
Abstract
Background:
Klebsiella pneumoniae is currently considered as an immediate threat to human health due to its various multidrug efflux pumps. Microbially synthesized silver nanoparticles (AgNPs) are an attractive and eco-friendly approach to prevent antibiotic resistance in bacteria. In the present study, we compared the inhibitory effect of both commercial and green AgNPs by Bifidobacterium bifidum on OxqAB efflux pump genes in ciprofloxacin-resistant strains of K. pneumoniae.
Materials and Methods:
AgNPs were characterized by ultraviolet-visible spectrophotometer, Fourier transform infrared spectroscopy, X-ray diffraction, zeta potential, transmission electron microscopy, and scanning electron microscopy. Antibiogram was used to identify resistant isolates and the effect of the biosynthesized AgNPs against OxqAB efflux pump strains was assessed by the minimum inhibitory concentration (MIC) method. The expression levels of oxqAB genes were evaluated using real-time polymerase chain reaction (PCR) followed by exposure to subMICs of the AgNPs.
Results:
PCR results showed that 25 strains had OxqAB efflux pump and the MIC method indicated that AgNPs had an inhibitory effect on all resistant strains with OxqAB efflux pump. The efficacy of the synthetic nanoparticles was assessed by comparing the antiefflux pump activity with commercial AgNPs. In ciprofloxacin-resistant isolates, the oxqAB genes expression levels reduced in the subMIC of both AgNPs, whereas biosynthesized AgNPs had greater bactericidal effects compared with the commercial AgNPs.
Conclusions:
Efflux pumps could be an attractive target for our biosynthesized AgNPs. The oxqAB genes expression levels reduced in subMIC of both AgNPs, whereas biosynthesized AgNPs had greater bactericidal effects than the commercial AgNPs.
Introduction
Biosynthesized nanoparticles are fundamental building blocks of nanotechnology. 1 Nanotechnology is based on controlling empirical processes for the synthesis of nanoparticles in different sizes and shapes. Microbially mediated synthesis of nanoparticles is cost-effective, nontoxic, and eco-friendly with important advantages over synthetic methods,2,3 due to the controlled size and shape of nanoparticles. 4
Silver is a nontoxic and reliable inorganic antimicrobial agent that has been used for centuries with the potential to destruct various types of pathogenic organisms. 5 Silver has been known as a potent antibacterial, antifungal, and antiviral agent; thus, it can be used to develop medical devices and therapeutic agents. 5 Silver nanoparticles (AgNPs) can be produced by many bacteria through both intracellular and extracellular processes. Various studies have indicated that several lactic acid bacteria, 6 psychrophilic bacteria, bacillus strains, as well as Escherichia coli and Bifidobacterium strains could reduce silver ions to AgNPs.7,8
The emergence of antibiotic-resistant infections is a serious concern worldwide. 9 Ciprofloxacin-resistant strains typically exhibit crossresistance to multiple antibiotics, including ceftriaxone, primarily due to the presence of PMQR gene and blaCTX-M-65, blaCTX-M-55 blaCMY-2, and blaCMY-72 genes. 10 Extended-spectrum β-lactamase produced by Klebsiella species can make them potentially resistant to virtually all beta-lactam antibiotics, except for carbapenems and aminoglycosides, fluoroquinolones, tetracyclines, chloramphenicol, and trimethoprim/sulfamethoxazole.10,11
Multidrug-resistant (MDR) Klebsiella pneumoniae is one of the most intensively investigated efflux systems. 12 Overexpression of the efflux pump is the pivotal mechanism of resistance caused by K. pneumoniae. 13 One of the most intensively investigated efflux systems in K. pneumoniae and other members of the Enterobacteriaceae family is the oqxAB operon considering antibiotic resistance. This operon encodes an efflux pump conferring resistance to olaquindox, chloramphenicol, and quinolones. 14 It has shown that the two genes flanking the oqxAB operon in the chromosome of K. pneumoniae are rarA, encoding an AraC-type transcriptional activator, and oqxR, encoding a GntR-type transcriptional repressor, where rarA upregulates and oqxR downregulates the expression of the OqxAB efflux pump operon.15,16 This study was done to compare the effect of both commercially and microbially synthesized AgNPs using Bifidobacterium bifidum on the expression of the efflux pump gene, oqxAB, in clinical isolates of antibiotic-resistant K. pneumoniae.
Materials and Methods
Microbially synthesized AgNPs
B. bifidum PTCC1644 strain was purchased from the National Iranian Research Organization for Science and Technology (IROST), Iran, and inoculated in 100 mL sterile DeMan Rogosa Sharpe broth supplemented with 0.5%
Commercial AgNPs
AgNPs were purchased from the Nanosany Co. (Mashhad, Iran) with the average particle size of 20 nm.
Characterization of AgNPs
AgNPs were characterized using ultraviolet-visible (UV-Vis) spectroscopy, Fourier transform infrared spectroscopy (FTIR), dynamic light scattering (DLS), zeta potential, transmission electron microscopy (TEM), 17 and scanning electron microscopy (SEM). 18 The reduction of silver ions was monitored by measuring the absorbance of the reaction mixture at the wavelengths of 250–700 nm using a UV-Vis spectrophotometer (Bio-Tek, Winooski, VA) and particle size distribution measurements were performed by a DLS (Malvern Instrument Ltd., Worcestershire, United Kingdom). The crystallographic nature of the prepared AgNPs was evaluated using an X-ray diffractometer instrument (XRD, PW3710; Almelo, the Netherlands) in the range of 2θ with Cu Kα radiation at 40 kV and 30 mA. The FTIR spectrum was recorded using PerkinElmer Spectrum RX-IFTIR spectrophotometer in the range of 400–4,000 cm−1 (PerkinElmer RX1, Spectrum Two Germany). Morphology, topography, and size of the synthesized nanoparticle were identified under SEM 18 and TEM. 17 Images were recorded using a Carl Zeiss Meditec AG (Jena, Germany).
Sample collection and characterization
From April 2017 to April 2018, 100 K. pneumoniae isolates were collected from the patients at Tehran hospitals. All bacterial isolates were identified as K. pneumoniae in the clinical laboratory by nonautomated biochemical tests according to the protocols suggested by MacFaddin. Furthermore, antibiotic susceptibility of the K. pneumoniae isolates to imipenem (10 μg), ciprofloxacin (5 μg), tetracycline (30 μg), gentamycin (20 μg), amikacin (30 μg), cefoxitin (30 μg), and ceftazidime (30 μg) was also assessed using the disk diffusion method to determine the drug-resistant isolates; the results were interpreted as sensitive, intermediate, or resistant according to the inhibitory zone diameters around the disks based on the Clinical and Laboratory Standards Institute (CLSI) guidelines. It should be noted that in all experiments, K. pneumoniae ATCC13884 was used as a standard sample. 6
Susceptibility determination
After identifying ciprofloxacin-resistant strains, the minimum inhibitory concentration (MIC) was determined by the dilution method in 96-well plates for both biosynthesized and commercial AgNPs. The experiment was performed in triplicate and the presence of oqxAB genes in K. pneumoniae isolates was analyzed by polymerase chain reaction (PCR) using the primers described in Supplementary Table S1. Genomic DNA was extracted from bacteria, using the Bacteria Genomic DNA Extraction Kit (Sinaclon Co., Iran) according to the manufacturer's protocol. The PCR reaction mixture with the final volume of 20 μL was prepared for each sample (including positive and negative controls) and DNA amplification was performed in a thermal cycler (Eppendorf Master Cycler Gradient, Germany).
Antibacterial assay
MIC-based assay in accordance with CLSI was performed by the dilution method in 96-well plates for ciprofloxacin. The antibacterial activity of biosynthesized AgNPs from Bifidobacterium bifidium was tested against pathogenic bacteria by the microdiffusion method. The AgNPs were poured into the 96-well dilution plates at the concentrations of 1.95–500 μg/mL and transferred to the Muller Hinton Broth (Merck Co., Germany) culture medium to achieve a final volume of 195 μL. Then, 5 μL of the ciprofloxacin-resistant strain culture equal to a 0.5 McFarland standard was added to all well plates to achieve a final inoculum of 200 μL. The results were determined by visual turbidity. MIC was recorded as the lowest dilution indicating no bacterial growth. Free bacterial AgNPs and AgNPs containing K. pneumoniae (strain ATCC700603) were used as negative and positive controls, respectively.
Treatment of the efflux pump inhibitor
To determine the role of efflux pump in the ciprofloxacin-resistant phenotypes in K. pneumoniae isolates, ciprofloxacin MIC was assessed in the presence of 12.5 μm efflux pump inhibitor carbonyl cyanide 3-chlorophenylhydrazone (CCCP). The CCCP was added to each well containing 1.5–384 μg/mL ciprofloxacin. MICs were redetermined then and compared with and without CCCP. One of the wells containing CCCP and without ciprofloxacin was used as control. A decrease in ciprofloxacin MIC after adding CCCP was considered as a positive criterion for the presence of an efflux pump in K. pneumoniae isolates.
RNA extraction and complementary DNA production
After treating the ciprofloxacin-resistant K. pneumoniae strain with AgNPs at sub-MIC concentration, it was incubated in Muller Hinton Broth at 37°C overnight. Subsequently, after overnight incubation, bacteria were harvested by centrifugation at 10,000 g for 2 min, and RNA extraction was performed using RNX-Plus Solution (Sinaclon Co.) according to the manufacturer's instructions. RNA quantity and quality were evaluated using NanoDrop and agarose gel electrophoresis methods. DNase treatment was applied to remove the remaining genomic DNA from the extract (Fermentas, Lithuania) and the complete removal of contaminated DNA was confirmed by PCR. Reverse transcription was carried out using the Revert Aid™ First-Strand cDNA Synthesis Kit (Fermentas).
Real-time PCR
To study the alteration of expression of the oqxAB genes in K. pneumoniae isolates, we carried out real-time PCR (RT-PCR) using specific primers described in the Supplementary Table S1. RT-PCR was performed in triplicate using a GreenStar Master Mix (Amplicon, Denmark) on a StepOne ABI real-time PCR equipment (Applied Biosystems, Foster City, CA). Then, 100 ng of complementary DNA (cDNA) was used as a template in 25-μL reactions, including 2 × GreenStar Master Mix (12.5 μL), forward primer (5 pmol/μL), reverse primer (5 pmol/μL), and diethyl pyrocarbonate water (9.5 μL). The quantitative PCR cycling was performed at 95°C for 10 min, followed by 40 cycles at 95°C for 20 sec, and 40 sec at 61°C, and eventually, a melting stage to determine the unspecific PCR product or possible primer dimers. Triplets of the negative control (resistant strain without AgNPs) were included in all qPCR runs, and the expression stability of the gmk gene (guanylate kinase) was used as the internal control.
Statistical analysis
Data were presented as mean ± standard error
18
of at least three tests done in duplicates. The relative expression of oqxAB efflux pump genes was determined using ΔΔC
Results
Synthesis of AgNPs
Visual identification
Bacterial biomass was mixed with silver nitrate solution (pH = 7.5) and incubated at 37°C for 24 hr. A change in the color of the mixture (yellow to dark brown) indicated excellent biosynthesis of the AgNPs (Fig. 1).

Visual identification of biosynthesized AgNPs. AgNPs, silver nanoparticles.
UV-Vis spectrophotometer
UV-Vis absorbance was used for the primary characterization of the synthesized AgNPs. Figure 2 illustrates the absorbance spectra of the reaction mixture containing an aqueous solution of 0.009 g silver nitrate and the culture supernatant of B. bifidum after incubation. The absorption spectrum indicated that the peak centered at 428 nm, corresponding to the surface plasmon resonance.
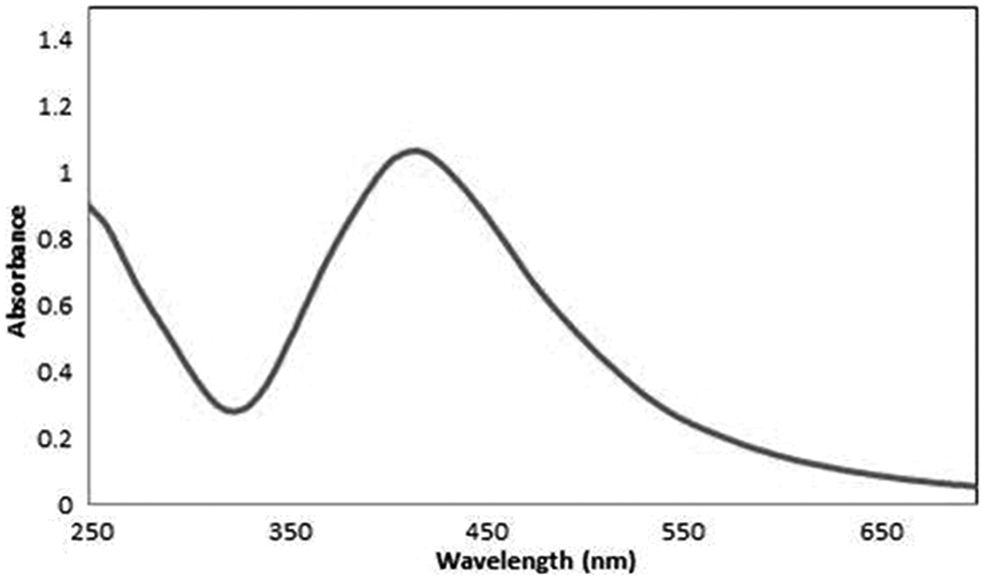
UV-visible absorbance spectra obtained from AgNPs. UV, ultraviolet.
Energy-dispersive spectroscopy and zeta analysis
The elemental composition of AgNPs with reduced ions was determined by energy-dispersive spectroscopy (EDX). The EDX profiles of the microbial-synthesized AgNPs indicated the presence of strong peaks for elemental silver along with weak oxygen and nitrogen peaks (Fig. 3). The silver nanocrystallites displayed an optical absorption peak between 2 and 4 keV, typical for metallic silver nanocrystallites due to their surface plasmon resonance. Four main characteristic diffraction peaks for Ag were observed at 2θ = 38.2, 46.2, 64.5, and 77.4 in the XRD test, which corresponded to the 111, 200, 220, and 311 crystallographic planes of face-centered cubic Ag crystals, respectively (Fig. 4).
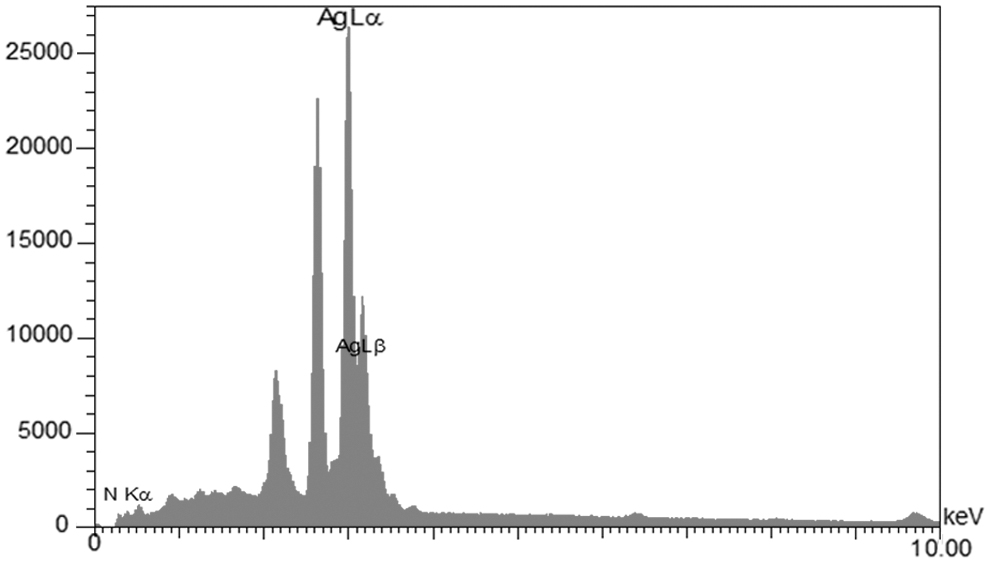
EDX analysis of biosynthesized AgNPs. EDX, energy-dispersive spectroscopy.
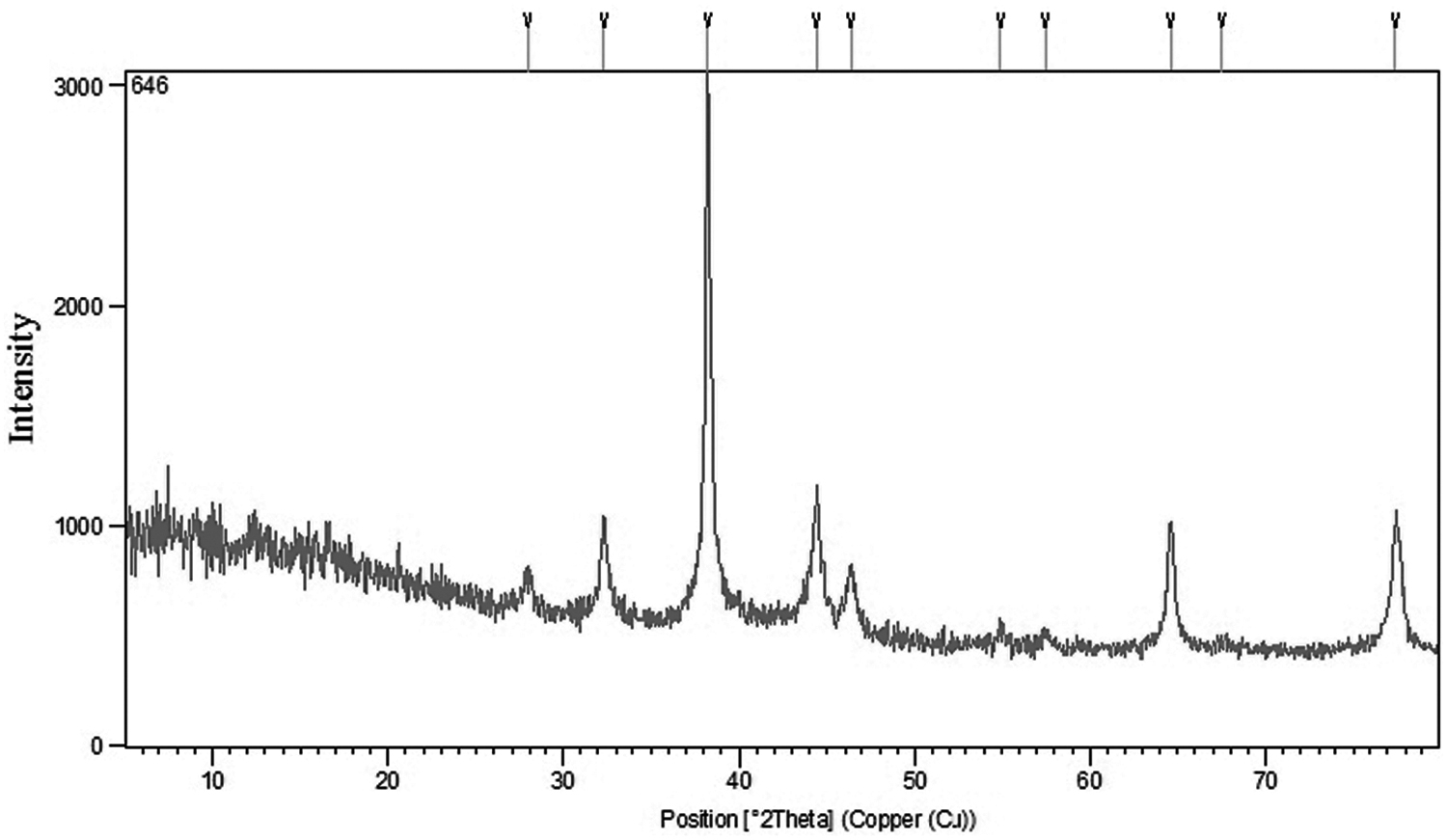
XRD analysis of the biologically biosynthesized AgNPs by Bifidobacterium bifidum under optimal conditions. XRD, X-ray diffraction.
Additionally, the zeta potential value was observed at −43.2 mV with zeta potential values outside the range of +30 to −30 mV (Fig. 5), which is indicative of the particle surface charge. The negative value indicated the stability of the nanoparticles and evaded the agglomeration of nanoparticles. This finding confirms the synthesis of AgNPs from B. bifidum.
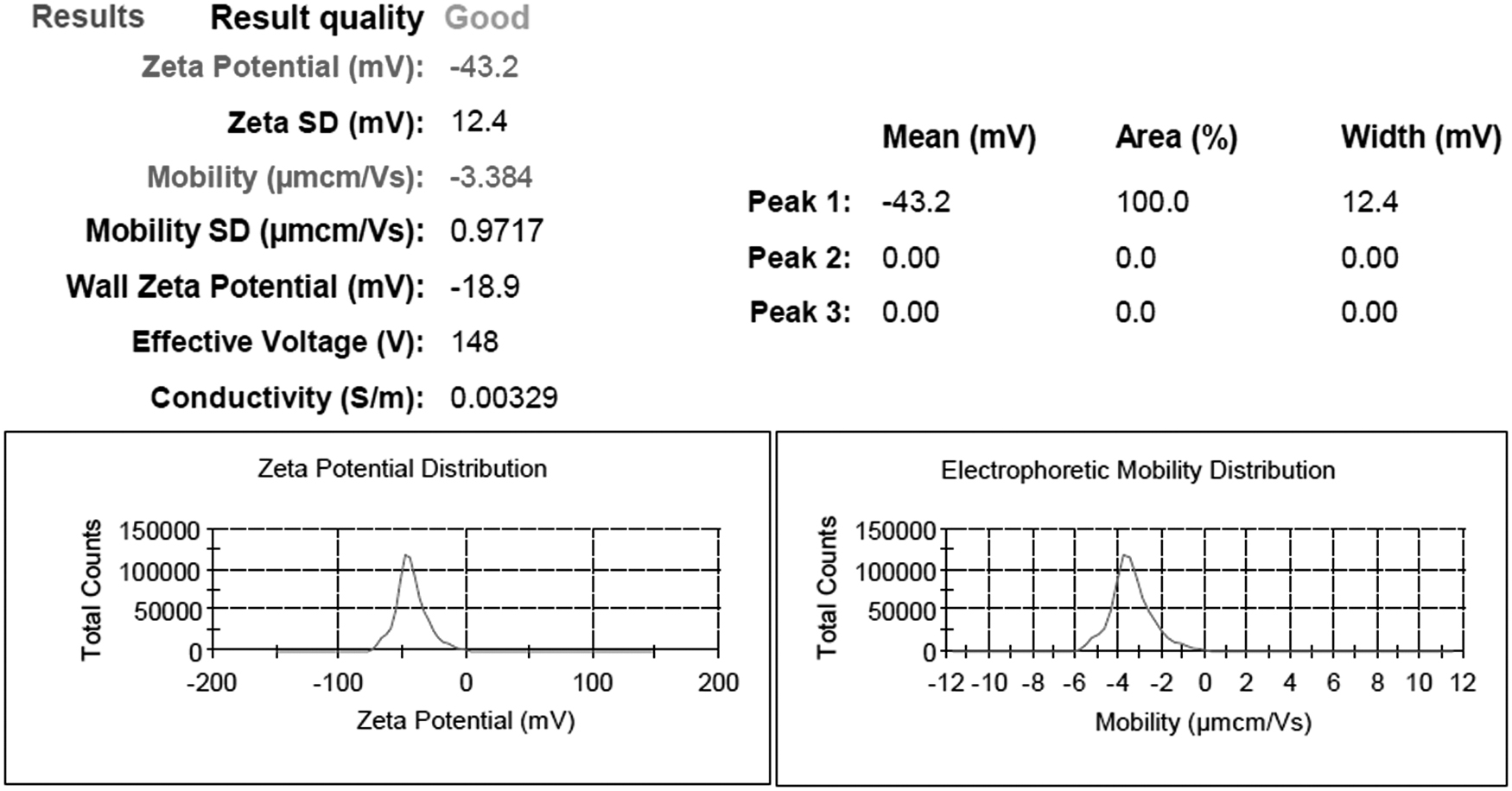
Zeta potential analysis of biosynthesized AgNPs.
SEM and TEM
To better clarify the morphology and size of the AgNPs, TEM and SEM analyses were performed. Figure 6 shows the size range of the nanoparticle between 17.63 and 24.60 nm, and the average particle size of 19.57 nm within the selected area of the SEM image confirmed the sizes in the nano range. The TEM image in Fig. 7 also confirms that the AgNPs are generally spherical and their average size was 7.7 nm. Also, all particles were single and pure. The small size of the AgNPs was useful as they could easily make a way to the cell membrane, which is an important characteristic of medical and pharmacologic purposes.

SEM image of AgNPs. SEM, scanning electron microscopy.

TEM image of AgNPs. TEM, transmission electron microscopy.
The exact size of the synthesized B. bifidum nanoparticles was obtained based on the TEM images. The results of the particle size distribution histogram showed that the nanoparticles' mean size was about 7.7 nm (Fig. 8).

Particle size distribution of synthetized AgNP.
FTIR analysis
FTIR analysis was carried out to characterize various functional groups in B. bifidum mixture responsible for reducing and stabilizing the synthesized AgNPs. As shown in Fig. 9, the FTIR spectrum represented several strong peaks at 3,435, 1,636, and 636 cm−1. The existence of peaks at 3,435 and 636 might be due to the OH stretching of alcohols and phenols or bending the stretching of hydrogen-bonded alcohols and phenols in the mixture with a typical strong peak centered at about 1,636 corresponding to stretching frequencies of the imidazolium ring. The absorption peak at 1,385 cm−1 indicated CH3 and C-N stretching vibration of aromatic amine.

FTIR spectra of AgNPs. FTIR, Fourier transform-infrared spectroscopy.
Disk diffusion test
According to the findings of the antibiogram assay for 100 strains of K. pneumoniae in Table 1, the highest resistance was observed for ciprofloxacin (70%) and tetracycline (60%), whereas the lowest was related to imipenem (60% susceptible) and cefoxitin (60% susceptible). Out of 70 isolates resistant to ciprofloxacin, 25 K. pneumoniae strains (25%) had efflux pump confirmed by PCR method and efflux pump inhibitor (CCCP) (Supplementary Table S2).
Antibiotic Resistance Profiles of Klebsiella pneumoniae Strains
Susceptibility determination
Significant antimicrobial effects, expressed as MIC of biosynthesized and commercial AgNPs against ciprofloxacin-resistant K. pneumonia, are given in Table 2. All isolates were exposed to the AgNPs at the concentrations of 1.95–500 μg/mL within 24 hr. The MIC results showed that the MIC values were within the range of 7.8–125 μg/mL against ciprofloxacin-resistant strains. Also, the isolates No. 7, 26, 46, and 53, and also the control positive strain (K. pneumoniae ATCC700603) showed the greatest toxicity at 125 μg/mL (MIC value).
The Minimum Inhibitory Concentration of Biosynthesized and Commercial Silver Nanoparticles on Ciprofloxacin-Resistant Strains of Klebsiella pneumoniae
MIC, minimum inhibitory concentration.
Real-time PCR of oqxAB genes
The relative expression of the oqxAB genes in 25 ciprofloxacin-resistant isolates of K. pneumoniae was assessed in the presence of both biosynthesized and commercial AgNPs by quantitative real-time PCR analysis. Figure 10 indicates that the oxqA expression significantly reduced in all K. pneumoniae isolates while exposing to biosynthesized and commercial AgNPs, whereas the greatest reduction occurred in isolates 20 and 21 (Supplementary Table S3). The results also revealed that the MIC of the synthetic AgNP remarkably reduced the expression level of oxqA gene in K. pneumoniae strains in comparison with the MIC of commercial AgNP (p < 0.001). Figure 11 shows a significant decrease in the expression of oqxB gene in the bacteria exposed to commercially and microbially synthesized AgNPs in comparison with gmk gene as the internal control, depicting the most considerable decrease in isolate 21 (Supplementary Table S4). In addition, both nanoparticles showed a relative decrease of more than 50% in all isolates, but the biosynthesized AgNPs were more toxic than the commercial ones (p < 0.001).

The expression level of oxqA decrease more than twofold in ciprofloxacin-resistant strains in the presence of both commercial and biosynthesized AgNPs (n = 3, **p < 0.01, ***p < 0.001).

The expression level of oxqB decrease in ciprofloxacin-resistant strains in the presence of commercial and biosynthesized AgNPs (n = 3, *p < 0.05, **p < 0.01, ***p < 0.001).
Discussion
Biological agents, including bacteria and fungi, are impressive candidates for nanoparticle synthesis due to their extreme diversity to be synthesized in different sizes and shapes.17,19 The actual and most reliable mechanism involved in the biosynthesis of AgNPs using microbes is not fully understood; however, it seems that the various enzymes secreted by microbes, like nitrate reductase aid in the bioreduction of metal ions to metal nanoparticles. 20
However, it is hypothesized that the nitrate reductase secreted by the microorganism is responsible for the reduction of Ag1+ ions and the bioreduction of silver ions to AgNPs. 21
AgNPs have attracted attention due to their broad application to novel technologies in chemistry, electronics, medicine, and biotechnology.22,23 In the present study, the B. bifidum strain was used for AgNPs synthesis, which was confirmed by changing its color from yellow to dark brown. This was further confirmed by UV-Vis, FTIR, TEM, SEM, EDX, XRD, and zeta analysis. Various reports indicate that bacteria are an appropriate option for the synthesis of AgNPs.24,25 Biological studies have indicated that Gram-positive Lactobacillus spp. can environmentally biosynthesize AgNPs, which act as reducing and capping agents.7,26 AgNPs can be synthesized using B. bifidum NCDC 229 by an economical and environmentally friendly process. 27
K. pneumoniae is currently considered as an immediate threat to human health due to the development of MDR strains associated with hospital-acquired infections and hypervirulent strains associated with severe community-acquired pneumonia. 28 K. pneumoniae uses different mechanisms to develop resistance against antibiotics, which may be due to its effect on the efflux system. 13 The multidrug efflux pump OqxAB is one of the most important efflux systems in K. pneumoniae. 29 Our study demonstrated the antibacterial effects of both commercial and biosynthesized AgNPs using B. bifidum against the OqxAB efflux pump on ciprofloxacin-resistant K. pneumoniae strains. The functions of the OqxAB efflux pump were investigated by the MIC method in association with the CCCP efflux pump inhibitor. As indicated in the results, MIC decreased in the presence of both commercial and synthesized AgNPs, which is consistent with other findings and reveals that efflux pumps play a pivotal role in resistance against antibiotics. We indicated that both commercially and microbially synthesized AgNPs could inhibit the visible growth of ciprofloxacin-resistant K. pneumoniae strains at a concentration within the range of 7.8–125 μg/mL.
Biofilm can be formed by attachment to most pathogenic bacteria on the surface. This structure can change bacterial properties, of which an increase in bacterial resistance is of great importance. The existence of an extracellular matrix around bacteria that prevents the penetration of drugs into the deeper layer of biofilm is the main mechanism to increase bacterial resistance. Almost all bacterial infections in humans are caused by mesophilic bacteria with an optimum temperature of around 37°C for growth; temperatures over 45°C can inhibit bacterial growth. Therefore, increasing the temperature can be used to kill such pathogenic bacteria. Increasing the temperature can be harmful to all body organs; however, local temperature increase to the body is not easily possible. Convenient tools developed by nanotechnology are helpful to solve this problem and are effective for the treatment of several diseases, like cancer and pathogenic infections. Metal nanoparticles (gold, silver, and copper) can excite free electrons on the surface of nanoparticles under external light at convenient wavelengths. This collective coherent oscillation of the conduction band electrons arises at the metal particles is called localized surface plasmon (LSP). Plasmonic metal nanoparticles (gold, silver, and cooper) absorb the light forming localized electric fields around the surface of nanoparticles, which is known as LSP resonance (LSPR). In this condition, some of the absorbed lights decay radiatively and nonradiatively by emitting photons and converting into phonons, respectively. By irradiating metal nanoparticles at the convenient wavelengths, free electrons can excite on the surface of nanoparticle leading to LSPR at the same frequency. This LSP can produce hot electrons. The energy of LSP can transfer to the electrons in the metal that can be relaxed as a thermal dissipation releasing thermal energy to the surroundings (photothermal effect). Through the light absorption process, the temperature inside the nanoparticles can be raised and transfer to the medium around particles, which can be used for photothermal therapy.
AgNPs can be used as antibacterial agents due to photothermal light-to-heat conversion and also they can exhibit strong plasmon resonance and LSP, which inhibit bacterial growth by increasing the temperature of the surrounding medium, where bacteria live.17,30–33
The exact mechanism behind the antimicrobial effect of AgNPs is still exploratory and has not been well understood. The antibacterial properties of AgNPs are the set of different mechanisms of action. Some researchers reported that silver ions reacted with proteins by binding with the sulfhydryl group and played a crucial role in the inactivation of pathogenic microbial species. It is also reported that they disengage respiratory electron transport from oxidative phosphorylation, which prevents enzymes involved in the respiratory chain or affects membrane permeability to protons and phosphate.34,35 Within the cell, silver ions can cause the deactivation of the cellular enzymes, DNA, and proteins by reacting with electron-donating groups and producing reactive oxygen species. Thus, it is rational to conclude that there is a higher chance of success with newer antimicrobial agents using microbial biogenic nanoparticles. 36
Drug efflux is a major mechanism of resistance in Gram-negative bacteria. In these systems, solutes are pumped out of the cell through an energy-dependent mechanism. Bacteria utilize efflux pumps to regulate their internal environment by removing toxic substances, including antimicrobial agents, metabolites, and quorum-sensing signal molecules. 37 Efflux pumps may be formed by a single component or multiple components, with the latter being found exclusively in Gram-negative bacteria. 38 Real-time PCR was applied to check the expression of the oqxAB genes after treatment by subinhibitory concentrations of our biosynthesized AgNPs. Our findings revealed that the synthesized AgNPs using probiotic bacteria significantly reduced the expression of the oqxAB genes in all K. pneumoniae strains. On the other hand, a comparison of antibacterial effects of commercial and biosynthesized AgNPs using B. bifidum on ciprofloxacin-resistant K. pneumoniae strains revealed that the antiefflux pump effects of microbially synthesized AgNPs was higher than commercial AgNPs and they were appropriate candidates for the treatment of an infection caused by K. pneumoniae strains. Several studies have indicated that AgNPs prevent the activity of the P-glycoprotein efflux pump in MDR cancer cells. 39 AgNPs have the potential to inhibit the multidrug-resistant strains of Staphylococcus aureus and Pseudomonas aeruginosa through the prevention of efflux pump activity. 40
Szabo et al. showed that ciprofloxacin concentration-dependent upregulation of the OqxAB efflux pump in K. pneumoniae is clonally related and contributes to the selection for higher fluoroquinolone resistance. 41
According to the TEM and zeta potential results, the synthesized AgNps were monodisperse and showed no agglomeration. The small size of nanoparticle (7.7 nm) can also suggest the appropriate effect of synthesized nanoparticles on the expression of the efflux pump gene in K. pneumoniae.
Our results generally supported that the biosynthesized AgNPs had higher antibacterial activity and could reduce the expression of oqxAB gene compared with commercial AgNPs. Strains 7, 26, 46, 53, and ATCC700603 showed the maximum resistance, and the most reduction of fold change belonged to the strain 21 in the expression of oqxA and oqxB genes.
Several studies have proposed that the toxicity of AgNps can be explained by surface oxidation and release of Ag+ on the medium. The Ag+ ion leaching results in the oxidation of metallic AgNp by dissolved oxygen and proton. In the synthesis process of biosynthesized nanoparticles, synthesized AgNp is dried in air and also exposure to oxygen can occur because of this oxidation of AgNp. There is an oxide layer on the AgNp surface, which can release Ag(I) or Ag(0) species that increase the antibacterial activity.
Another mechanism of the antibacterial effect of AgNps is that Ag+ can cause an increase in cellular oxidation stress, which induces reactive oxygen species production by interacting with respiratory chain proteins on the membrane and interrupts intracellular O2 reduction.
This biosynthesized AgNp mostly induces oxidation reaction in comparison with commercial AgNps. For the synthesis of commercial AgNps, chemical solutions, such as N,N-dimethylformamide, N2H4, NaBH4, and sodium citrate are used. However, in the biological method, a mixture of metabolites, organic compounds, and enzymes, like nitrate reductase is applied. Regarding commercially synthesized AgNps, stabilizers are added to prevent agglomeration of the nanoparticle, which can have a toxic effect, whereas in the biosynthesized methods, no stabilizers are needed. Chemical methods are very energy consuming associated with the risks of environmental pollution. Biological synthesis methods are eco-friendly and safe enough and consume no energy.
Conclusion
In conclusion, using bacteria to synthesize AgNps is an eco-friendly, cost-effective, and exciting approach. The synthesis of AgNps using B. bifidum with spherical shape was confirmed by characterization assays. Biosynthesized AgNPs have the potential to inhibit the growth of ciprofloxacin-resistant K. pneumoniae strains and reduce the expression of OxqAB efflux pump genes in these isolates. Evaluation of the toxicity of these biosynthesized AgNPs in animal models should be considered in future studies. The current study suggested that efflux pumps can be an attractive target for our biosynthesized AgNPs.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.