
This study was approved by the Ethical Committee of the Second Hospital of Anhui Medical University (Hefei, China) and received Joint Commission International certification (approval no.: PJYX2018-001).
Disclosure Statement
The author reports no conflicts of interest in this work.
Funding Information
Funding for this work is supported by grant 81673242 from the National Natural Science Foundation of China.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.